
Single-Molecule Localization Microscopy of Presynaptic Active Zones in Drosophila melanogaster after Rapid Cryofixation
 , ,
, , 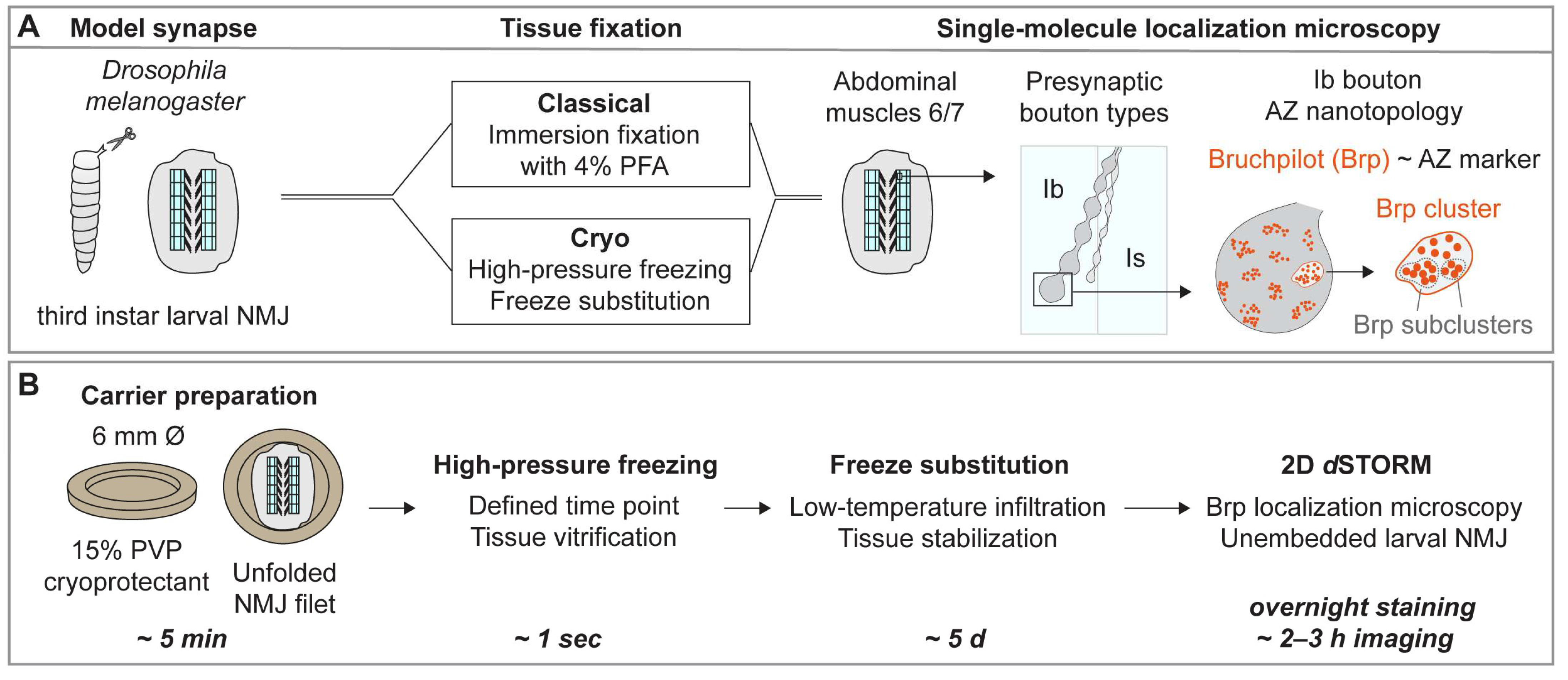{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- Which HPF/FS protocol is most suitable to preserve Brp clusters in AZs for quantitative (sub-)cluster analysis?
- How does the structure of Brp clusters differ in HPF/FS and conventionally fixed samples using standardized quantitative parameters?
2. Results
2.1. dSTORM Imaging on HPF/FS Samples
2.2. Preservation of Bouton Architecture after HPF/FS
2.3. AZ Nanotopology in HPF/FS Larval Muscle Filets
2.4. AZ Subcluster Analysis in HPF/FS Larval Muscle Filets
3. Discussion
3.1. Standardized HPF/FS-SMLM Workflow for AZ Nanotopology
3.2. Substantially Different Bouton Morphology in HPF/FS Samples
3.3. Smaller AZs in HPF/FS Samples
3.4. Brp SCs Are Preserved in Cryofixated Samples
3.5. Perspectives on Presynaptic Structure–Function Relationships
4. Material and Methods
4.1. Fly Stocks
4.2. High-Pressure Freezing
4.3. Freeze Substitution
4.4. Chemical Fixation
4.5. Staining and Immunofluorescence
4.6. dSTORM
4.7. Data Evaluation
4.8. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Emperador-Melero, J.; Kaeser, P.S. Assembly of the presynaptic active zone. Curr. Opin. Neurobiol. 2020, 63, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.; Sauer, M.; Geis, C. Super-resolving Microscopy in Neuroscience. Chem. Rev. 2021, 121, 11971–12015. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, G. Fine Structure Immunocytochemistry; Springer: Berlin, Germany, 1993; pp. 26–89. [Google Scholar]
- Tanaka, K.A.; Suzuki, K.G.; Shirai, Y.M.; Shibutani, S.T.; Miyahara, M.S.; Tsuboi, H.; Yahara, M.; Yoshimura, A.; Mayor, S.; Fujiwara, T.K.; et al. Membrane molecules mobile even after chemical fixation. Nat. Methods 2010, 7, 865–866. [Google Scholar] [CrossRef]
- Imig, C.; Min, S.W.; Krinner, S.; Arancillo, M.; Rosenmund, C.; Südhof, T.C.; Rhee, J.; Brose, N.; Cooper, B.H. The morphological and molecular nature of synaptic vesicle priming at presynaptic active zones. Neuron 2014, 84, 416–431. [Google Scholar] [CrossRef] [Green Version]
- Weimer, R.M.; Gracheva, E.O.; Meyrignac, O.; Miller, K.G.; Richmond, J.E.; Bessereau, J.L. UNC-13 and UNC-10/rim localize synaptic vesicles to specific membrane domains. J. Neurosci. 2006, 26, 8040–8047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moor, H.; Bellin, G.; Sandri, C.; Akert, K. The influence of high pressure freezing on mammalian nerve tissue. Cell Tissue Res. 1980, 209, 201–216. [Google Scholar] [CrossRef]
- Riehle, U. Über die Vitrifizierung verdünnter wässriger Lösungen. Doctoral Thesis, ETH Zürich, Zürich, Switzerland, 1968. [Google Scholar]
- McDonald, K. High-pressure freezing for preservation of high resolution fine structure and antigenicity for immunolabeling. Methods Mol. Biol. 1999, 117, 77–97. [Google Scholar]
- Ripper, D.; Schwarz, H.; Stierhof, Y.D. Cryo-section immunolabelling of difficult to preserve specimens: Advantages of cryofixation, freeze-substitution and rehydration. Biol. Cell 2008, 100, 109–123. [Google Scholar] [CrossRef]
- Steinbrecht, R.A.; Müller, M. Freeze-Substitution and Freeze-Drying. In Cryotechniques in Biological Electron Microscopy; Steinbrecht, R.A., Zierold, K., Eds.; Springer: Berlin Heidelberg, Germany, 1987; pp. 149–172. [Google Scholar]
- Korogod, N.; Petersen, C.C.; Knott, G.W. Ultrastructural analysis of adult mouse neocortex comparing aldehyde perfusion with cryo fixation. eLife 2015, 4, e05793. [Google Scholar] [CrossRef] [PubMed]
- Stigloher, C.; Zhan, H.; Zhen, M.; Richmond, J.; Bessereau, J.L. The presynaptic dense projection of the Caenorhabditis elegans cholinergic neuromuscular junction localizes synaptic vesicles at the active zone through SYD-2/liprin and UNC-10/RIM-dependent interactions. J. Neurosci. 2011, 31, 4388–4396. [Google Scholar] [CrossRef] [Green Version]
- Zhan, H.; Bruckner, J.; Zhang, Z.; O’Connor-Giles, K. Three-dimensional imaging of Drosophila motor synapses reveals ultrastructural organizational patterns. J. Neurogenet 2016, 30, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, R.; Michanski, S.; Wichmann, C. Vesicle sub-pool organization at inner hair cell ribbon synapses. EMBO Rep. 2018, 19, e44937. [Google Scholar] [CrossRef]
- Lichter, K.; Paul, M.M.; Pauli, M.; Schoch, S.; Kollmannsberger, P.; Stigloher, C.; Heckmann, M.; Sirén, A.L. Ultrastructural analysis of wild-type and RIM1alpha knockout active zones in a large cortical synapse. Cell Rep. 2022, 40, 111382. [Google Scholar] [CrossRef]
- Ehmann, N.; van de Linde, S.; Alon, A.; Ljaschenko, D.; Keung, X.Z.; Holm, T.; Rings, A.; DiAntonio, A.; Hallermann, S.; Ashery, U.; et al. Quantitative super-resolution imaging of Bruchpilot distinguishes active zone states. Nat. Commun. 2014, 5, 4650. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.M.; Pauli, M.; Ehmann, N.; Hallermann, S.; Sauer, M.; Kittel, R.J.; Heckmann, M. Bruchpilot and Synaptotagmin collaborate to drive rapid glutamate release and active zone differentiation. Front Cell Neurosci. 2015, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrestani, A.; Pauli, M.; Kollmannsberger, P.; Repp, F.; Kittel, R.J.; Eilers, J.; Doose, S.; Sauer, M.; Sirén, A.L.; Heckmann, M.; et al. Active zone compaction correlates with presynaptic homeostatic potentiation. Cell Rep. 2021, 37, 109770. [Google Scholar] [CrossRef]
- Paul, M.M.; Dannhäuser, S.; Morris, L.; Mrestani, A.; Hübsch, M.; Gehring, J.; Hatzopoulos, G.N.; Pauli, M.; Auger, G.M.; Bornschein, G.; et al. The human cognition-enhancing CORD7 mutation increases active zone number and synaptic release. Brain 2022, 145, 3787–3802. [Google Scholar] [CrossRef] [PubMed]
- Cavalier, A.; Spehner, D.; Humbel, B.M. Handbook of Cryo-Preparation Methods for Electron Microscopy; CRC Press: Boca Raton, FL, USA, 2009; pp. 321–341. [Google Scholar]
- Jiao, W.; Masich, S.; Franzen, O.; Shupliakov, O. Two pools of vesicles associated with the presynaptic cytosolic projection in Drosophila neuromuscular junctions. J. Struct. Biol. 2010, 172, 389–394. [Google Scholar] [CrossRef] [Green Version]
- Collins, J.S.; Goldsmith, T.H. Spectral properties of fluorescence induced by glutaraldehyde fixation. J. Histochem. Cytochem. 1981, 29, 411–414. [Google Scholar] [CrossRef]
- Pauli, M.; Paul, M.M.; Proppert, S.; Mrestani, A.; Sharifi, M.; Repp, F.; Kürzinger, L.; Kollmannsberger, P.; Sauer, M.; Heckmann, M.; et al. Targeted volumetric single-molecule localization microscopy of defined presynaptic structures in brain sections. Commun. Biol. 2021, 4, 407. [Google Scholar] [CrossRef]
- Tang, A.H.; Chen, H.; Li, T.P.; Metzbower, S.R.; MacGillavry, H.D.; Blanpied, T.A. A trans-synaptic nanocolumn aligns neurotransmitter release to receptors. Nature 2016, 536, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, H.; Ariyoshi, T.; Kimpara, N.; Sugao, K.; Taiko, I.; Takikawa, K.; Asanuma, D.; Namiki, S.; Hirose, K. Synaptic weight set by Munc13-1 supramolecular assemblies. Nat. Neurosci. 2018, 21, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Böhme, M.A.; McCarthy, A.W.; Grasskamp, A.T.; Beuschel, C.B.; Goel, P.; Jusyte, M.; Laber, D.; Huang, S.; Rey, U.; Petzoldt, A.G.; et al. Rapid active zone remodeling consolidates presynaptic potentiation. Nat. Commun. 2019, 10, 1085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dannhäuser, S.; Mrestani, A.; Gundelach, F.; Pauli, M.; Komma, F.; Kollmannsberger, P.; Sauer, M.; Heckmann, M.; Paul, M.M. Endogenous tagging of Unc-13 reveals nanoscale reorganization at active zones during presynaptic homeostatic potentiation. Front. Cell Neurosci. 2022, 16, 1074304. [Google Scholar] [CrossRef]
- Akbergenova, Y.; Cunningham, K.L.; Zhang, Y.V.; Weiss, S.; Littleton, J.T. Characterization of developmental and molecular factors underlying release heterogeneity at Drosophila synapses. eLife 2018, 7, e38268. [Google Scholar] [CrossRef]
- Gratz, S.J.; Goel, P.; Bruckner, J.J.; Hernandez, R.X.; Khateeb, K.; Macleod, G.T.; Dickman, D.; O’Connor-Giles, K.M. Endogenous Tagging Reveals Differential Regulation of Ca2+ Channels at Single Active Zones during Presynaptic Homeostatic Potentiation and Depression. J. Neurosci. 2019, 39, 2416–2429. [Google Scholar]
- Newman, Z.L.; Bakshinskaya, D.; Schultz, R.; Kenny, S.J.; Moon, S.; Aghi, K.; Stanley, C.; Marnani, N.; Li, R.; Bleier, J.; et al. Determinants of synapse diversity revealed by super-resolution quantal transmission and active zone imaging. Nat. Commun. 2022, 13, 229. [Google Scholar] [CrossRef]
- Moor, H. Theory and Practice of High Pressure Freezing. In Cryotechniques in Biological Electron Microscopy; Steinbrecht, R.A., Zierold, K., Eds.; Springer: Berlin Heidelberg, Germany, 1987; pp. 175–191. [Google Scholar]
- Watanabe, S.; Rost, B.R.; Camacho-Perez, M.; Davis, M.W.; Sohl-Kielczynski, B.; Rosenmund, C.; Jorgensen, E.M. Ultrafast endocytosis at mouse hippocampal synapses. Nature 2013, 504, 242–247. [Google Scholar] [CrossRef] [Green Version]
- Kusick, G.F.; Chin, M.; Raychaudhuri, S.; Lippmann, K.; Adula, K.P.; Hujber, E.J.; Vu, T.; Davis, M.W.; Jorgensen, E.M.; Watanabe, S. Synaptic vesicles transiently dock to refill release sites. Nat. Neurosci. 2020, 23, 1329–1338. [Google Scholar] [CrossRef]
- Dawydow, A.; Gueta, R.; Ljaschenko, D.; Ullrich, S.; Hermann, M.; Ehmann, N.; Gao, S.; Fiala, A.; Langenhan, T.; Nagel, G.; et al. Channelrhodopsin-2-XXL, a powerful optogenetic tool for low-light applications. Proc. Natl. Acad. Sci. USA 2014, 111, 13972–13977. [Google Scholar] [CrossRef] [Green Version]
- Schieber, N.L.; Machado, P.; Markert, S.M.; Stigloher, C.; Schwab, Y.; Steyer, A.M. Minimal resin embedding of multicellular specimens for targeted FIB-SEM imaging. Methods Cell Biol. 2017, 140, 69–83. [Google Scholar] [PubMed]
- Tokuyasu, K.T. A technique for ultracryotomy of cell suspensions and tissues. J. Cell Biol. 1973, 57, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Stewart, B.A.; Atwood, H.L.; Renger, J.J.; Wang, J.; Wu, C.F. Improved stability of Drosophila larval neuromuscular preparations in haemolymph-like physiological solutions. J. Comp. Physiol. A 1994, 175, 179–191. [Google Scholar] [CrossRef] [PubMed]
- van de Linde, S.; Kasper, R.; Heilemann, M.; Sauer, M. Photoswitching microscopy with standard fluorophores. Appl. Phys. B 2008, 93, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Tokunaga, M.; Imamoto, N.; Sakata-Sogawa, K. Highly inclined thin illumination enables clear single-molecule imaging in cells. Nat. Methods 2008, 5, 159–161. [Google Scholar] [CrossRef]
- Heilemann, M.; van de Linde, S.; Schüttpelz, M.; Kasper, R.; Seefeldt, B.; Mukherjee, A.; Tinnefeld, P.; Sauer, M. Subdiffraction-resolution fluorescence imaging with conventional fluorescent probes. Angew Chem. 2008, 47, 6172–6176. [Google Scholar] [CrossRef]
- Wolter, S.; Schüttpelz, M.; Tscherepanow, M.; van de Linde, S.; Heilemann, M.; Sauer, M. Real-time computation of subdiffraction-resolution fluorescence images. J. Microsc. 2010, 237, 12–22. [Google Scholar] [CrossRef]
- Wolter, S.; Löschberger, A.; Holm, T.; Aufmkolk, S.; Dabauvalle, M.C.; van de Linde, S.; Sauer, M. rapidSTORM: Accurate, fast open-source software for localization microscopy. Nat. Methods 2012, 9, 1040–1041. [Google Scholar] [CrossRef]
- van de Linde, S.; Löschberger, A.; Klein, T.; Heidbreder, M.; Wolter, S.; Heilemann, M.; Sauer, M. Direct stochastic optical reconstruction microscopy with standard fluorescent probes. Nat. Protoc. 2011, 6, 991–1009. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Endesfelder, U.; Malkusch, S.; Fricke, F.; Heilemann, M. A simple method to estimate the average localization precision of a single-molecule localization microscopy experiment. Histochem. Cell Biol. 2014, 141, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Malkusch, S.; Heilemann, M. Extracting quantitative information from single-molecule super-resolution imaging data with LAMA—LocAlization Microscopy Analyzer. Sci Rep 2016, 6, 34486. [Google Scholar] [CrossRef] [PubMed]
- McInnes, L.; Healy, J.; Astels, S. hdbscan: Hierarchical density based clustering. J. Open Source Softw. 2017, 2, 205. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mrestani, A.; Lichter, K.; Sirén, A.-L.; Heckmann, M.; Paul, M.M.; Pauli, M. Single-Molecule Localization Microscopy of Presynaptic Active Zones in Drosophila melanogaster after Rapid Cryofixation. Int. J. Mol. Sci. 2023, 24, 2128. https://doi.org/10.3390/ijms24032128
Mrestani A, Lichter K, Sirén A-L, Heckmann M, Paul MM, Pauli M. Single-Molecule Localization Microscopy of Presynaptic Active Zones in Drosophila melanogaster after Rapid Cryofixation. International Journal of Molecular Sciences. 2023; 24(3):2128. https://doi.org/10.3390/ijms24032128
Chicago/Turabian StyleMrestani, Achmed, Katharina Lichter, Anna-Leena Sirén, Manfred Heckmann, Mila M. Paul, and Martin Pauli. 2023. "Single-Molecule Localization Microscopy of Presynaptic Active Zones in Drosophila melanogaster after Rapid Cryofixation" International Journal of Molecular Sciences 24, no. 3: 2128. https://doi.org/10.3390/ijms24032128