
The Genetic Dissection of Nitrogen Use-Related Traits in Flax (Linum usitatissimum L.) at the Seedling Stage through the Integration of Multi-Locus GWAS, RNA-seq and Genomic Selection

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Variations in Root and Biomass Phenotypic Traits
2.2. Genetic Structure and Linkage Disequilibrium
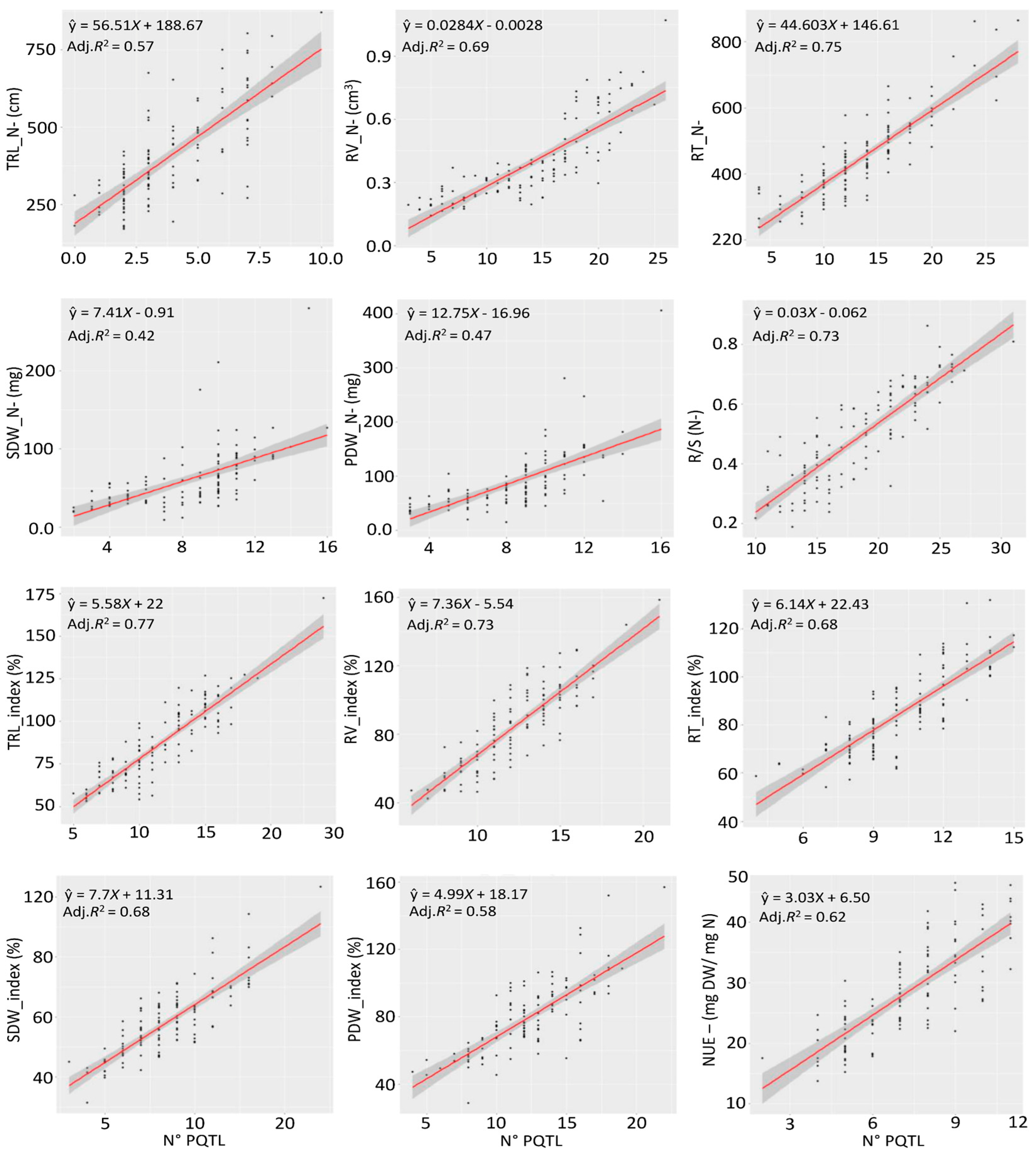
2.3. ML-GWAS of 21 NUE-Related Traits in Flax
2.4. Differentially Expressed Genes between High and Low NUE Genotypes
2.5. Differentially Expressed Candidate Genes at QTLs
2.6. GWAS-Assisted Genomic Selection
3. Discussion
3.1. Phenotypic Variation of Root and Biomass Traits
3.2. Genetic Structure and Linkage Disequilibrium
3.3. ML-GWAS of 21 NUE-Related Traits in Flax
3.4. Differentially Expressed Genes between High and Low NUE Genotypes
3.5. Differentially Expressed Genes at QTLs
3.6. GWAS-Assisted Genomic Selection
4. Materials and Methods
4.1. Plant Materials
4.2. Plant Growth Conditions and Phenotyping
4.3. Phenotypic Data Analysis
4.4. Genotyping, Genetic Structure and Linkage Disequilibrium
4.5. Multi-Locus Genome-Wide Association Analyses
4.6. Transcriptome Sequencing and Analysis
4.7. Cross-Referencing of Differentially Expressed Genes and QTL-Associated Candidate Genes
4.8. GWAS-Assisted Genomic Selection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qin, L.; Walk, T.C.; Han, P.; Chen, L.; Zhang, S.; Li, Y.; Hu, X.; Xie, L.; Yang, Y.; Liu, J.; et al. Adaption of Roots to Nitrogen Deficiency Revealed by 3D Quantification and Proteomic Analysis. Plant Physiol. 2019, 179, 329–347. [Google Scholar] [CrossRef] [PubMed]
- Shrawat, A.K.; Carroll, R.T.; DePauw, M.; Taylor, G.J.; Good, A.G. Genetic Engineering of Improved Nitrogen Use Efficiency in Rice by the Tissue-Specific Expression of Alanine Aminotransferase. Plant Biotechnol. J. 2008, 6, 722–732. [Google Scholar] [CrossRef] [PubMed]
- UNEP. Frontiers 2018/2019 Emerging Issues of Environmental Concern; United Nations Environment Programme: Nairobi, Kenya, 2019. [Google Scholar]
- Lei, L.; Li, G.; Zhang, H.; Powers, C.; Fang, T.; Chen, Y.; Wang, S.; Zhu, X.; Carver, B.F.; Yan, L. Nitrogen Use Efficiency Is Regulated by Interacting Proteins Relevant to Development in Wheat. Plant Biotechnol. J. 2018, 16, 1214–1226. [Google Scholar] [CrossRef] [PubMed]
- Billen, G.; Garnier, J.; Lassaletta, L. The Nitrogen Cascade from Agricultural Soils to the Sea: Modelling Nitrogen Transfers at Regional Watershed and Global Scales. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20130123. [Google Scholar] [CrossRef] [PubMed]
- Gruber, N.; Galloway, J.N. An Earth-System Perspective of the Global Nitrogen Cycle. Nature 2008, 451, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, A.S.; Laperche, A.; Bissuel-Belaygue, C.; Snowdon, R.; Nesi, N.; Stahl, A. Nitrogen Use Efficiency in Rapeseed. A Review. Agron. Sustain. Dev. 2016, 36, 38. [Google Scholar] [CrossRef]
- Moll, R.H.; Kamprath, E.J.; Jackson, W.A. Analysis and Interpretation of Factors Which Contribute to Efficiency of Nitrogen Utilization. Agron. J. 1982, 74, 562–564. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen Uptake, Assimilation and Remobilization in Plants: Challenges for Sustainable and Productive Agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef]
- Li, P.; Chen, F.; Cai, H.; Liu, J.; Pan, Q.; Liu, Z.; Gu, R.; Mi, G.; Zhang, F.; Yuan, L. A Genetic Relationship between Nitrogen Use Efficiency and Seedling Root Traits in Maize as Revealed by QTL Analysis. J. Exp. Bot. 2015, 66, 3175–3188. [Google Scholar] [CrossRef]
- Kant, S.; Bi, Y.M.; Rothstein, S.J. Understanding Plant Response to Nitrogen Limitation for the Improvement of Crop Nitrogen Use Efficiency. J. Exp. Bot. 2011, 62, 1499–1509. [Google Scholar] [CrossRef]
- Abenavoli, M.R.; Longo, C.; Lupini, A.; Miller, A.J.; Araniti, F.; Mercati, F.; Princi, M.P.; Sunseri, F. Phenotyping Two Tomato Genotypes with Different Nitrogen Use Efficiency. Plant Physiol. Biochem. 2016, 107, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Zhang, Y.; Xu, G. How Does Nitrogen Shape Plant Architecture? J. Exp. Bot. 2020, 71, 4415–4427. [Google Scholar] [CrossRef] [PubMed]
- Léran, S.; Muños, S.; Brachet, C.; Tillard, P.; Gojon, A.; Lacombe, B. Arabidopsis NRT1.1 Is a Bidirectional Transporter Involved in Root-to-Shoot Nitrate Translocation. Mol. Plant 2013, 6, 1984–1987. [Google Scholar] [CrossRef] [PubMed]
- Kanno, Y.; Hanada, A.; Chiba, Y.; Ichikawa, T.; Nakazawa, M.; Matsui, M.; Koshiba, T.; Kamiya, Y.; Seo, M. Identification of an Abscisic Acid Transporter by Functional Screening Using the Receptor Complex as a Sensor. Proc. Natl. Acad. Sci. USA 2012, 109, 9653–9658. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Fu, Y.L.; Pike, S.M.; Bao, J.; Tian, W.; Zhang, Y.; Chen, C.Z.; Zhang, Y.; Li, H.M.; Huang, J.; et al. The Arabidopsis Nitrate Transporter NRT1.8 Functions in Nitrate Removal from the Xylem Sap and Mediates Cadmium Tolerance. Plant Cell 2010, 22, 1633–1646. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Tsay, Y.F. Arabidopsis Nitrate Transporter NRT1.9 Is Important in Phloem Nitrate Transport. Plant Cell 2011, 23, 1945–1957. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.K.; Tsay, Y.F. Two Phloem Nitrate Transporters, NRT1.11 and NRT1.12, Are Important for Redistributing Xylem-Borne Nitrate to Enhance Plant Growth. Plant Physiol. 2013, 163, 844–856. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hu, B.; Chu, C. Nitrogen Use Efficiency in Crops: Lessons from Arabidopsis and Rice. J. Exp. Bot. 2017, 68, 2477–2488. [Google Scholar] [CrossRef]
- Hudson, D.; Guevara, D.; Yaish, M.W.; Hannam, C.; Long, N.; Clarke, J.D.; Bi, Y.M.; Rothstein, S.J. GNC and CGA1 Modulate Chlorophyll Biosynthesis and Glutamate Synthase (GLU1/FD-GOGAT) Expression in Arabidopsis. PLoS ONE 2011, 6, e26765. [Google Scholar] [CrossRef]
- Alvarez, J.M.; Riveras, E.; Vidal, E.A.; Gras, D.E.; Contreras-López, O.; Tamayo, K.P.; Aceituno, F.; Gómez, I.; Ruffel, S.; Lejay, L.; et al. Systems Approach Identifies TGA1 and TGA4 Transcription Factors as Important Regulatory Components of the Nitrate Response of Arabidopsis Thaliana Roots. Plant J. 2014, 80, 1–13. [Google Scholar] [CrossRef]
- Garnett, T.; Plett, D.; Heuer, S.; Okamoto, M. Genetic Approaches to Enhancing Nitrogen-Use Efficiency (NUE) in Cereals: Challenges and Future Directions. Funct. Plant Biol. 2015, 42, 921–941. [Google Scholar] [CrossRef]
- Cormier, F.; Foulkes, J.; Hirel, B.; Gouache, D.; Moënne-Loccoz, Y.; Le Gouis, J. Breeding for Increased Nitrogen-Use Efficiency: A Review for Wheat (T. aestivum L.). Plant Breed. 2016, 135, 255–278. [Google Scholar] [CrossRef]
- Plett, D.; Garnett, T.; Okamoto, M. Molecular Genetics to Discover and Improve Nitrogen Use Efficiency in Crop Plants. In Plant Macronutrient Use Efficiency: Molecular and Genomic Perspectives in Crop Plants; Elsevier: Amsterdam, The Netherlands, 2017; pp. 93–122. ISBN 9780128113080. [Google Scholar]
- Hirel, B.; Bertin, P.; Quilleré, I.; Bourdoncle, W.; Attagnant, C.; Dellay, C.; Gouy, A.; Cadiou, S.; Retailliau, C.; Falque, M.; et al. Towards a Better Understanding of the Genetic and Physiological Basis for Nitrogen Use Efficiency in Maize. Plant Physiol. 2001, 125, 1258–1270. [Google Scholar] [CrossRef]
- Kang, Y.; Sakiroglu, M.; Krom, N.; Stanton-Geddes, J.; Wang, M.; Lee, Y.C.; Young, N.D.; Udvardi, M. Genome-Wide Association of Drought-Related and Biomass Traits with HapMap SNPs in Medicago Truncatula. Plant Cell Environ. 2015, 38, 1997–2011. [Google Scholar] [CrossRef]
- Cormier, F.; Le Gouis, J.; Dubreuil, P.; Lafarge, S.; Praud, S. A Genome-Wide Identification of Chromosomal Regions Determining Nitrogen Use Efficiency Components in Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2014, 127, 2679–2693. [Google Scholar] [CrossRef]
- Monostori, I.; Szira, F.; Tondelli, A.; Árendás, T.; Gierczik, K.; Cattivelli, L.; Galiba, G.; VÁgújfalvi, A. Genome-Wide Association Study and Genetic Diversity Analysis on Nitrogen Use Efficiency in a Central European Winter Wheat (Triticum aestivum L.) Collection. PLoS ONE 2017, 12, e0189265. [Google Scholar] [CrossRef]
- Morosini, J.S.; Mendonça, L.d.F.; Lyra, D.H.; Galli, G.; Vidotti, M.S.; Fritsche-Neto, R. Association Mapping for Traits Related to Nitrogen Use Efficiency in Tropical Maize Lines under Field Conditions. Plant Soil 2017, 421, 453–463. [Google Scholar] [CrossRef]
- Gupta, N.; Gupta, M.; Akhatar, J.; Goyal, A.; Kaur, R.; Sharma, S.; Goyal, P.; Mukta, A.; Kaur, N.; Mittal, M.; et al. Association Genetics of the Parameters Related to Nitrogen Use Efficiency in Brassica juncea L. Plant Mol. Biol. 2021, 105, 161–175. [Google Scholar] [CrossRef]
- Ahmad, N.; Su, B.; Ibrahim, S.; Kuang, L.; Tian, Z.; Wang, X.; Wang, H.; Dun, X. Deciphering the Genetic Basis of Root and Biomass Traits in Rapeseed (Brassica napus L.) through the Integration of GWAS and RNA-Seq under Nitrogen Stress. Int. J. Mol. Sci. 2022, 23, 7958. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Li, C.; Zhang, X.; Li, Y.; Zhang, D.; Shi, Y.; Song, Y.; Li, Y.; Yang, D.; Wang, T. Transcriptome and GWAS Analyses Reveal Candidate Gene for Seminal Root Length of Maize Seedlings under Drought Stress. Plant Sci. 2020, 292, 110380. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, L.; Wang, L.; Tan, M.; Ogutu, C.O.; Yin, Z.; Zhou, J.; Wang, J.; Wang, L.; Yan, X. Transcriptome Analysis and Molecular Mechanism of Linseed (Linum usitatissimum L.) Drought Tolerance under Repeated Drought Using Single-Molecule Long-Read Sequencing. BMC Genom. 2021, 22, 109. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, X.; Zhang, Q.; Yuan, P.; Liu, L.; King, G.J.; Ding, G.; Wang, S.; Cai, H.; Wang, C.; et al. Integrating a Genome-Wide Association Study with Transcriptomic Data to Predict Candidate Genes and Favourable Haplotypes Influencing Brassica napus Seed Phytate. DNA Res. 2021, 28, dsab011. [Google Scholar] [CrossRef] [PubMed]
- Soto-Cerda, B.J.; Larama, G.; Gajardo, H.; Inostroza-Blancheteau, C.; Cloutier, S.; Fofana, B.; Abanto, M.; Aravena, G. Integrating Multi-Locus Genome-Wide Association Studies with Transcriptomic Data to Identify Genetic Loci Underlying Adult Root Trait Responses to Drought Stress in Flax (Linum usitatissimum L.). Environ. Exp. Bot. 2022, 202, 105019. [Google Scholar] [CrossRef]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of Total Genetic Value Using Genome-Wide Dense Marker Maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, Y.; Hu, Z.; Xu, C. Genomic Selection Methods for Crop Improvement: Current Status and Prospects. Crop. J. 2018, 6, 330–340. [Google Scholar] [CrossRef]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Bi, I.V.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A Unified Mixed-Model Method for Association Mapping That Accounts for Multiple Levels of Relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef]
- Spindel, J.E.; Begum, H.; Akdemir, D.; Collard, B.; Redoña, E.; Jannink, J.L.; McCouch, S. Genome-Wide Prediction Models That Incorporate de Novo GWAS Are a Powerful New Tool for Tropical Rice Improvement. Heredity 2016, 116, 395–408. [Google Scholar] [CrossRef]
- Minamikawa, M.F.; Nonaka, K.; Kaminuma, E.; Kajiya-Kanegae, H.; Onogi, A.; Goto, S.; Yoshioka, T.; Imai, A.; Hamada, H.; Hayashi, T.; et al. Genome-Wide Association Study and Genomic Prediction in Citrus: Potential of Genomics-Assisted Breeding for Fruit Quality Traits. Sci. Rep. 2017, 7, 4721. [Google Scholar] [CrossRef]
- Lan, S.; Zheng, C.; Hauck, K.; McCausland, M.; Duguid, S.D.; Booker, H.M.; Cloutier, S.; You, F.M. Genomic Prediction Accuracy of Seven Breeding Selection Traits Improved by QTL Identification in Flax. Int. J. Mol. Sci. 2020, 21, 1577. [Google Scholar] [CrossRef]
- He, H.; Yang, R.; Li, Y.; Ma, A.; Cao, L.; Wu, X.; Chen, B.; Tian, H.; Gao, Y. Genotypic Variation in Nitrogen Utilization Efficiency of Oilseed Rape (Brassica napus) under Contrasting N Supply in Pot and Field Experiments. Front. Plant Sci. 2017, 8, 1825. [Google Scholar] [CrossRef]
- Dash, P.K.; Cao, Y.; Jailani, A.K.; Gupta, P.; Venglat, P.; Xiang, D.; Rai, R.; Sharma, R.; Thirunavukkarasu, N.; Abdin, M.Z.; et al. Genome-Wide Analysis of Drought Induced Gene Expression Changes in Flax (Linum usitatissimum). GM Crop. Food 2014, 5, 106–119. [Google Scholar] [CrossRef]
- Rabetafika, H.N.; Van Remoortel, V.; Danthine, S.; Paquot, M.; Blecker, C. Flaxseed Proteins: Food Uses and Health Benefits. Int. J. Food Sci. Technol. 2011, 46, 221–228. [Google Scholar] [CrossRef]
- Soto-Cerda, B.J.; Cloutier, S.; Gajardo, H.A.; Aravena, G.; Quian, R. Identifying Drought-Resilient Flax Genotypes and Related-Candidate Genes Based on Stress Indices, Root Traits and Selective Sweep. Euphytica 2019, 215, 41. [Google Scholar] [CrossRef]
- Dordas, C.A. Nitrogen Nutrition Index and Its Relationship to N Use Efficiency in Linseed. Eur. J. Agron. 2011, 34, 124–132. [Google Scholar] [CrossRef]
- Xie, Y.; Gan, Y.; Li, Y.; Niu, J.; Gao, Y.; An, H.; Li, A. Effect of Nitrogen Fertilizer on Nitrogen Accumulation, Translocation, and Use Efficiency in Dryland Oilseed Flax. Agron. J. 2015, 107, 1931–1939. [Google Scholar] [CrossRef]
- You, F.M.; Xiao, J.; Li, P.; Yao, Z.; Jia, G.; He, L.; Zhu, T.; Luo, M.C.; Wang, X.; Deyholos, M.K.; et al. Chromosome-Scale Pseudomolecules Refined by Optical, Physical and Genetic Maps in Flax. Plant J. 2018, 95, 371–384. [Google Scholar] [CrossRef]
- Brooks, M.D.; Cirrone, J.; Pasquino, A.V.; Alvarez, J.M.; Swift, J.; Mittal, S.; Juang, C.L.; Varala, K.; Gutiérrez, R.A.; Krouk, G.; et al. Network Walking Charts Transcriptional Dynamics of Nitrogen Signaling by Integrating Validated and Predicted Genome-Wide Interactions. Nat. Commun. 2019, 10, 1569. [Google Scholar] [CrossRef]
- Xu, N.; Wang, R.; Zhao, L.; Zhang, C.; Li, Z.; Lei, Z.; Liu, F.; Guan, P.; Chu, Z.; Crawford, N.M. The Arabidopsis NRG2 Protein Mediates Nitrate Signaling and Interacts with and Regulates Key Nitrate Regulators. Plant Cell 2015, 28, 485–504. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.H.; Kan, C.C.; Wu, H.Y.; Yang, H.C.; Hsieh, M.H. Early Molecular Events Associated with Nitrogen Deficiency in Rice Seedling Roots. Sci. Rep. 2018, 8, 12207. [Google Scholar] [CrossRef] [PubMed]
- Bouly, J.P.; Gissot, L.; Lessard, P.; Kreis, M.; Thomas, M. Arabidopsis Thaliana Proteins Related to the Yeast SIP and SNF4 Interact with AKINα1, an SNF1-like Protein Kinase. Plant J. 1999, 18, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, M.; Georgescu, M.N.; Takahashi, M. A Nitrite Transporter Associated with Nitrite Uptake by Higher Plant Chloroplasts. Plant Cell Physiol. 2007, 48, 1022–1035. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Tischner, R.; Gutiérrez, R.A.; Hoffman, M.; Xing, X.; Chen, M.; Coruzzi, G.; Crawford, N.M. Genomic Analysis of the Nitrate Response Using a Nitrate Reductase-Null Mutant of Arabidopsis. Plant Physiol. 2004, 136, 2512–2522. [Google Scholar] [CrossRef] [PubMed]
- Sorger, G.; Gooden, D.O.; Earle, E.D.; Mckinnon, J. NADH Nitrate Reductase and NAD(P)H Nitrate Reductase in Genetic Variants and Regenerating Callus of Maize. Plant Physiol. 1986, 82, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Funayama, K.; Kojima, S.; Tabuchi-Kobayashi, M.; Sawa, Y.; Nakayama, Y.; Hayakawa, T.; Yamaya, T. Cytosolic Glutamine Synthetase1;2 Is Responsible for the Primary Assimilation of Ammonium in Rice Roots. Plant Cell Physiol. 2013, 54, 934–943. [Google Scholar] [CrossRef]
- Delay, C.; Imin, N.; Djordjevic, M.A. CEP Genes Regulate Root and Shoot Development in Response to Environmental Cues and Are Specific to Seed Plants. J. Exp. Bot. 2013, 64, 5383–5394. [Google Scholar] [CrossRef] [PubMed]
- Remans, T.; Nacry, P.; Pervent, M.; Filleur, S.; Diatloff, E.; Mounier, E.; Tillard, P.; Forde, B.G.; Gojon, A. The Arabidopsis NRT1.1 Transporter Participates in the Signaling Pathway Triggering Root Colonization of Nitrate-Rich Patches. Proc. Natl. Acad. Sci. USA 2006, 103, 19206–19211. [Google Scholar] [CrossRef] [PubMed]
- Kotur, Z.; Glass, A.D.M. A 150kDa Plasma Membrane Complex of AtNRT2.5 and AtNAR2.1 Is the Major Contributor to Constitutive High-Affinity Nitrate Influx in Arabidopsis Thaliana. Plant Cell Environ. 2015, 38, 1490–1502. [Google Scholar] [CrossRef]
- Kanno, Y.; Kamiya, Y.; Seo, M. Nitrate Does Not Compete with Abscisic Acid as a Substrate of AtNPF4.6/NRT1.2/AIT1 in Arabidopsis. Plant Signal. Behav. 2013, 8, e26624. [Google Scholar] [CrossRef]
- Gazzarrini, S.; Lejay, L.; Gojon, A.; Ninnemann, O.; Frommer, W.B.; Von Wirén, N. Three Functional Transporters for Constitutive, Diurnally Regulated, and Starvation-Induced Uptake of Ammonium into Arabidopsis Roots. Plant Cell 1999, 11, 937–947. [Google Scholar] [CrossRef]
- Park, C.H.; Roh, J.; Youn, J.H.; Son, S.H.; Park, J.H.; Kim, S.Y.; Kim, T.W.; Kim, S.K. Arabidopsis ACC Oxidase 1 Coordinated by Multiple Signals Mediates Ethylene Biosynthesis and Is Involved in Root Development. Mol. Cells 2018, 41, 923–932. [Google Scholar] [CrossRef]
- Racolta, A.; Bryan, A.C.; Tax, F.E. The Receptor-like Kinases GSO1 and GSO2 Together Regulate Root Growth in Arabidopsis through Control of Cell Division and Cell Fate Specification. Dev. Dyn. 2014, 243, 257–278. [Google Scholar] [CrossRef] [PubMed]
- Seifert, G.J.; Xue, H.; Acet, T. The Arabidopsis Thaliana FASCICLIN LIKE ARABINOGALACTAN PROTEIN 4 Gene Acts Synergistically with Abscisic Acid Signalling to Control Root Growth. Ann. Bot. 2014, 114, 1125–1133. [Google Scholar] [CrossRef] [PubMed]
- Devaiah, B.N.; Karthikeyan, A.S.; Raghothama, K.G. WRKY75 Transcription Factor Is a Modulator of Phosphate Acquisition and Root Development in Arabidopsis. Plant Physiol. 2007, 143, 1789–1801. [Google Scholar] [CrossRef]
- Xue, X.H.; Guo, C.Q.; Du, F.; Lu, Q.L.; Zhang, C.M.; Ren, H.Y. AtFH8 Is Involved in Root Development under Effect of Low-Dose Latrunculin B in Dividing Cells. Mol. Plant 2011, 4, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Liberman, L.M.; Sparks, E.E.; Moreno-Risueno, M.A.; Petricka, J.J.; Benfey, P.N. MYB36 Regulates the Transition from Proliferation to Differentiation in the Arabidopsis Root. Proc. Natl. Acad. Sci. USA 2015, 112, 12099–12104. [Google Scholar] [CrossRef] [PubMed]
- Van Leene, J.; Blomme, J.; Kulkarni, S.R.; Cannoot, B.; De Winne, N.; Eeckhout, D.; Persiau, G.; Van De Slijke, E.; Vercruysse, L.; Vanden Bossche, R.; et al. Functional Characterization of the Arabidopsis Transcription Factor BZIP29 Reveals Its Role in Leaf and Root Development. J. Exp. Bot. 2016, 67, 5825–5840. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Frugis, G.; Colgan, D.; Chua, N.H. Arabidopsis NAC1 Transduces Auxin Signal Downstream of TIR1 to Promote Lateral Root Development. Genes Dev. 2000, 14, 3024–3036. [Google Scholar] [CrossRef]
- Moriwaki, T.; Miyazawa, Y.; Kobayashi, A.; Uchida, M.; Watanabe, C.; Fujii, N.; Takahashi, H. Hormonal Regulation of Lateral Root Development in Arabidopsis Modulated by MIZ1 and Requirement of GNOM Activity for MIZ1 Function. Plant Physiol. 2011, 157, 1209–1220. [Google Scholar] [CrossRef]
- Tabata, R.; Sumida, K.; Yoshii, T.; Ohyama, K.; Shinohara, H.; Matsubayashi, Y. Perception of Root-Derived Peptides by Shoot LRR-RKs Mediates Systemic N-Demand Signaling. Science 2014, 346, 343–346. [Google Scholar] [CrossRef]
- Shin, R.; Burch, A.Y.; Huppert, K.A.; Tiwari, S.B.; Murphy, A.S.; Guilfoyle, T.J.; Schachtman, D.P. The Arabidopsis Transcription Factor MYB77 Modulates Auxin Signal Transduction. Plant Cell 2007, 19, 2440–2453. [Google Scholar] [CrossRef]
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. ARF7 and ARF19 Regulate Lateral Root Formation via Direct Activation of LBD/ASL Genes in Arabidopsis. Plant Cell 2007, 19, 118–130. [Google Scholar] [CrossRef] [PubMed]
- De Smet, I.; Vassileva, V.; De Rybel, B.; Levesque, M.P.; Grunewald, W.; Van Damme, D.; Van Noorden, G.; Naudts, M.; Van Isterdael, G.; De Clercq, R.; et al. Receptor-Like Kinase ACR4 Formative Cell Divisions in the Arabidopsis Root. Science 2008, 322, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Watt, M.; Mathesius, U. The Autoregulation Gene SUNN Mediates Changes in Root Organ Formation in Response to Nitrogen through Alteration of Shoot-to-Root Auxin Transport. Plant Physiol. 2012, 159, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Lewis, D.R.; Spalding, E.P. Mutations in Arabidopsis Multidrug Resistance-Like ABC Transporters Separate the Roles of Acropetal and Basipetal Auxin Transport in Lateral Root Development. Plant Cell 2007, 19, 1826–1837. [Google Scholar] [CrossRef] [PubMed]
- Santelia, D.; Vincenzetti, V.; Azzarello, E.; Bovet, L.; Fukao, Y.; Düchtig, P.; Mancuso, S.; Martinoia, E.; Geisler, M. MDR-like ABC Transporter AtPGP4 Is Involved in Auxin-Mediated Lateral Root and Root Hair Development. FEBS Lett. 2005, 579, 5399–5406. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.H.; Frommer, W.B.; Ludewig, U. Molecular and Functional Characterization of a Family of Amino Acid Transporters from Arabidopsis. Plant Physiol. 2004, 136, 3104–3113. [Google Scholar] [CrossRef]
- Joshi, N.C.; Meyer, A.J.; Bangash, S.A.K.; Zheng, Z.L.; Leustek, T. Arabidopsis γ-Glutamylcyclotransferase Affects Glutathione Content and Root System Architecture during Sulfur Starvation. New Phytol. 2019, 221, 1387–1397. [Google Scholar] [CrossRef]
- Dündar, E.; Bush, D.R. BAT1, a Bidirectional Amino Acid Transporter in Arabidopsis. Planta 2009, 229, 1047–1056. [Google Scholar] [CrossRef]
- Russnak, R.; Konczal, D.; McIntire, S.L. A Family of Yeast Proteins Mediating Bidirectional Vacuolar Amino Acid Transport. J. Biol. Chem. 2001, 276, 23849–23857. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, W.; Yang, Y.; Li, Z.; Li, N.; Qi, S.; Crawford, N.M.; Wang, Y. The Arabidopsis NLP7 Gene Regulates Nitrate Signaling via NRT1.1-Dependent Pathway in the Presence of Ammonium. Sci. Rep. 2018, 8, 1487. [Google Scholar] [CrossRef]
- Fischer, W.; Kwart, M.; Hummel, S.; Frommer, W.B. Substrate Specificity and Expression Profile of Amino Acid Transporters (AAPs) in Arabidopsis. J. Biol. Chem. 1995, 270, 16315–16320. [Google Scholar] [CrossRef] [PubMed]
- Planchais, S.; Cabassa, C.; Toka, I.; Justin, A.M.; Renou, J.P.; Savouré, A.; Carol, P. BASIC AMINO ACID CARRIER 2 Gene Expression Modulates Arginine and Urea Content and Stress Recovery in Arabidopsis Leaves. Front. Plant Sci. 2014, 5, 330. [Google Scholar] [CrossRef] [PubMed]
- Pratelli, R.; Voll, L.M.; Horst, R.J.; Frommer, W.B.; Pilot, G. Stimulation of Nonselective Amino Acid Export by Glutamine Dumper Proteins. Plant Physiol. 2010, 152, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Diederichsen, A.; Kusters, P.M.; Kessler, D.; Bainas, Z.; Gugel, R.K. Assembling a Core Collection from the Flax World Collection Maintained by Plant Gene Resources of Canada. Genet. Resour. Crop. Evol. 2013, 60, 1479–1485. [Google Scholar] [CrossRef]
- Sertse, D.; You, F.M.; Ravichandran, S.; Soto-Cerda, B.J.; Duguid, S.; Cloutier, S. Loci Harboring Genes with Important Role in Drought and Related Abiotic Stress Responses in Flax Revealed by Multiple GWAS Models. Theor. Appl. Genet. 2021, 134, 191–212. [Google Scholar] [CrossRef]
- Sertse, D.; You, F.M.; Ravichandran, S.; Cloutier, S. The Complex Genetic Architecture of Early Root and Shoot Traits in Flax Revealed by Genome-Wide Association Analyses. Front. Plant Sci. 2019, 10, 1483. [Google Scholar] [CrossRef]
- Chen, J.; Liu, L.; Wang, Z.; Zhang, Y.; Sun, H.; Song, S.; Bai, Z.; Lu, Z.; Li, C. Nitrogen Fertilization Increases Root Growth and Coordinates the Root–Shoot Relationship in Cotton. Front. Plant Sci. 2020, 11, 880. [Google Scholar] [CrossRef]
- Myles, S.; Peiffer, J.; Brown, P.J.; Ersoz, E.S.; Zhang, Z.; Costich, D.E.; Buckler, E. Association Mapping: Critical Considerations Shift from Genotyping to Experimental Design. Plant Cell 2009, 21, 2194–2202. [Google Scholar] [CrossRef]
- Shi, H.; Chen, M.; Gao, L.; Wang, Y.; Bai, Y.; Yan, H.; Xu, C.; Zhou, Y.; Xu, Z.; Chen, J.; et al. Genome-Wide Association Study of Agronomic Traits Related to Nitrogen Use Efficiency in Wheat. Theor. Appl. Genet. 2022, 135, 4289–4302. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, J.; Xia, Z.; Wang, Q.; Zhang, S.; Zhang, G.; Lu, H. Genome-Wide Association Studies of Maize Seedling Root Traits under Different Nitrogen Levels. Plants 2022, 11, 1417. [Google Scholar] [CrossRef]
- Karunarathne, S.D.; Han, Y.; Zhang, X.Q.; Zhou, G.; Hill, C.B.; Chen, K.; Angessa, T.; Li, C. Genome-Wide Association Study and Identification of Candidate Genes for Nitrogen Use Efficiency in Barley (Hordeum vulgare L.). Front. Plant Sci. 2020, 11, 571912. [Google Scholar] [CrossRef] [PubMed]
- Soto-Cerda, B.J.; Cloutier, S.; Quian, R.; Gajardo, H.A.; Olivos, M.; You, F.M. Genome-Wide Association Analysis of Mucilage and Hull Content in Flax (Linum usitatissimum L.) Seeds. Int. J. Mol. Sci. 2018, 19, 2870. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Xiao, J.; Rashid, K.Y.; Yao, Z.; Li, P.; Jia, G.; Wang, X.; Cloutier, S.; You, F.M. Genome-Wide Association Studies for Pasmo Resistance in Flax (Linum usitatissimum L.). Front. Plant Sci. 2019, 9, 1982. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Dai, Z.; Yang, Z.; Tang, Q.; Sun, J.; Yang, X.; Song, X.; Lu, Y.; Zhao, D.; Zhang, L.; et al. Genomic Variations and Association Study of Agronomic Traits in Flax. BMC Genom. 2018, 19, 512. [Google Scholar] [CrossRef]
- You, F.M.; Xiao, J.; Li, P.; Yao, Z.; Jia, G.; He, L.; Kumar, S.; Soto-Cerda, B.; Duguid, S.D.; Booker, H.M.; et al. Genome-Wide Association Study and Selection Signatures Detect Genomic Regions Associated with Seed Yield and Oil Quality in Flax. Int. J. Mol. Sci. 2018, 19, 2303. [Google Scholar] [CrossRef]
- Soto-Cerda, B.J.; Cloutier, S.; Gajardo, H.A.; Aravena, G.; Quian, R.; You, F.M. Drought Response of Flax Accessions and Identification of Quantitative Trait Nucleotides (QTNs) Governing Agronomic and Root Traits by Genome-Wide Association Analysis. Mol. Breed. 2020, 40, 1–24. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Jia, Z.; Dunwell, J.M. Editorial: The Applications of New Multi-Locus Gwas Methodologies in the Genetic Dissection of Complex Traits. Front. Plant Sci. 2019, 10, 100. [Google Scholar] [CrossRef]
- Soto-Cerda, B.J.; Aravena, G.; Cloutier, S. Genetic Dissection of Flowering Time in Flax (Linum usitatissimum L.) through Single- and Multi-Locus Genome-Wide Association Studies. Mol. Genet. Genom. 2021, 296, 877–891. [Google Scholar] [CrossRef]
- You, F.M.; Rashid, K.Y.; Zheng, C.; Khan, N.; Li, P.; Xiao, J.; He, L.; Yao, Z.; Cloutier, S. Insights into the Genetic Architecture and Genomic Prediction of Powdery Mildew Resistance in Flax (Linum usitatissimum L.). Int. J. Mol. Sci. 2022, 23, 4960. [Google Scholar] [CrossRef]
- Fan, X.; Zhang, W.; Zhang, N.; Chen, M.; Zheng, S.; Zhao, C.; Han, J.; Liu, J.; Zhang, X.; Song, L.; et al. Identification of QTL Regions for Seedling Root Traits and Their Effect on Nitrogen Use Efficiency in Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2018, 131, 2677–2698. [Google Scholar] [CrossRef]
- Zhu, J.; Kaeppler, S.M.; Lynch, J.P. Mapping of QTLs for Lateral Root Branching and Length in Maize (Zea mays L.) under Differential Phosphorus Supply. Theor. Appl. Genet. 2005, 111, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, J.K.; Buckseth, T.; Zinta, R.; Saraswati, A.; Singh, R.K.; Rawat, S.; Dua, V.K.; Chakrabarti, S.K. Transcriptome Analysis of Potato Shoots, Roots and Stolons under Nitrogen Stress. Sci. Rep. 2020, 10, 1152. [Google Scholar] [CrossRef] [PubMed]
- Xin, W.; Zhang, L.; Gao, J.; Zhang, W.; Yi, J.; Zhen, X.; Bi, C.; He, D.; Liu, S.; Zhao, X. Adaptation Mechanism of Roots to Low and High Nitrogen Revealed by Proteomic Analysis. Rice 2021, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, A.; Dong, Q.; Wang, X.; Gui, H.; Zhang, H.; Zhang, X.; Song, M. Transcriptome Analysis Reveals Differences in Key Genes and Pathways Regulating Carbon and Nitrogen Metabolism in Cotton Genotypes under n Starvation and Resupply. Int. J. Mol. Sci. 2020, 21, 1500. [Google Scholar] [CrossRef]
- Goel, P.; Sharma, N.K.; Bhuria, M.; Sharma, V.; Chauhan, R.; Pathania, S.; Swarnkar, M.K.; Chawla, V.; Acharya, V.; Shankar, R.; et al. Transcriptome and Co-Expression Network Analyses Identify Key Genes Regulating Nitrogen Use Efficiency in Brassica Juncea L. Sci. Rep. 2018, 8, 7451. [Google Scholar] [CrossRef]
- Liu, S.H.; Shen, P.C.; Chen, C.Y.; Hsu, A.N.; Cho, Y.C.; Lai, Y.L.; Chen, F.H.; Li, C.Y.; Wang, S.C.; Chen, M.; et al. DriverDBv3: A Multi-Omics Database for Cancer Driver Gene Research. Nucleic Acids Res. 2020, 48, D863–D870. [Google Scholar] [CrossRef]
- Wu, Y.; Shi, H.; Yu, H.; Ma, Y.; Hu, H.; Han, Z.; Zhang, Y.; Zhen, Z.; Yi, L.; Hou, J. Combined GWAS and Transcriptome Analyses Provide New Insights Into the Response Mechanisms of Sunflower Against Drought Stress. Front. Plant Sci. 2022, 13, 847435. [Google Scholar] [CrossRef]
- He, F.; Zhang, F.; Jiang, X.; Long, R.; Wang, Z.; Chen, Y.; Li, M.; Gao, T.; Yang, T.; Wang, C.; et al. A Genome-Wide Association Study Coupled With a Transcriptomic Analysis Reveals the Genetic Loci and Candidate Genes Governing the Flowering Time in Alfalfa (Medicago sativa L.). Front. Plant Sci. 2022, 13, 913947. [Google Scholar] [CrossRef]
- Wei, Z.; Yuan, Q.; Lin, H.; Li, X.; Zhang, C.; Gao, H.; Zhang, B.; He, H.; Liu, T.; Jie, Z.; et al. Linkage Analysis, GWAS, Transcriptome Analysis to Identify Candidate Genes for Rice Seedlings in Response to High Temperature Stress. BMC Plant Biol. 2021, 21, 85. [Google Scholar] [CrossRef]
- Wong, C.K.; Bernardo, R. Genomewide Selection in Oil Palm: Increasing Selection Gain per Unit Time and Cost with Small Populations. Theor. Appl. Genet. 2008, 116, 815–824. [Google Scholar] [CrossRef]
- Lorenzana, R.E.; Bernardo, R. Accuracy of Genotypic Value Predictions for Marker-Based Selection in Biparental Plant Populations. Theor. Appl. Genet. 2009, 120, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Hosseini-Vardanjani, S.M.; Shariati, M.M.; Shahrebabak, H.M.; Tahmoorespur, M. Incorporating Prior Knowledge of Principal Components in Genomic Prediction. Front. Genet. 2018, 9, 289. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Cairns, J.E.; Babu, R.; Gowda, M.; Makumbi, D.; Magorokosho, C.; Zhang, A.; Liu, Y.; Wang, N.; Hao, Z.F.; et al. Genome-Wide Association Mapping and Genomic Prediction Analyses Reveal the Genetic Architecture of Grain Yield and Flowering Time under Drought and Heat Stress Conditions in Maize. Front. Plant Sci. 2019, 9, 1919. [Google Scholar] [CrossRef] [PubMed]
- Zhang-Biehn, S.; Fritz, A.K.; Zhang, G.; Evers, B.; Regan, R.; Poland, J. Accelerating Wheat Breeding for End-Use Quality through Association Mapping and Multivariate Genomic Prediction. Plant Genome 2021, 14, e20164. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Huerta, R.F.; Guevara-Gonzalez, R.G.; Contreras-Medina, L.M.; Torres-Pacheco, I.; Prado-Olivarez, J.; Ocampo-Velazquez, R.V. A Review of Methods for Sensing the Nitrogen Status in Plants: Advantages, Disadvantages and Recent Advances. Sensors 2013, 13, 10823–10843. [Google Scholar] [CrossRef] [PubMed]
- VSN International GenStat for Windows 2015. Available online: https://vsni.co.uk/software/genstat (accessed on 7 January 2023).
- Bouslama, M.; Schapaugh, W.T. Stress Tolerance in Soybeans. I. Evaluation of Three Screening Techniques for Heat and Drought Tolerance. Crop. Sci. 1984, 24, 933–937. [Google Scholar] [CrossRef]
- Fernandez, G.C.J. Effective Selection Criteria for Assessing Plant Stress Tolerance; Kuo, C.G., Ed.; International Symposium on Adaptation of Food Crops to Temperature and Water Stress: Tainan, Taiwan, 1992. [Google Scholar]
- Wickham, H. Ggplot2-Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Rosenberg, N.A.; Donnelly, P. Association Mapping in Structured Populations. Am. J. Hum. Genet. 2000, 67, 170–181. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software STRUCTURE: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for Association Mapping of Complex Traits in Diverse Samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Wen, Y.J.; Zhang, H.; Ni, Y.L.; Huang, B.; Zhang, J.; Feng, J.Y.; Wang, S.B.; Dunwell, J.M.; Zhang, Y.M.; Wu, R. Methodological Implementation of Mixed Linear Models in Multi-Locus Genome-Wide Association Studies. Brief Bioinform. 2018, 19, 700–712. [Google Scholar] [CrossRef]
- Tamba, C.L.; Zhang, Y.-M. A Fast MrMLM Algorithm for Multi-Locus Genome-Wide Association Studies. bioRxiv 2018. [Google Scholar] [CrossRef]
- Tamba, C.L.; Ni, Y.L.; Zhang, Y.M. Iterative Sure Independence Screening EM-Bayesian LASSO Algorithm for Multi-Locus Genome-Wide Association Studies. PLoS Comput. Biol. 2017, 13, e1005357. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.B.; Feng, J.Y.; Ren, W.L.; Huang, B.; Zhou, L.; Wen, Y.J.; Zhang, J.; Dunwell, J.M.; Xu, S.; Zhang, Y.M. Improving Power and Accuracy of Genome-Wide Association Studies via a Multi-Locus Mixed Linear Model Methodology. Sci. Rep. 2016, 6, 19444. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.L.; Wen, Y.J.; Dunwell, J.M.; Zhang, Y.M. PKWmEB: Integration of Kruskal-Wallis Test with Empirical Bayes under Polygenic Background Control for Multi-Locus Genome-Wide Association Study. Heredity 2018, 120, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Feng, J.Y.; Ni, Y.L.; Wen, Y.J.; Niu, Y.; Tamba, C.L.; Yue, C.; Song, Q.; Zhang, Y.M. PLARmEB: Integration of Least Angle Regression with Empirical Bayes for Multilocus Genome-Wide Association Studies. Heredity 2017, 118, 517–524. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Tamba, C.L.; Wen, Y.J.; Li, P.; Ren, W.L.; Ni, Y.L.; Gao, J.; Zhang, Y.M. MrMLM v4.0.2: An R Platform for Multi-Locus Genome-Wide Association Studies. Genom. Proteom. Bioinform. 2020, 18, 481–487. [Google Scholar] [CrossRef]
- Vos, P.G.; Paulo, M.J.; Voorrips, R.E.; Visser, R.G.F.; van Eck, H.J.; van Eeuwijk, F.A. Evaluation of LD Decay and Various LD-Decay Estimators in Simulated and SNP-Array Data of Tetraploid Potato. Theor. Appl. Genet. 2017, 130, 123–135. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Aramaki, T.; Blanc-Mathieu, R.; Endo, H.; Ohkubo, K.; Kanehisa, M.; Goto, S.; Ogata, H. KofamKOALA: KEGG Ortholog Assignment Based on Profile HMM and Adaptive Score Threshold. Bioinformatics 2020, 36, 2251–2252. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.B.; Dias, K.O.G.; Ferreira, D.F.; Brown, P.J. Efficiency of Multi-Trait, Indirect, and Trait-Assisted Genomic Selection for Improvement of Biomass Sorghum. Theor. Appl. Genet. 2018, 131, 747–755. [Google Scholar] [CrossRef]
- Pérez, P.; De Los Campos, G. Genome-Wide Regression and Prediction with the BGLR Statistical Package. Genetics 2014, 198, 483–495. [Google Scholar] [CrossRef]
- De Los Campos, G.; Pérez, P.; Vazquez, A.I.; Crossa, J. Genome-Enabled Prediction Using the BLR (Bayesian Linear Regression) R-Package. Methods Mol. Biol. 2013, 1019, 299–320. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Mean | Standard Deviation | Range | C.V. (%) |
|---|---|---|---|---|
| Total root length N+ (cm) | 469.2 | 137.7 | 168.6–882.0 | 29.2 |
| Total root length N− (cm) | 407.2 | 156.8 | 171.0–870.9 | 38.3 |
| Root volume N+ (cm3) | 0.464 | 0.181 | 0.161–1.056 | 38.7 |
| Root volume N− (cm3) | 0.389 | 0.188 | 0.143–1.069 | 48.1 |
| Number of root tips N+ | 533.7 | 120.7 | 292.4–1060.4 | 22.5 |
| Number of root tips N− | 447.8 | 125.1 | 236.6–864.5 | 27.8 |
| Plant dry weight N+ (mg) | 105.7 | 52.9 | 11.8–390.9 | 49.8 |
| Plant dry weight N− (mg) | 86.4 | 53.3 | 6.8–405.8 | 61.5 |
| Shoot dry weight N+ (mg) | 77.1 | 41.3 | 9.1–290.6 | 53.3 |
| Shoot dry weight N− (mg) | 59.9 | 38.5 | 4.6–280.0 | 64.0 |
| Root/shoot N+ | 0.407 | 0.116 | 0.215–0.784 | 28.3 |
| Root/shoot N− | 0.490 | 0.158 | 0.188–0.862 | 32.0 |
| TRL_Index (%) | 87.3 | 21.6 | 53.1–172.4 | 24.7 |
| RV_Index (%) | 85.1 | 23.7 | 42.4–158.4 | 27.7 |
| RT_Index (%) | 84.6 | 16.9 | 53.9–131.8 | 19.9 |
| PDW_Index (%) | 80.1 | 21.1 | 28.7–156.9 | 26.2 |
| SDW_Index (%) | 76.1 | 22.9 | 22.9–166.7 | 30.0 |
| NUE N+ | 25.6 | 7.2 | 10.4–37.2 | 28.0 |
| NUE N− | 28.3 | 7.8 | 13.7–46.5 | 27.5 |
| NUE_Index (%) | 114.6 | 25.0 | 59.7–204.7 | 21.7 |
| NUE_STI | 1.17 | 0.571 | 0.23–2.55 | 48.5 |
| Flax DEG | Gene Name | Protein Name | QTL | Trait | Function | Up/Down (Log2FC) | Reference |
|---|---|---|---|---|---|---|---|
| Lus10027573 | CRF4 | Ethylene-responsive transcription factor CRF4 | Not in QTL | Unknown | N signaling | Up LN (1.20) | [49] |
| Lus10029683 | NRG2 | Nitrate regulatory gene2 protein | Lu9_18443162 | RV_Index | N signaling | Down LN (−1.33) | [50] |
| Lus10019292 | LBD38 | LOB domain-containing protein 38 | Not in QTL | Unknown | NO3− response regulation | Up LN (1.27) | [51] |
| Lus10038783 | KINB2 | SNF1-related protein kinase regulatory subunit beta-2 | Lu11_1841121 | NUE_N−, NUE_N+, NUE_STI | NO3− assimilation | Down LN (−1.32) | [52] |
| Lus10041466 | NPF3.1 | Protein NRT1/PTR FAMILY 3.1 | Lu4_14298191 | R/S_N− | NO3− assimilation | Up LN (1.49) | [53] |
| Lus10035402 | NIA | Nitrate reductase [NADH] | Not in QTL | Unknown | NO3− assimilation | Down LN (−1.26) | [54] |
| Lus10027270 | NR | Nitrate reductase [NADH] | Not in QTL | Unknown | NO3− assimilation | Down LN (−1.46) | [55] |
| Lus10004037 | GS1-2 | Glutamine synthetase cytosolic isozyme 2 | Not in QTL | Unknown | NH4+ assimilation | Down LN (−1.37), Down HN (−1.03) | [56] |
| Lus10029256 | CEP14 | Precursor of CEP14 | Not in QTL | Unknown | NH4+ assimilation, root development | Down LN (−1.16) | [57] |
| Lus10016120 | NRT2.1 | High-affinity nitrate transporter 2.1 | Not in QTL | Unknown | NO3− transporter | Down LN (−1.17) | [58] |
| Lus10030902 | NRT2.5 | High affinity nitrate transporter 2.5 | Lu13_18363934 | RT_Index, R/S_N+ | NO3− transporter | Down LN (−2.38), Down HN (−1.83) | [59] |
| Lus10014537 | NPF1.1 | Protein NRT1/PTR FAMILY 1.1 | Not in QTL | Unknown | NO3− transporter | Up LN (1.12) | [18] |
| Lus10032876 | NPF4.6 | Protein NRT1/PTR FAMILY 4.6 | Not in QTL | Unknown | NO3− transporter | Down LN (−1.06) | [60] |
| Lus10032252 | NPF6.3 | Protein NRT1/PTR FAMILY 6.3 | Not in QTL | Unknown | NO3− transporter | Down LN (−1.21) | [58] |
| Lus10004760 | AMT1-2 | Ammonium transporter 1 member 2 | Lu14_9866805 | RV_N− | NH4+ transporter | Down LN (−3.14), Down HN (−2.20) | [61] |
| Lus10008564 | ACO1 | 1-aminocyclopropane-1-carboxylate oxidase 1 | Lu11_14898826 | NUE_N− | Root development | Up LN (1.04) | [62] |
| Lus10036251 | GSO1 | LRR receptor-like serine/threonine-protein kinase GSO1 | Lu12_1386203 | RT_N+, R/S_N− | Root development | Up LN (1.91) | [63] |
| Lus10016554 | FLA4 | Fasciclin-like arabinogalactan protein 4 | Lu12_4334157 | PDW_N+ | Root development | Down LN (−1.02) | [64] |
| Lus10011346 | WRKY75 | Probable WRKY transcription factor 75 | Lu13_19862281 | TRL_Index | Root development | Up LN (1.46) | [65] |
| Lus10042154 | FH8 | Formin-like protein 8 | Lu14_15927781 | RV_N+ | Root development | Up LN (1.53) | [66] |
| Lus10021428 | MYB36 | Transcription factor MYB36 | Lu14_3980668 | R/S_N− | Root development | Down LN (−1.02) | [67] |
| Lus10035126 | RPK2 | LRR receptor-like serine/threonine-protein kinase RPK2 | Lu2_21668160 | PDW_N+ | Root development | Down LN (−1.28) | [63] |
| Lus10024314 | BZIP29 | bZIP transcription factor 29 | Lu6_17695007 | NUE_N+, R/S_N+ | Root development | Down LN (−1.06) | [68] |
| Lus10024908 | NAC021 | NAC domain-containing protein 21/22 | Lu9_19469655 | R/S_N−, RV_Index | Root development | Up LN (1.20) | [69] |
| Lus10031059 | MIZ1 | Protein MIZU-KUSSEI 1 | Lu9_6368499 | NUE_N+ | Root development | Down LN (−1.54), Down HN (−1.40) | [70] |
| Lus10012461 | CEPR2 | Receptor protein-tyrosine kinase CEPR2 | Not in QTL | Unknown | Root development | Up LN (1.08) | [71] |
| Lus10010238 | MYB77 | Transcription factor MYB77 | Not in QTL | Unknown | Root development | Down LN (−1.22) | [72] |
| Lus10040274 | LBD29 | LOB domain-containing protein 29 | Not in QTL | Unknown | Root development | Up LN (1.80) | [73] |
| Lus10025455 | ACR4 | Serine/threonine-protein kinase-like protein ACR4 | Not in QTL | Unknown | Root development | Up LN (1.63) | [74] |
| Lus10040592 | SUNN | Leucine-rich repeat receptor-like kinase protein SUNN | Not in QTL | Unknown | Root development | Down LN (−1.32) | [75] |
| Lus10017271 | ABCB19 | ABC transporter B family member 19 | Not in QTL | Unknown | Root development | Up LN (1.52) | [76] |
| Lus10010012 | ABCB4 | ABC transporter B family member 4 | Not in QTL | Unknown | Root development | Down LN (−1.06) | [77] |
| Lus10005574 | CAT8 | Cationic amino acid transporter 8 | Lu14_17132252 | R/S_N+ | Amino acid transport | Up LN (1.15) | [78] |
| Lus10020181 | GGCT2.1 | Gamma-glutamylcyclotransferase 2.1 | Lu2_4220131 | TRL_Index, RT_Index | Amino acid transport | Up LN (1.38) | [79] |
| Lus10029533 | BAT1 | Amino-acid permease BAT1 homolog | Not in QTL | Unknown | Amino acid transport | Up LN (1.12) | [80] |
| Lus10012824 | AVT1J | Amino acid transporter AVT1J | Not in QTL | Unknown | Amino acid transport | Up LN (1.72) | [81] |
| Lus10008910 | GDU5 | Protein GLUTAMINE DUMPER 5 | Not in QTL | Unknown | Amino acid transport | Down LN (−1.20), Down HN (−1.99) | [82] |
| Lus10042740 | AAP3 | Amino acid permease 3 | Not in QTL | Unknown | Amino acid transport | Down LN (−1.49) | [83] |
| Lus10011451 | BAC2 | Mitochondrial arginine transporter BAC2 | Not in QTL | Unknown | Amino acid transport | Down LN (−1.23) | [84] |
| Lus10015513 | GDU3 | Protein GLUTAMINE DUMPER 3 | Not in QTL | Unknown | Amino acid transport | Down LN (−1.19) | [85] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soto-Cerda, B.J.; Larama, G.; Cloutier, S.; Fofana, B.; Inostroza-Blancheteau, C.; Aravena, G. The Genetic Dissection of Nitrogen Use-Related Traits in Flax (Linum usitatissimum L.) at the Seedling Stage through the Integration of Multi-Locus GWAS, RNA-seq and Genomic Selection. Int. J. Mol. Sci. 2023, 24, 17624. https://doi.org/10.3390/ijms242417624
Soto-Cerda BJ, Larama G, Cloutier S, Fofana B, Inostroza-Blancheteau C, Aravena G. The Genetic Dissection of Nitrogen Use-Related Traits in Flax (Linum usitatissimum L.) at the Seedling Stage through the Integration of Multi-Locus GWAS, RNA-seq and Genomic Selection. International Journal of Molecular Sciences. 2023; 24(24):17624. https://doi.org/10.3390/ijms242417624
Chicago/Turabian StyleSoto-Cerda, Braulio J., Giovanni Larama, Sylvie Cloutier, Bourlaye Fofana, Claudio Inostroza-Blancheteau, and Gabriela Aravena. 2023. "The Genetic Dissection of Nitrogen Use-Related Traits in Flax (Linum usitatissimum L.) at the Seedling Stage through the Integration of Multi-Locus GWAS, RNA-seq and Genomic Selection" International Journal of Molecular Sciences 24, no. 24: 17624. https://doi.org/10.3390/ijms242417624