
Inhibiting Intracellular α2C-Adrenoceptor Surface Translocation Using Decoy Peptides: Identification of an Essential Role of the C-Terminus in Receptor Trafficking


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Transient Co-Transfections
2.2. Domain-Swapping Studies
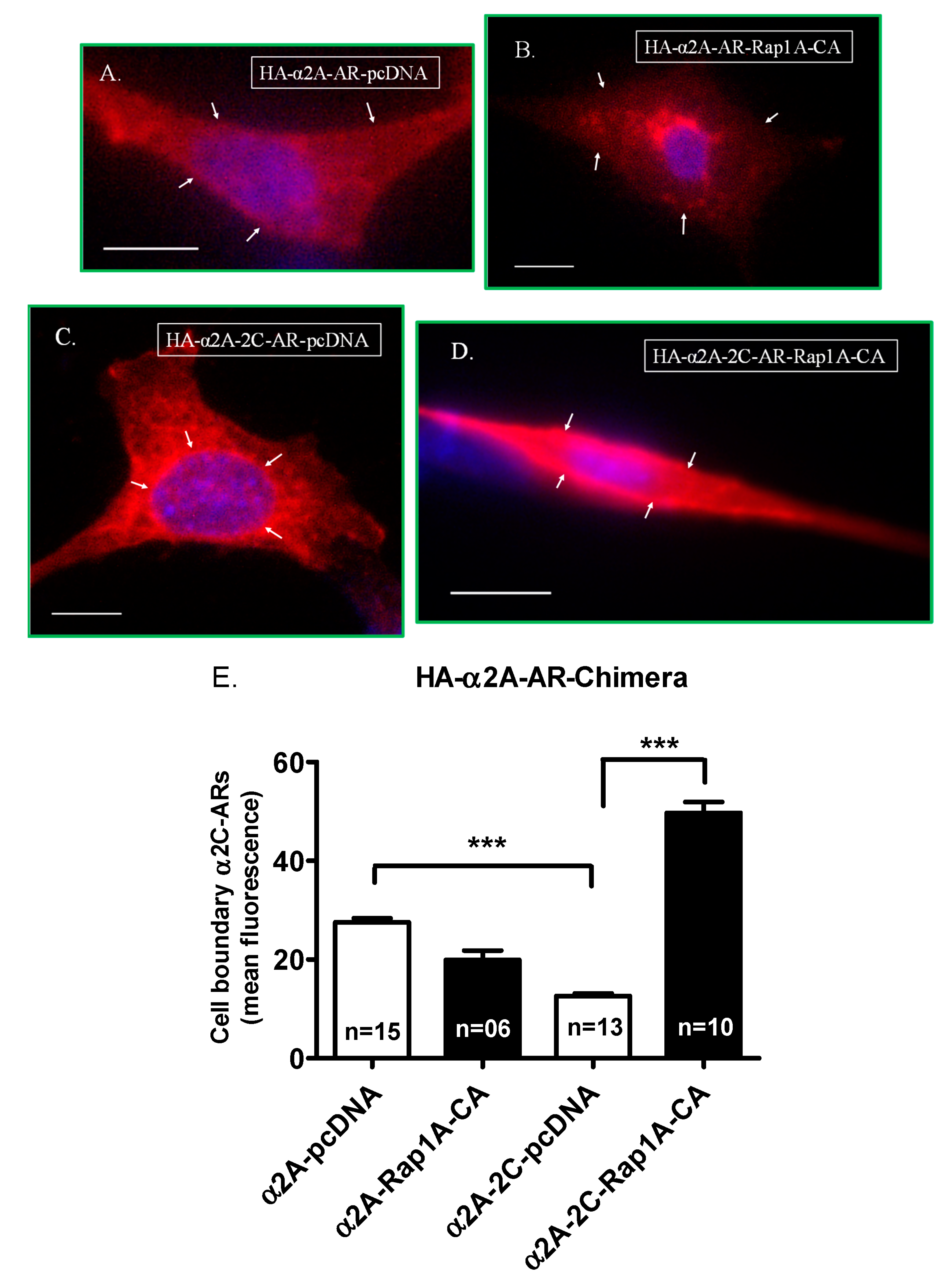
2.2.1. α2A-Adrenoceptor Chimera Studies
2.2.2. α2C-Adrenoceptor Chimera Studies
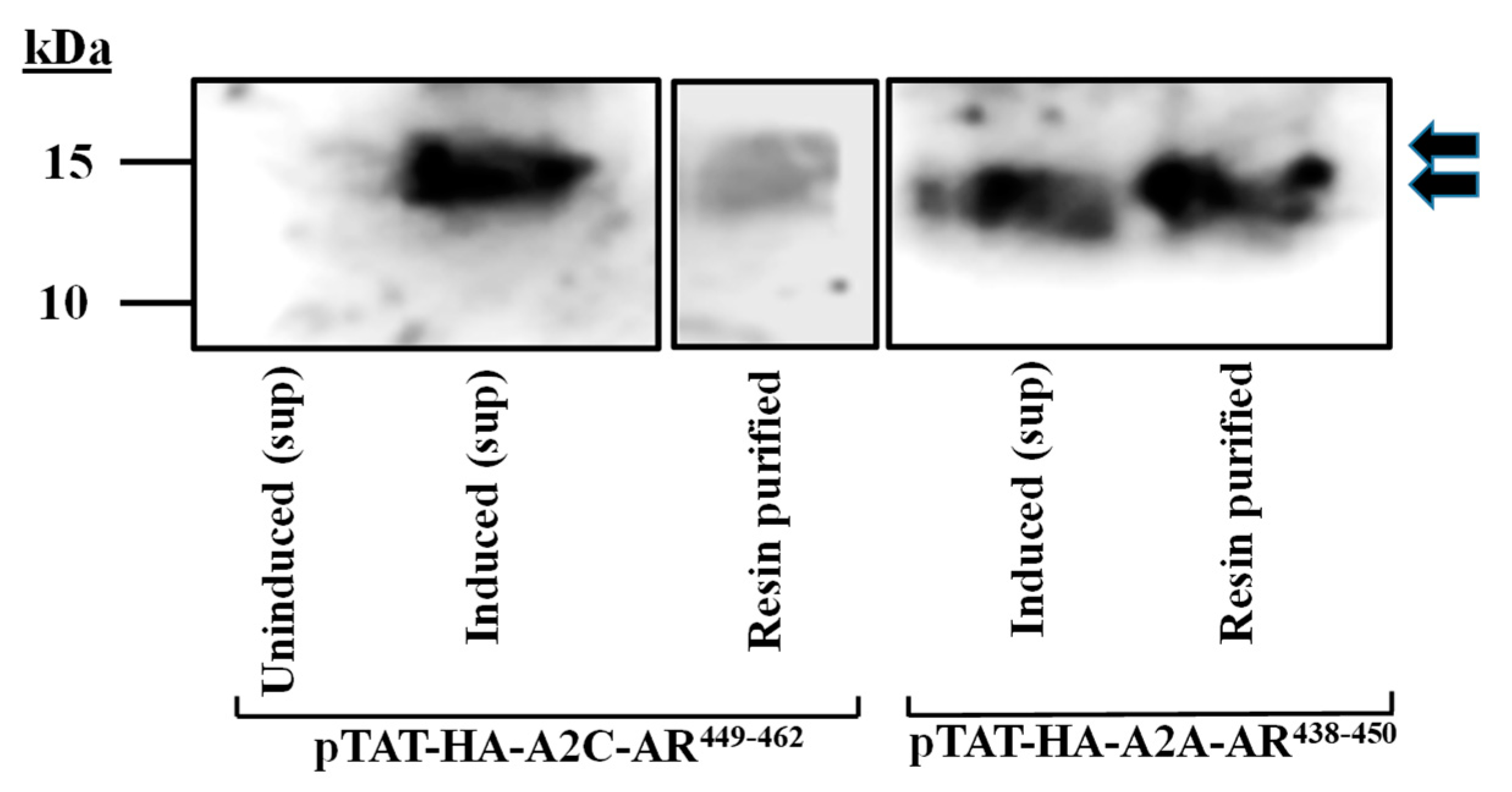
2.3. Peptide Expression, Purification, and Detection by Western Blotting
2.3.1. Peptide Delivery and Localization in NIH/3T3 Cells
2.3.2. Peptide Specificity to Inhibit Receptor Translocation in NIH/3T3 Cells
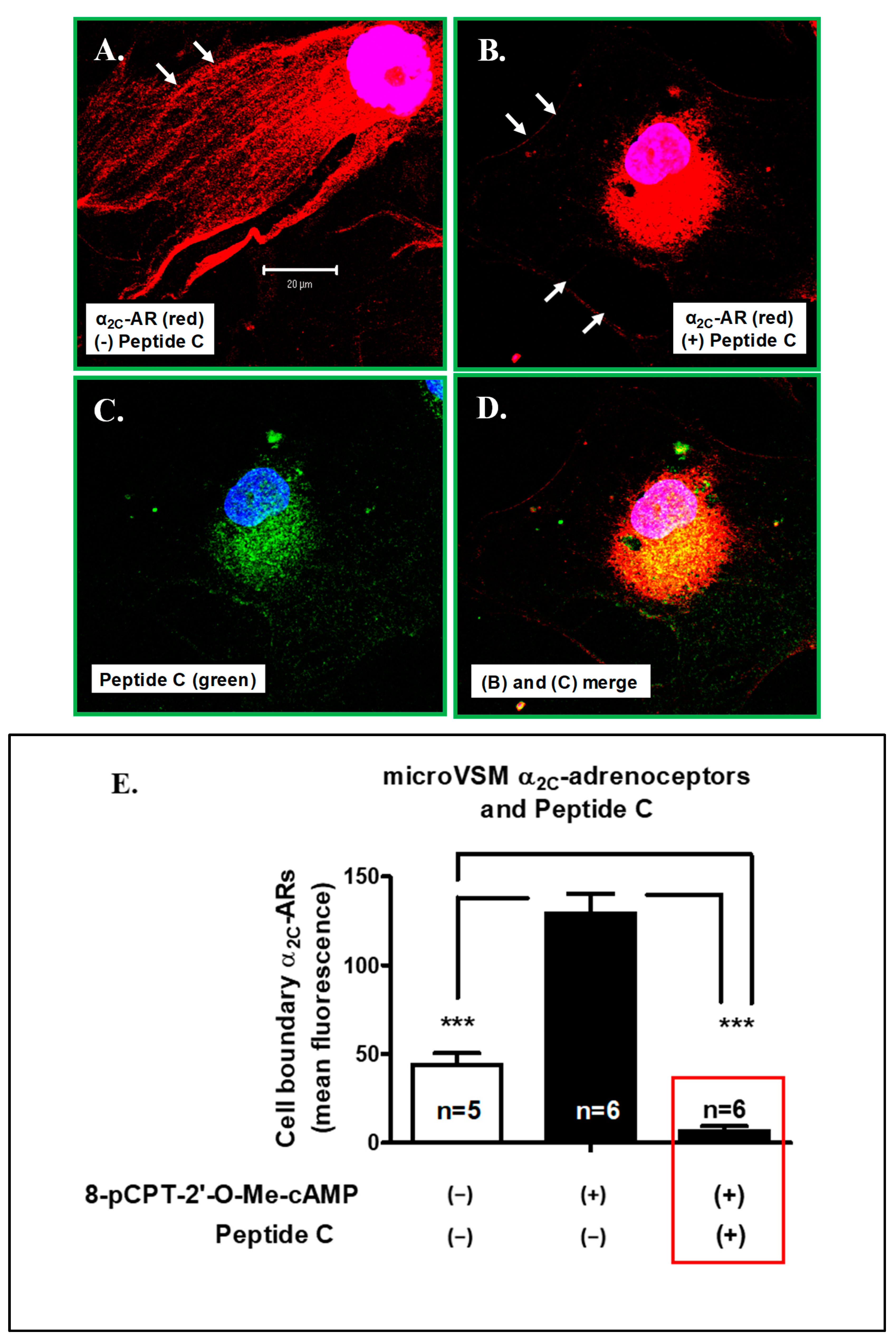
2.3.3. Peptide C Effect on Endogenous microVSM α2C-Adrenoceptor Translocation and Function
3. Discussion
4. Materials and Methods
4.1. Recombinant DNA Used in the Study
4.1.1. Plasmid Constructs
For Domain-Swapping Studies
Hemagglutinin-Tagged Receptors
Green Fluorescent Protein (GFP)-Tagged α2C-Adrenoceptor
Rap1A Expression Construct
For Peptide Expression and Uptake Studies
4.2. Peptide Expression
Peptide Purification with Immobilized Metal Ion Chromatography
4.3. Mammalian Cell Culture
4.3.1. MicroVSM
4.3.2. NIH/3T3
4.4. Transient Transfections and Co-Transfection Studies
Transient Co-Transfections and Peptide Studies
4.5. Immunostaining and Imaging
Imaging and Receptor Quantification
4.6. Assessing the Effect of Peptide C on Receptor Function
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Musa, R.; Qurie, A. Raynaud Disease (Raynaud Phenomenon, Raynaud Syndrome). Available online: https://scholar.google.com.hk/scholar?hl=en&as_sdt=0%2C5&q=Musa%2C+R.%3B+Qurie%2C+A.++Raynaud+Disease+%28Raynaud+Phenomenon%2C+Raynaud+Syndrome%29.+&btnG= (accessed on 25 September 2023).
- Belch, J.J. The Phenomenon, Syndrome and Disease of Maurice Raynaud; Oxford University Press: Oxford, UK, 1990; Volume 29, pp. 162–165. [Google Scholar]
- Hughes, M.; Allanore, Y.; Chung, L.; Pauling, J.D.; Denton, C.P.; Matucci-Cerinic, M. Raynaud phenomenon and digital ulcers in systemic sclerosis. Nat. Rev. Rheumatol. 2020, 16, 208–221. [Google Scholar] [CrossRef] [PubMed]
- Maundrell, A.; Proudman, S.M. Epidemiology of Raynaud’s phenomenon. In Raynaud’s Phenomenon: A Guide to Pathogenesis and Treatment; Springer: Berlin/Heidelberg, Germany, 2014; pp. 21–35. [Google Scholar]
- Pistorius, M.; Pottier, P.; Aymard, B.; Planchon, B. Cold climate could be an etiologic factor involved in Raynaud’s phenomenon physiopathology. Epidemiological investigation from 954 consultations in general practic. Int. Angiol. A J. Int. Union Angiol. 2014, 34, 467–474. [Google Scholar]
- Maricq, H.; Carpentier, P.; Weinrich, M.; Keil, J.; Palesch, Y.; Biro, C.; Vionnet-Fuasset, M.; Jiguet, M.; Valter, I. Geographic variation in the prevalence of Raynaud’s phenomenon: A 5 region comparison. J. Rheumatol. 1997, 24, 879–889. [Google Scholar] [PubMed]
- Block, J.A.; Sequeira, W. Raynaud’s phenomenon. Lancet 2001, 357, 2042–2048. [Google Scholar] [CrossRef] [PubMed]
- Temprano, K.K. A review of Raynaud’s disease. Mo. Med. 2016, 113, 123. [Google Scholar] [PubMed]
- Choi, E.; Henkin, S. Raynaud’s phenomenon and related vasospastic disorders. Vasc. Med. 2021, 26, 56–70. [Google Scholar] [CrossRef]
- Cooke, J.P.; Marshall, J.M. Mechanisms of Raynaud’s disease. Vasc. Med. 2005, 10, 293–307. [Google Scholar] [CrossRef]
- Bunker, C.; Dowd, P.; Terenghi, G.; Springall, D.; Polak, J. Deficiency of calcitonin gene-related peptide in Raynaud’s phenomenon. Lancet 1990, 336, 1530–1533. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Herrick, A.L.; Wigley, F.M. Raynaud’s phenomenon. Best Pract. Res. Clin. Rheumatol. 2020, 34, 101474. [Google Scholar] [CrossRef]
- Nawaz, I.; Nawaz, Y.; Nawaz, E.; Manan, M.R.; Mahmood, A. Raynaud’s phenomenon: Reviewing the pathophysiology and management strategies. Cureus 2022, 14, e21681. [Google Scholar] [CrossRef] [PubMed]
- Flavahan, N.A. Pathophysiological regulation of the cutaneous vascular system in Raynaud’s phenomenon. In Raynaud’s Phenomenon: A Guide to Pathogenesis and Treatment; Springer: Berlin/Heidelberg, Germany, 2014; pp. 57–79. [Google Scholar]
- Langer, S.Z. Presynaptic regulation of catecholamine release. Biochem. Pharmacol. 1974, 23, 1793–1800. [Google Scholar] [CrossRef]
- Link, R.E.; Desai, K.; Hein, L.; Stevens, M.E.; Chruscinski, A.; Bernstein, D.; Barsh, G.S.; Kobilka, B.K. Cardiovascular regulation in mice lacking α2-adrenergic receptor subtypes b and c. Science 1996, 273, 803–805. [Google Scholar] [CrossRef]
- MacMillan, L.B.; Hein, L.; Smith, M.S.; Piascik, M.T.; Limbird, L.E. Central hypotensive effects of the α2A-adrenergic receptor subtype. Science 1996, 273, 801–803. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, E.; Kobilka, B.K.; Scheinin, M. Gene targeting—Homing in on α2-adrenoceptor-subtype function. Trends Pharmacol. Sci. 1997, 18, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Flavahan, N.A. A vascular mechanistic approach to understanding Raynaud Phenomenon. Nat. Rev. Rheumatol. 2015, 11, 146–158. [Google Scholar] [CrossRef]
- Daunt, D.A.; Hurt, C.; Hein, L.; Kallio, J.; Feng, F.; Kobilka, B.K. Subtype-specific intracellular trafficking of α2-adrenergic receptors. Mol. Pharmacol. 1997, 51, 711–720. [Google Scholar] [CrossRef]
- Angelotti, T.; Daunt, D.; Shcherbakova, O.G.; Kobilka, B.; Hurt, C.M. Regulation of G-Protein Coupled Receptor Traffic by an Evolutionary Conserved Hydrophobic Signal. Traffic 2010, 11, 560–578. [Google Scholar] [CrossRef]
- Bailey, S.; Eid, A.; Mitra, S.; Flavahan, S.; Flavahan, N.A. Rho kinase mediates cold-induced constriction of cutaneous arteries: Role of α2C-adrenoceptor translocation. Circ. Res. 2004, 94, 1367–1374. [Google Scholar] [CrossRef]
- Fardoun, M.M.; Nassif, J.; Issa, K.; Baydoun, E.; Eid, A.H. Raynaud’s Phenomenon: A brief review of the underlying mechanisms. Front. Pharmacol. 2016, 7, 438. [Google Scholar] [CrossRef]
- Ma, D.; Zerangue, N.; Lin, Y.-F.; Collins, A.; Yu, M.; Jan, Y.N.; Jan, L.Y. Role of ER export signals in controlling surface potassium channel numbers. Science 2001, 291, 316–319. [Google Scholar] [CrossRef] [PubMed]
- Thompson-Torgerson, C.S.; Holowatz, L.A.; Flavahan, N.A.; Kenney, W.L. Cold-induced cutaneous vasoconstriction is mediated by Rho kinase in vivo in human skin. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1700–H1705. [Google Scholar] [CrossRef]
- Chotani, M.A.; Mitra, S.; Su, B.Y.; Flavahan, S.; Eid, A.H.; Clark, K.R.; Montague, C.R.; Paris, H.; Handy, D.E.; Flavahan, N.A. Regulation of α2-adrenoceptors in human vascular smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H59–H67. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.H.; Chotani, M.A.; Mitra, S.; Miller, T.J.; Flavahan, N.A. Cyclic AMP acts through Rap1 and JNK signaling to increase expression of cutaneous smooth muscle α2C-adrenoceptors. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H266–H272. [Google Scholar] [CrossRef] [PubMed]
- Jeyaraj, S.C.; Unger, N.T.; Eid, A.H.; Mitra, S.; Paul El-Dahdah, N.; Quilliam, L.A.; Flavahan, N.A.; Chotani, M.A. Cyclic AMP-Rap1A signaling activates RhoA to induce α2c-adrenoceptor translocation to the cell surface of microvascular smooth muscle cells. Am. J. Physiol. Cell Physiol. 2012, 303, C499–C511. [Google Scholar] [CrossRef] [PubMed]
- Vadlamudi, R.K.; Li, F.; Adam, L.; Nguyen, D.; Ohta, Y.; Stossel, T.P.; Kumar, R. Filamin is essential in actin cytoskeletal assembly mediated by p21-activated kinase 1. Nat. Cell Biol. 2002, 4, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.S.; Ohta, Y.; Rabinovitz, I.; Stossel, T.P.; Blenis, J. Ribosomal S6 kinase (RSK) regulates phosphorylation of filamin A on an important regulatory site. Mol. Cell. Biol. 2004, 24, 3025–3035. [Google Scholar] [CrossRef]
- Shahwar, D.; Saeed, M.; Hazrat, H.; Javaid, S.; Mohsin, S.; Ul-Haq, Z.; Chotani, M.A. The nucleoside adenosine inhibits intracellular microvascular α2C-adrenoceptor surface trafficking. J. Mol. Struct. 2022, 1267, 133637. [Google Scholar] [CrossRef]
- Motawea, H.K.; Jeyaraj, S.C.; Eid, A.H.; Mitra, S.; Unger, N.T.; Ahmed, A.A.; Flavahan, N.A.; Chotani, M.A. Cyclic AMP-Rap1A signaling mediates cell surface translocation of microvascular smooth muscle α2C-adrenoceptors through the actin-binding protein filamin-2. Am. J. Physiol. Cell Physiol. 2013, 305, C829–C845. [Google Scholar] [CrossRef]
- Gong, J.; Liu, J.; Ronan, E.A.; He, F.; Cai, W.; Fatima, M.; Zhang, W.; Lee, H.; Li, Z.; Kim, G.-H. A cold-sensing receptor encoded by a glutamate receptor gene. Cell 2019, 178, 1375–1386.e1311. [Google Scholar] [CrossRef]
- Chen, X.; Xu, Y.; Qu, L.; Wu, L.; Han, G.W.; Guo, Y.; Wu, Y.; Zhou, Q.; Sun, Q.; Chu, C. Molecular mechanism for ligand recognition and subtype selectivity of α2C adrenergic receptor. Cell Rep. 2019, 29, 2936–2943.e2934. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.A.; Wigley, F.M.; White, B.; Leatherman, G.; Zhong, J.; Krasa, H.; Kambayashi, J.i.; Orlandi, C.; Czerwiec, F.S. Efficacy and tolerability of a selective α2C-adrenergic receptor blocker in recovery from cold-induced vasospasm in scleroderma patients: A single-center, double-blind, placebo-controlled, randomized crossover study. Arthritis Rheum. 2004, 50, 3994–4001. [Google Scholar] [CrossRef] [PubMed]
- Kapsa, S.; Thuss, U.; Boxnick, S.; Schaumann, F.; Schultz, A.; Unger, S.; Otto, C. Pharmacokinetics, Safety, and Tolerability of the α2C-Adrenoreceptor Antagonist BAY 1193397 in Healthy Male Subjects. Clin. Pharmacol. Drug Dev. 2022, 11, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Chotani, M.A.; Flavahan, N.A. Intracellular α2C-adrenoceptors: Storage depot, stunted development or signaling domain? Biochim. Et Biophys. Acta (BBA) Mol. Cell Res. 2011, 1813, 1495–1503. [Google Scholar] [CrossRef] [PubMed]
- Hummers, L.K.; Wigley, F.M. Management of Raynaud’s phenomenon and digital ischemic lesions in scleroderma. Rheum. Dis. Clin. 2003, 29, 293–313. [Google Scholar] [CrossRef]
- Boin, F.; Wigley, F.M. Understanding, assessing and treating Raynaud’s phenomenon. Curr. Opin. Rheumatol. 2005, 17, 752–760. [Google Scholar] [PubMed]
- Henness, S.; Wigley, F.M. Current drug therapy for scleroderma and secondary Raynaud’s phenomenon: Evidence-based review. Curr. Opin. Rheumatol. 2007, 19, 611–618. [Google Scholar] [CrossRef]
- Riemekasten, G.; Sunderkötter, C. Vasoactive therapies in systemic sclerosis. Rheumatology 2006, 45, iii49–iii51. [Google Scholar] [CrossRef]
- Pawlowski, M.; Saraswathi, S.; Motawea, H.K.; Chotani, M.A.; Kloczkowski, A. In silico modeling of human α2C-adrenoreceptor interaction with filamin-2. PLoS ONE 2014, 9, e103099. [Google Scholar] [CrossRef]
- Philippe, G.J.; Craik, D.J.; Henriques, S.T. Converting peptides into drugs targeting intracellular protein–protein interactions. Drug Discov. Today 2021, 26, 1521–1531. [Google Scholar] [CrossRef]
- Chotani, M.A.; Mitra, S.; Eid, A.H.; Han, S.A.; Flavahan, N.A. Distinct cAMP signaling pathways differentially regulate α2C-adrenoceptor expression: Role in serum induction in human arteriolar smooth muscle cells. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H69–H76. [Google Scholar] [CrossRef] [PubMed]
- Rosengren, A.H.; Jokubka, R.; Tojjar, D.; Granhall, C.; Hansson, O.; Li, D.Q.; Nagaraj, V.; Reinbothe, T.M.; Tuncel, J.; Eliasson, L.; et al. Overexpression of alpha2A-adrenergic receptors contributes to type 2 diabetes. Science 2010, 327, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Tervi, A.R.M.; Abner, E.; Cheng, P.; Lane, J.M.; Maher, M.; Lammi, V.; Strausz, S.; Nguyen, T.; Docampo, M.L.; Gu, W.; et al. The α2A-adrenergic receptor (ADRA2A) modulates 3 susceptibility to Raynaud’s syndrome. Medrxiv 2023. [Google Scholar] [CrossRef]
- Hartmann, S.; Yasmeen, S.; Jacobs, B.M.; Denaxas, S.; Pirmohamed, M.; Gamazon, E.R.; Caulfield, M.J.; Genes & Health Research Team; Hemingway, H.; Pietzner, M.; et al. ADRA2A and IRX1 are putative risk genes for Raynaud’s phenomenon. Nat. Commun. 2023, 14, 6156. [Google Scholar] [CrossRef]
- Salvi, E.; Kutalik, Z.; Glorioso, N.; Benaglio, P.; Frau, F.; Kuznetsova, T.; Arima, H.; Hoggart, C.; Tichet, J.; Nikitin, Y.P.; et al. Genomewide association study using a high-density single nucleotide polymorphism array and case-control design identifies a novel essential hypertension susceptibility locus in the promoter region of endothelial NO synthase. Hypertension 2012, 59, 248–255. [Google Scholar] [CrossRef]
- Sauzeau, V.; Le Jeune, H.; Cario-Toumaniantz, C.; Smolenski, A.; Lohmann, S.M.; Bertoglio, J.; Chardin, P.; Pacaud, P.; Loirand, G. Cyclic GMP-dependent protein kinase signaling pathway inhibits RhoA-induced Ca2+ sensitization of contraction in vascular smooth muscle. J. Biol. Chem. 2000, 275, 21722–21729. [Google Scholar] [CrossRef]
- Sauzeau, V.; Sevilla, M.A.; Montero, M.J.; Bustelo, X.R. The Rho/Rac exchange factor Vav2 controls nitric oxide-dependent responses in mouse vascular smooth muscle cells. J. Clin. Investig. 2010, 120, 315–330. [Google Scholar] [CrossRef]
- Freedman, R.R.; Girgis, R.; Mayes, M.D. Acute effect of nitric oxide on Raynaud’s phenomenon in scleroderma. Lancet 1999, 354, 739. [Google Scholar] [CrossRef]
- Rembold, C.M.; Ayers, C.R. Oral L-arginine can reverse digital necrosis in Raynaud’s phenomenon. Mol. Cell. Biochem. 2003, 244, 139–141. [Google Scholar] [CrossRef]
- Levien, T.L. Phosphodiesterase inhibitors in Raynaud’s phenomenon. Ann. Pharmacother. 2006, 40, 1388–1393. [Google Scholar] [CrossRef]
- Matucci Cerinic, M.; Kahaleh, M.B. Beauty and the beast. The nitric oxide paradox in systemic sclerosis. Rheumatology 2002, 41, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Kamata, Y.; Kamimura, T.; Iwamoto, M.; Minota, S. Comparable effects of sildenafil citrate and alprostadil on severe Raynaud’s phenomenon in a patient with systemic sclerosis. Clin. Exp. Dermatol. 2005, 30, 451. [Google Scholar] [CrossRef] [PubMed]
- Fries, R.; Shariat, K.; von Wilmowsky, H.; Bohm, M. Sildenafil in the treatment of Raynaud’s phenomenon resistant to vasodilatory therapy. Circulation 2005, 112, 2980–2985. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, H.; Vocero-Akbani, A.M.; Snyder, E.L.; Ho, A.; Latham, D.G.; Lissy, N.A.; Becker-Hapak, M.; Ezhevsky, S.A.; Dowdy, S.F. Transduction of full-length TAT fusion proteins into mammalian cells: TAT-p27Kip1 induces cell migration. Nat. Med. 1998, 4, 1449–1452. [Google Scholar] [CrossRef]
- Wadia, J.S.; Dowdy, S.F. Transmembrane delivery of protein and peptide drugs by TAT-mediated transduction in the treatment of cancer. Adv. Drug Deliv. Rev. 2005, 57, 579–596. [Google Scholar] [CrossRef]
- Gump, J.M.; Dowdy, S.F. TAT transduction: The molecular mechanism and therapeutic prospects. Trends Mol. Med. 2007, 13, 443–448. [Google Scholar] [CrossRef]

















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raza, A.; Mohsin, S.; Saeed, F.; Ali, S.A.; Chotani, M.A. Inhibiting Intracellular α2C-Adrenoceptor Surface Translocation Using Decoy Peptides: Identification of an Essential Role of the C-Terminus in Receptor Trafficking. Int. J. Mol. Sci. 2023, 24, 17558. https://doi.org/10.3390/ijms242417558
Raza A, Mohsin S, Saeed F, Ali SA, Chotani MA. Inhibiting Intracellular α2C-Adrenoceptor Surface Translocation Using Decoy Peptides: Identification of an Essential Role of the C-Terminus in Receptor Trafficking. International Journal of Molecular Sciences. 2023; 24(24):17558. https://doi.org/10.3390/ijms242417558
Chicago/Turabian StyleRaza, Aisha, Saima Mohsin, Fasiha Saeed, Syed Abid Ali, and Maqsood A. Chotani. 2023. "Inhibiting Intracellular α2C-Adrenoceptor Surface Translocation Using Decoy Peptides: Identification of an Essential Role of the C-Terminus in Receptor Trafficking" International Journal of Molecular Sciences 24, no. 24: 17558. https://doi.org/10.3390/ijms242417558