
Promotion of Protein Solubility and Reduction in Stiffness in Human Lenses by Aggrelyte-1: Implications for Reversing Presbyopia

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Stability of Aggrelytes under Physiological Conditions
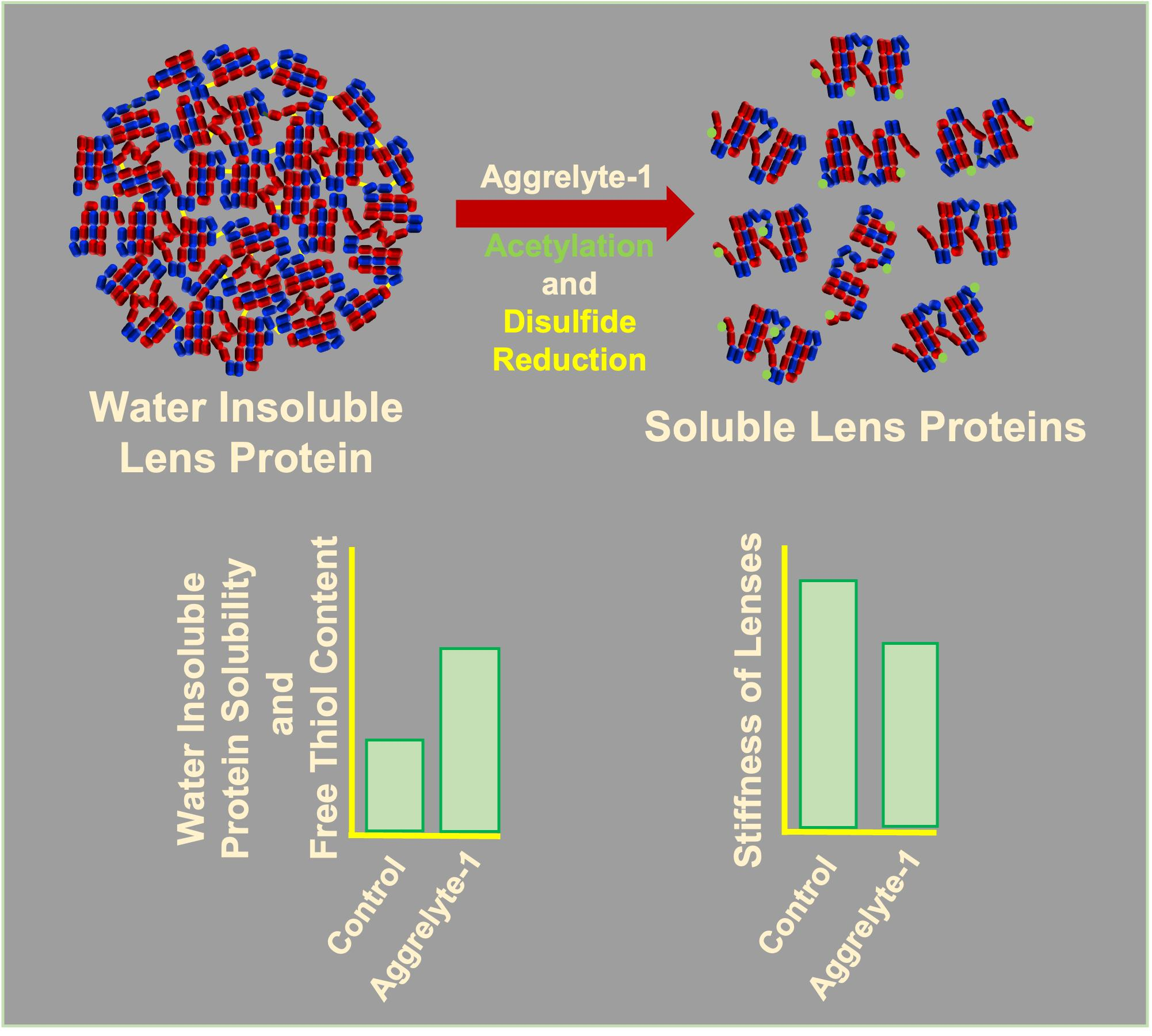
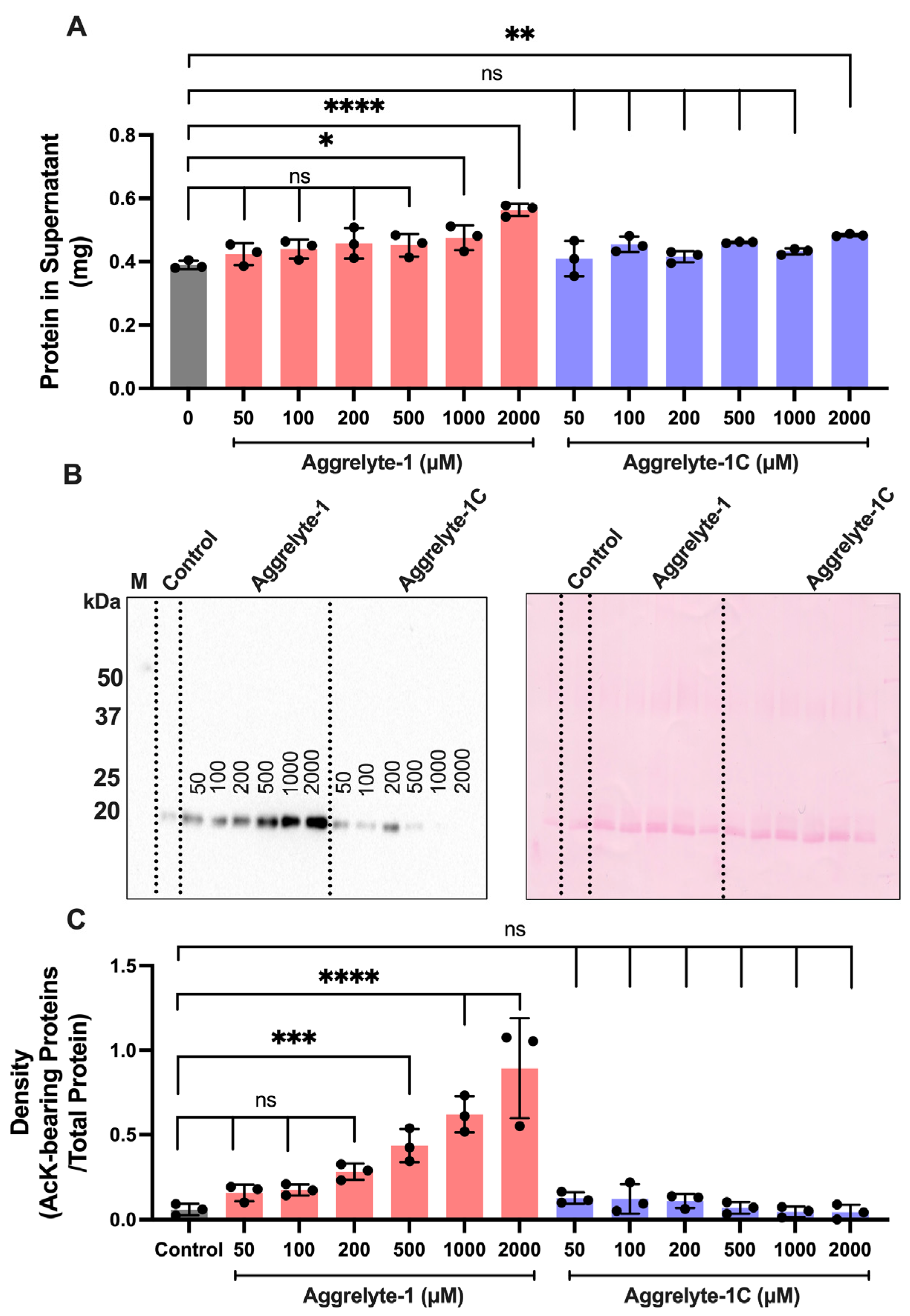
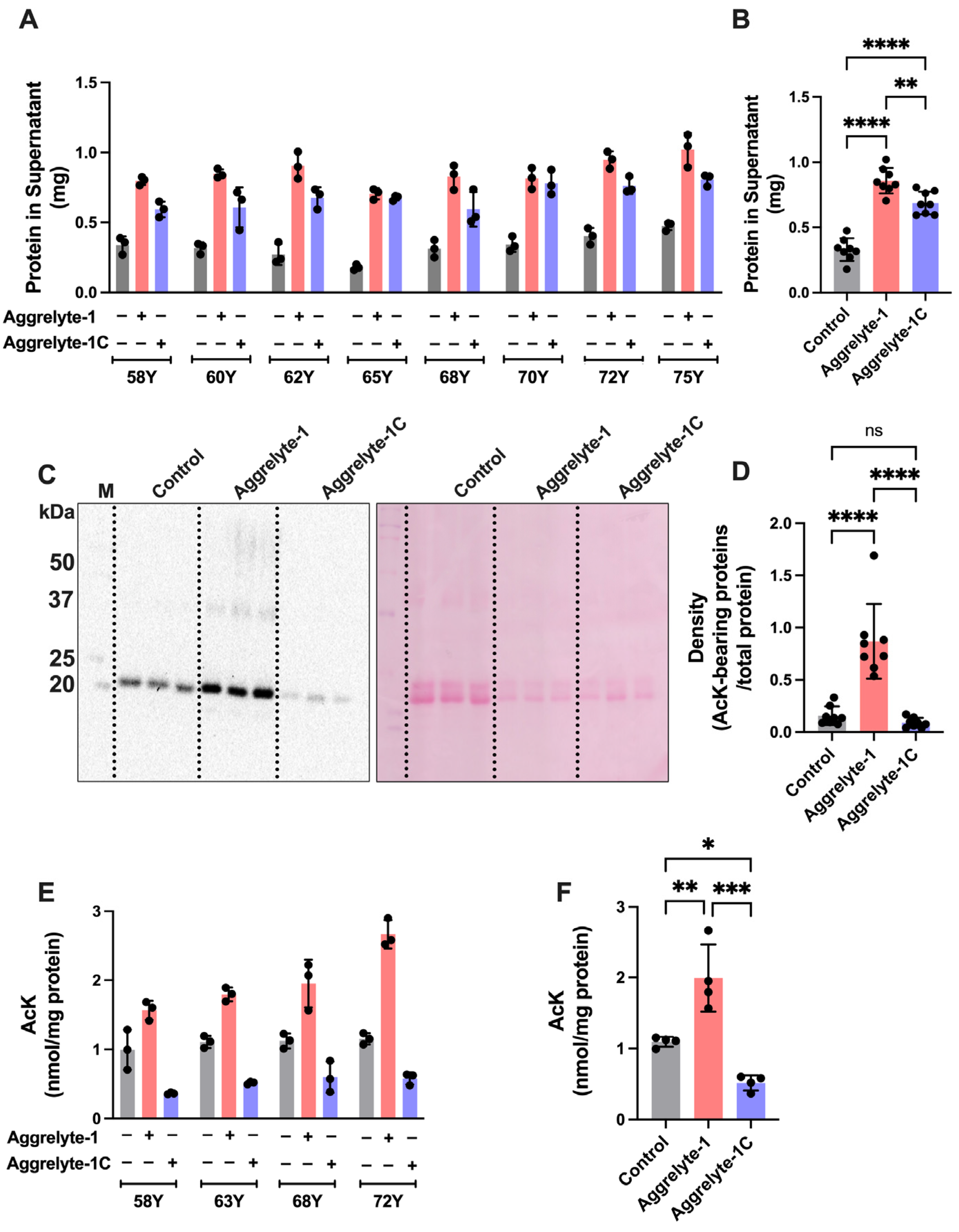
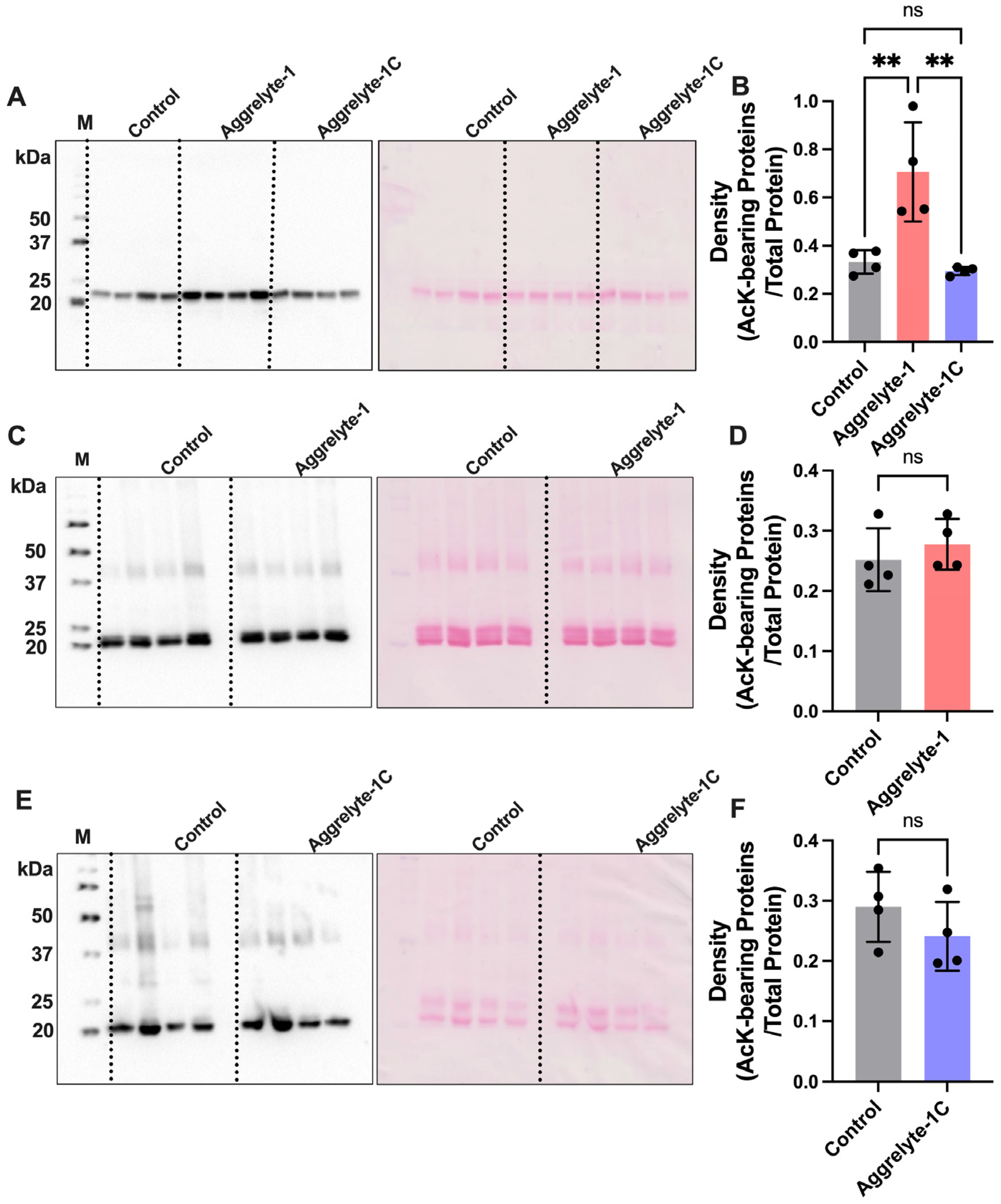
2.2. Aggrelyte-1 Solubilizes Water-Insoluble Lens Proteins (WI) and Increases the AcK Modification in Solubilized Proteins of Aged Human Lenses
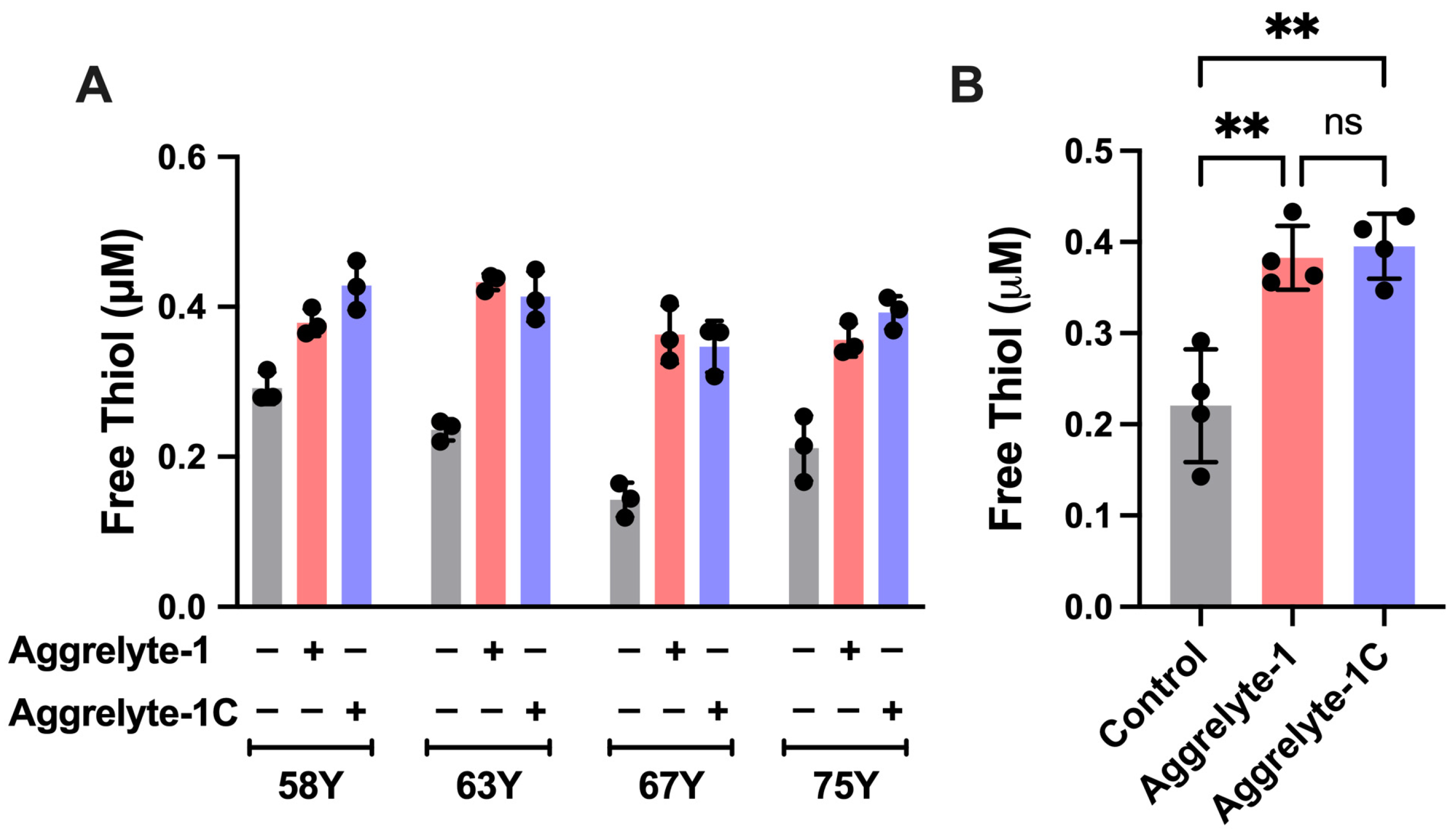
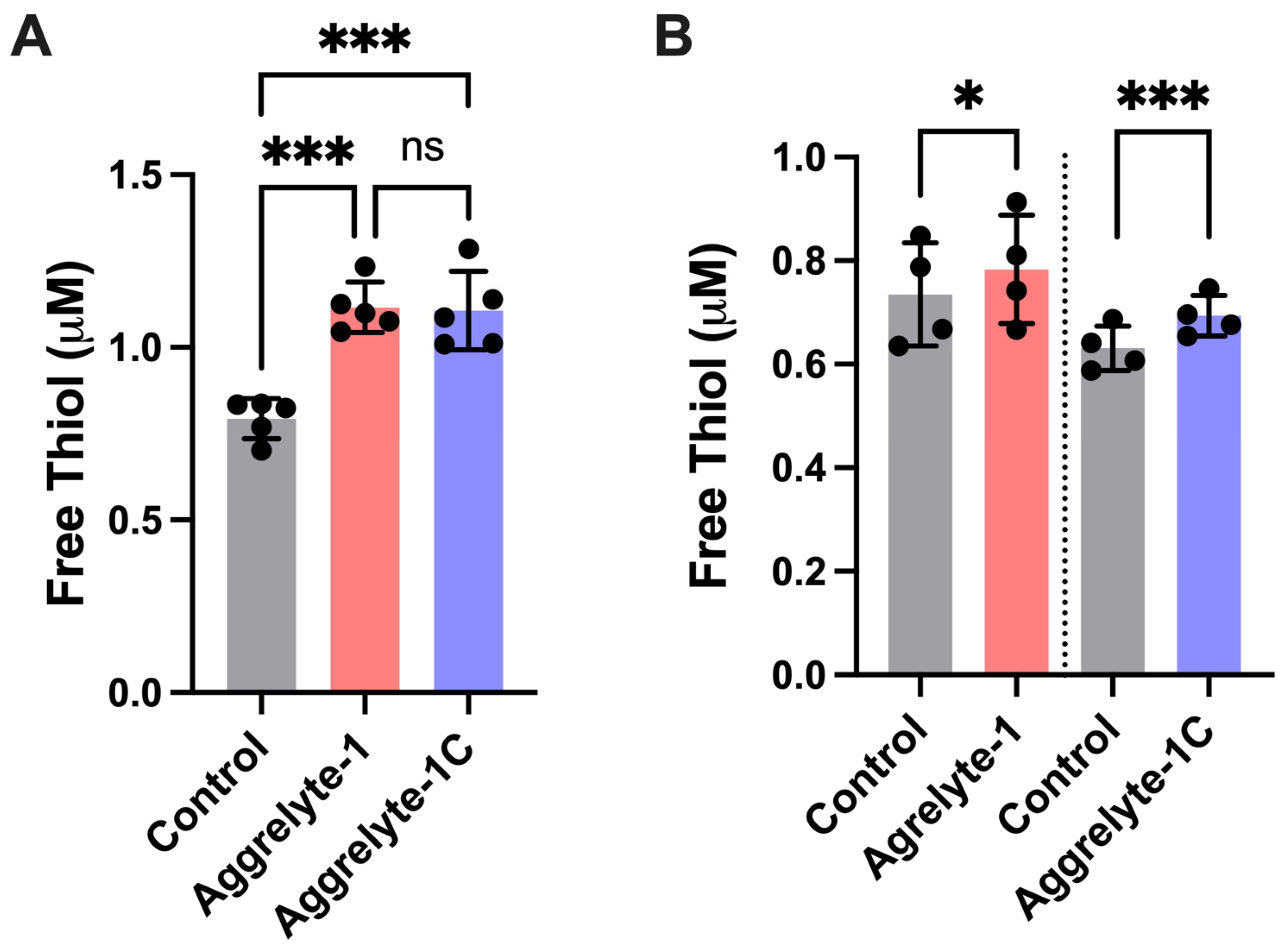
2.3. Aggrelytes Increase the Protein–Thiol Content
2.4. Aggrelyte-1 Mildly Alters the Structure of α-Crystallin
2.5. Aggrelytes Are Not Cytotoxic
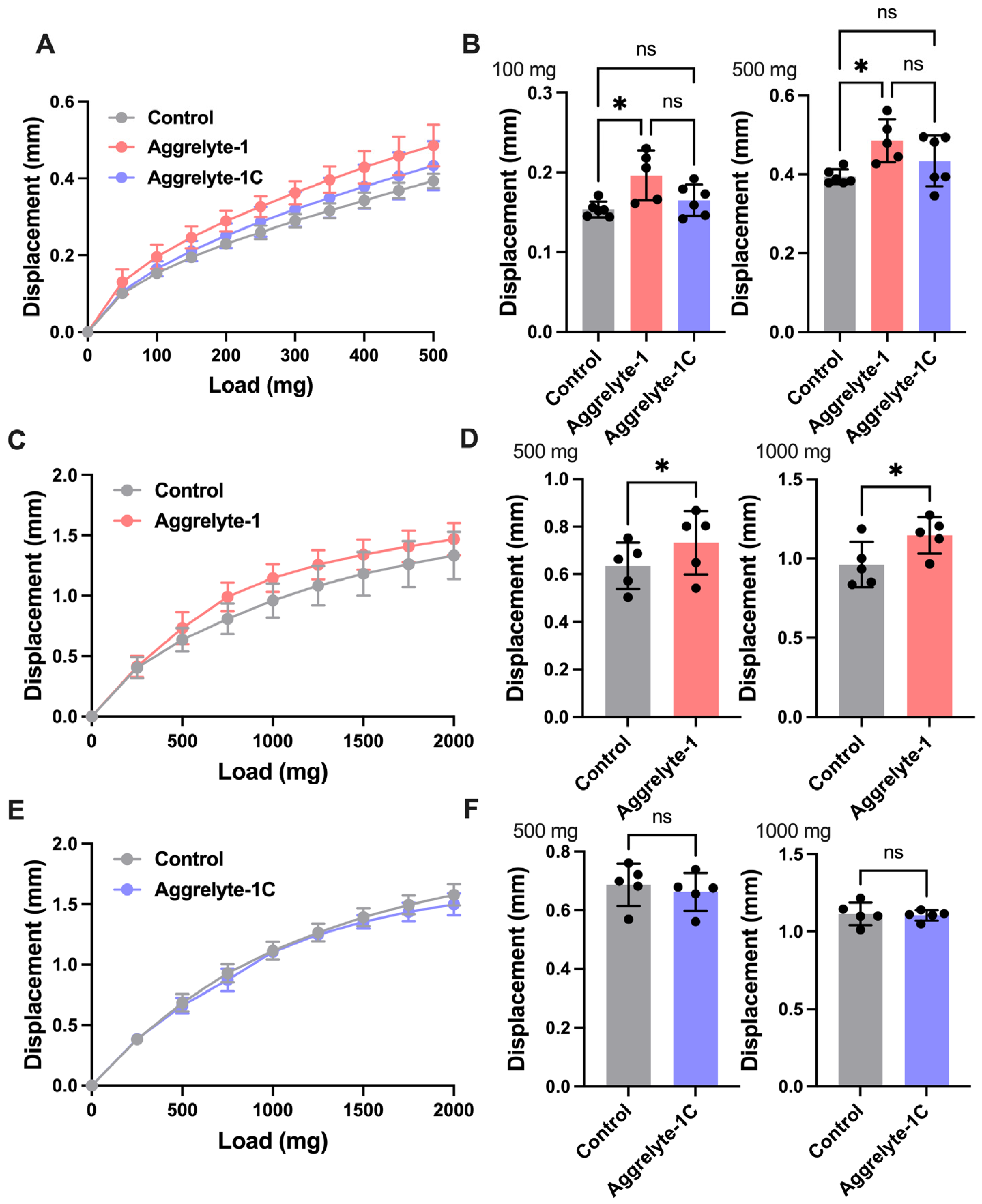
2.6. Aggrelytes Reduce Lens Stiffness
3. Discussion
4. Materials and Methods
4.1. Chemicals
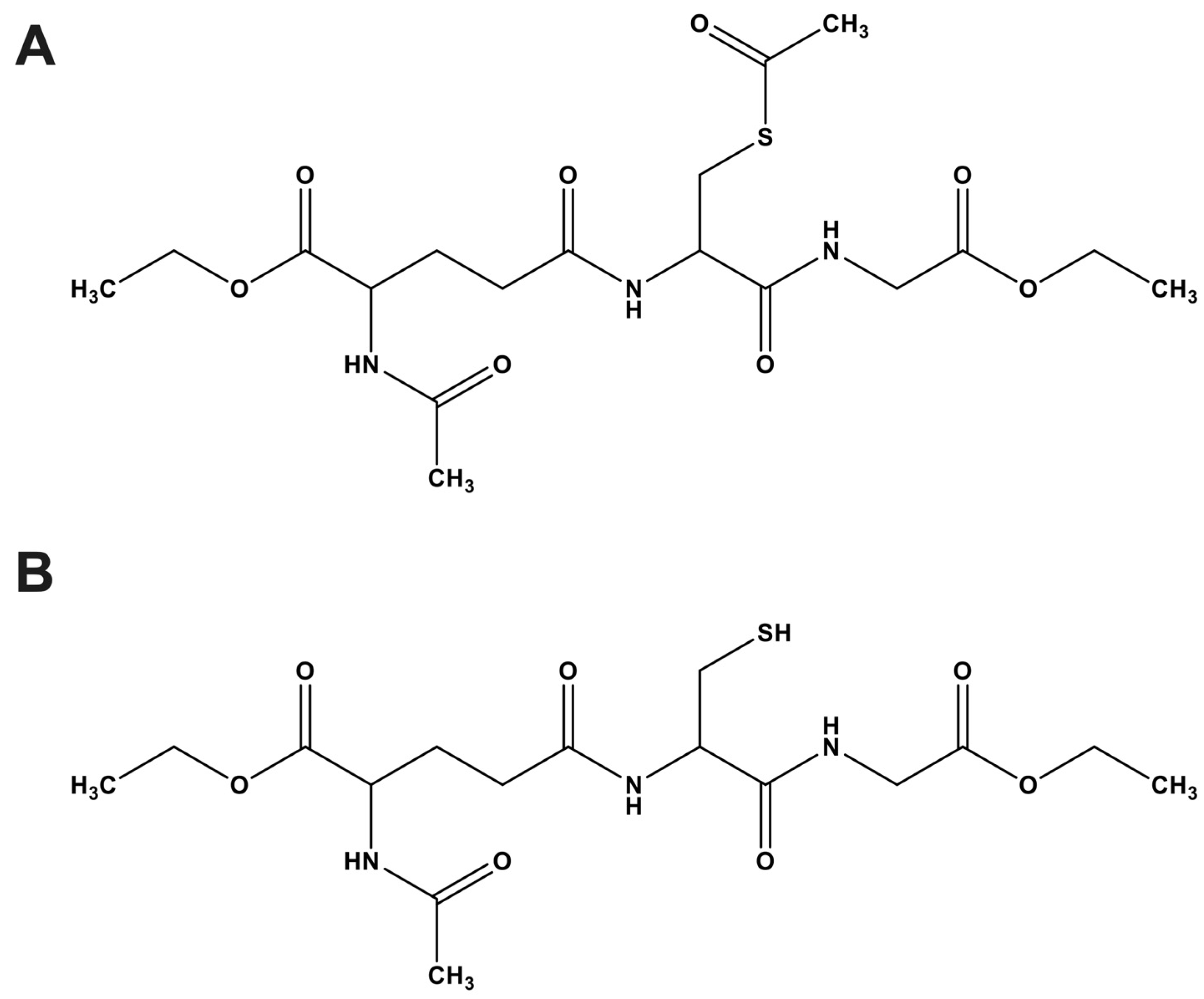
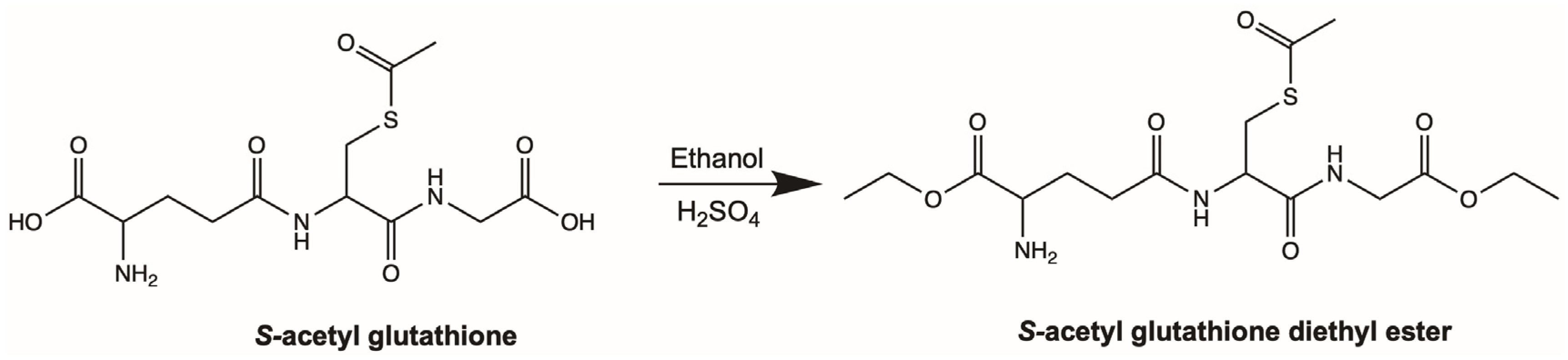
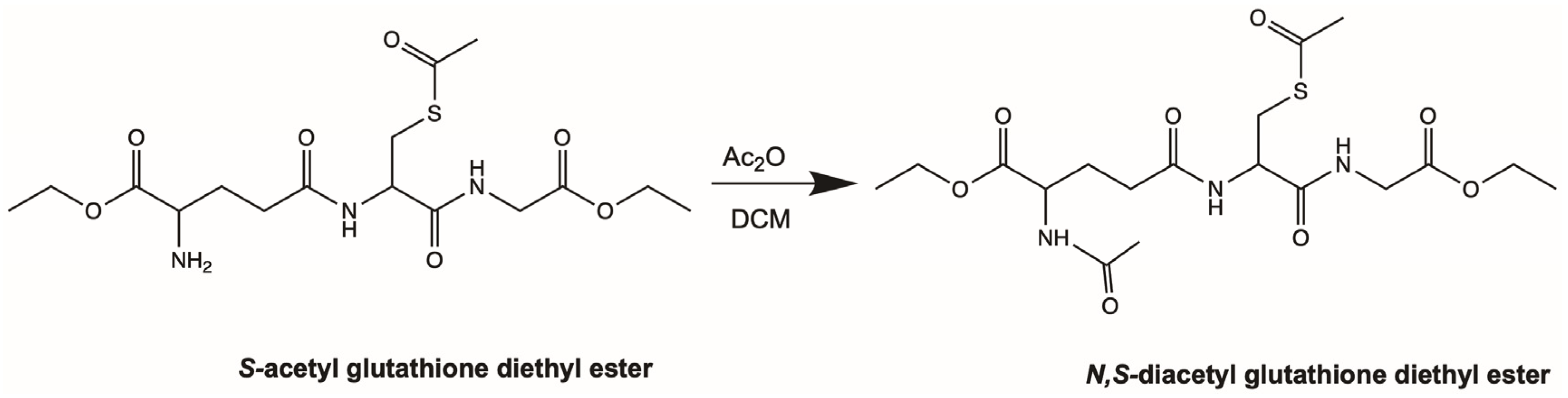
4.2. Synthesis of Aggrelyte-1
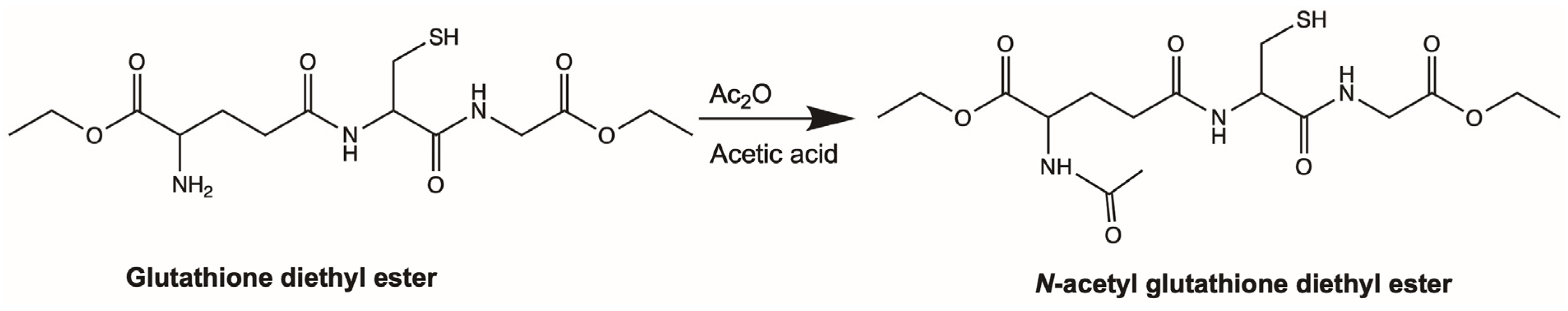
4.3. Synthesis of Aggrelyte-1C
4.4. Isolation of Water-Insoluble (WI) Protein from Aged Human Lenses
4.5. Protein Solubility by Aggrelytes
4.6. Western Blotting for AcK and Crystallin Subunits
4.7. LC–MS/MS Measurement of AcK
4.8. Free Protein–Thiol Estimation
4.9. Fluorescence and CD Experiments
4.10. Chaperone Activity Assay
4.11. Toxicity Studies
4.12. Lens Stiffness Measurements
4.13. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Fricke, T.R.; Tahhan, N.; Resnikoff, S.; Papas, E.; Burnett, A.; Ho, S.M.; Naduvilath, T.; Naidoo, K.S. Global Prevalence of Presbyopia and Vision Impairment from Uncorrected Presbyopia: Systematic Review, Meta-analysis, and Modelling. Ophthalmology 2018, 125, 1492–1499. [Google Scholar]
- Holden, B.A.; Fricke, T.R.; Ho, S.M.; Wong, R.; Schlenther, G.; Cronje, S.; Burnett, A.; Papas, E.; Naidoo, K.S.; Frick, K.D. Global vision impairment due to uncorrected presbyopia. Arch Ophthalmol. 2008, 126, 1731–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frick, K.D.; Joy, S.M.; Wilson, D.A.; Naidoo, K.S.; Holden, B.A. The Global Burden of Potential Productivity Loss from Uncorrected Presbyopia. Ophthalmology 2015, 122, 1706–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandi, S.K.; Rankenberg, J.; Rakete, S.; Nahomi, R.B.; Glomb, M.A.; Linetsky, M.D.; Nagaraj, R.H. Glycation-mediated protein crosslinking and stiffening in mouse lenses are inhibited by carboxitin in vitro. Glycoconj. J. 2021, 38, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Boscia, F.; Grattagliano, I.; Vendemiale, G.; Micelli-Ferrari, T.; Altomare, E. Protein oxidation and lens opacity in humans. Investig. Ophthalmol. Vis. Sci. 2000, 41, 2461–2465. [Google Scholar] [PubMed]
- Wilmarth, P.A.; Tanner, S.; Dasari, S.; Nagalla, S.R.; Riviere, M.A.; Bafna, V.; Pevzner, P.A.; David, L.L. Age-related changes in human crystallins determined from comparative analysis of post-translational modifications in young and aged lens: Does deamidation contribute to crystallin insolubility? J. Proteome Res. 2006, 5, 2554–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaraj, R.H.; Linetsky, M.; Stitt, A.W. The pathogenic role of Maillard reaction in the aging eye. Amino Acids 2012, 42, 1205–1220. [Google Scholar] [CrossRef]
- Korlimbinis, A.; Berry, Y.; Thibault, D.; Schey, K.L.; Truscott, R.J. Protein aging: Truncation of aquaporin 0 in human lens regions is a continuous age-dependent process. Exp. Eye Res. 2009, 88, 966–973. [Google Scholar] [CrossRef] [Green Version]
- Hooi, M.Y.; Raftery, M.J.; Truscott, R.J. Racemization of two proteins over our lifespan: Deamidation of asparagine 76 in gammaS crystallin is greater in cataract than in normal lenses across the age range. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3554–3561. [Google Scholar] [CrossRef]
- Sharma, K.K.; Santhoshkumar, P. Lens aging: Effects of crystallins. Biochim. Biophys. Acta 2009, 1790, 1095–1108. [Google Scholar] [CrossRef] [Green Version]
- Berthoud, V.M.; Beyer, E.C. Oxidative stress, lens gap junctions, and cataracts. Antioxid. Redox. Signal 2009, 11, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.C.; Peng, Y.J.; Chen, S.J.; Chiou, S.H. Senile cataracts and oxidative stress. J. Clin. Gerontol. Geri. 2010, 1, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Garner, M.H.; Spector, A. Selective oxidation of cysteine and methionine in normal and senile cataractous lenses. Proc. Natl. Acad. Sci. USA 1980, 77, 1274–1277. [Google Scholar] [CrossRef] [Green Version]
- Garner, W.H.; Garner, M.H. Protein Disulfide Levels and Lens Elasticity Modulation: Applications for Presbyopia. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2851–2863. [Google Scholar] [CrossRef] [Green Version]
- Korenfeld, M.S.; Robertson, S.M.; Stein, J.M.; Evans, D.G.; Rauchman, S.H.; Sall, K.N.; Venkataraman, S.; Chen, B.L.; Wuttke, M.; Burns, W. Topical lipoic acid choline ester eye drop for improvement of near visual acuity in subjects with presbyopia: A safety and preliminary efficacy trial. Eye 2021, 35, 3292–3301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Q.; Singh, M.; Nair, A.; Larin, K.V.; Aglyamov, S.R. Elasticity Changes in the Crystalline Lens during Oxidative Damage and the Antioxidant Effect of Alpha-Lipoic Acid Measured by Optical Coherence Elastography. Photonics 2021, 8, 207. [Google Scholar] [CrossRef]
- Drazic, A.; Myklebust, L.M.; Ree, R.; Arnesen, T. The world of protein acetylation. Biochim. Biophys. Acta 2016, 1864, 1372–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, I.; Conrad, R.J.; Verdin, E.; Ott, M. Lysine Acetylation Goes Global: From Epigenetics to Metabolism and Therapeutics. Chem. Rev. 2018, 118, 1216–1252. [Google Scholar] [CrossRef] [Green Version]
- James, A.M.; Smith, C.L.; Smith, A.C.; Robinson, A.J.; Hoogewijs, K.; Murphy, M.P. The Causes and Consequences of Nonenzymatic Protein Acylation. Trends Biochem. Sci. 2018, 43, 921–932. [Google Scholar] [CrossRef] [Green Version]
- Nagaraj, R.H.; Nahomi, R.B.; Shanthakumar, S.; Linetsky, M.; Padmanabha, S.; Pasupuleti, N.; Wang, B.; Santhoshkumar, P.; Panda, A.K.; Biswas, A. Acetylation of alphaA-crystallin in the human lens: Effects on structure and chaperone function. Biochim. Biophys. Acta 2012, 1822, 120–129. [Google Scholar] [CrossRef] [Green Version]
- Nandi, S.K.; Rakete, S.; Nahomi, R.B.; Michel, C.; Dunbar, A.; Fritz, K.S.; Nagaraj, R.H. Succinylation Is a Gain-of-Function Modification in Human Lens alphaB-Crystallin. Biochemistry 2019, 58, 1260–1274. [Google Scholar] [CrossRef]
- Nahomi, R.B.; Nandi, S.K.; Rakete, S.; Michel, C.; Fritz, K.S.; Nagaraj, R.H. Lysine malonylation and propionylation are prevalent in human lens proteins. Exp. Eye Res. 2020, 190, 107864. [Google Scholar] [CrossRef]
- Nandi, S.K.; Nahomi, R.B.; Harris, P.S.; Michel, C.R.; Fritz, K.S.; Nagaraj, R.H. The absence of SIRT3 and SIRT5 promotes the acetylation of lens proteins and improves the chaperone activity of alpha-crystallin in mouse lenses. Exp. Eye Res. 2019, 182, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nahomi, R.B.; Huang, R.; Nandi, S.K.; Wang, B.; Padmanabha, S.; Santhoshkumar, P.; Filipek, S.; Biswas, A.; Nagaraj, R.H. Acetylation of lysine 92 improves the chaperone and anti-apoptotic activities of human alphaB-crystallin. Biochemistry 2013, 52, 8126–8138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, A.K.; Teranishi, K.; Ambroso, M.R.; Isas, J.M.; Vazquez-Sarandeses, E.; Lee, J.Y.; Melo, A.A.; Pandey, P.; Merken, D.; Berndt, L.; et al. Lysine acetylation regulates the interaction between proteins and membranes. Nat. Commun. 2021, 12, 6466. [Google Scholar] [CrossRef] [PubMed]
- Ree, R.; Varland, S.; Arnesen, T. Spotlight on protein N-terminal acetylation. Exp. Mol. Med. 2018, 50, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phadte, A.S.; Santhoshkumar, P.; Sharma, K.K. alphaA-crystallin-derived minichaperone stabilizes alphaAG98R-crystallin by affecting its zeta potential. Mol. Vis. 2018, 24, 297–304. [Google Scholar] [PubMed]
- Rasmussen, T.; van de Weert, M.; Jiskoot, W.; Kasimova, M.R. Thermal and acid denaturation of bovine lens alpha-crystallin. Proteins 2011, 79, 1747–1758. [Google Scholar] [CrossRef]
- Kamei, A. Glutathione levels of the human crystalline lens in aging and its antioxidant effect against the oxidation of lens proteins. Biol. Pharm. Bull. 1993, 16, 870–875. [Google Scholar] [CrossRef] [Green Version]
- Baradia, H.; Nikahd, N.; Glasser, A. Mouse lens stiffness measurements. Exp. Eye Res. 2010, 91, 300–307. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panja, S.; Gaikwad, H.; Rankenberg, J.; Nam, M.-H.; Nagaraj, R.H. Promotion of Protein Solubility and Reduction in Stiffness in Human Lenses by Aggrelyte-1: Implications for Reversing Presbyopia. Int. J. Mol. Sci. 2023, 24, 2196. https://doi.org/10.3390/ijms24032196
Panja S, Gaikwad H, Rankenberg J, Nam M-H, Nagaraj RH. Promotion of Protein Solubility and Reduction in Stiffness in Human Lenses by Aggrelyte-1: Implications for Reversing Presbyopia. International Journal of Molecular Sciences. 2023; 24(3):2196. https://doi.org/10.3390/ijms24032196
Chicago/Turabian StylePanja, Sudipta, Hanmant Gaikwad, Johanna Rankenberg, Mi-Hyun Nam, and Ram H. Nagaraj. 2023. "Promotion of Protein Solubility and Reduction in Stiffness in Human Lenses by Aggrelyte-1: Implications for Reversing Presbyopia" International Journal of Molecular Sciences 24, no. 3: 2196. https://doi.org/10.3390/ijms24032196