
Neurological Improvement via Lysophosphatidic Acid Administration in a Rodent Model of Cardiac Arrest-Induced Brain Injury
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Baseline Characteristics
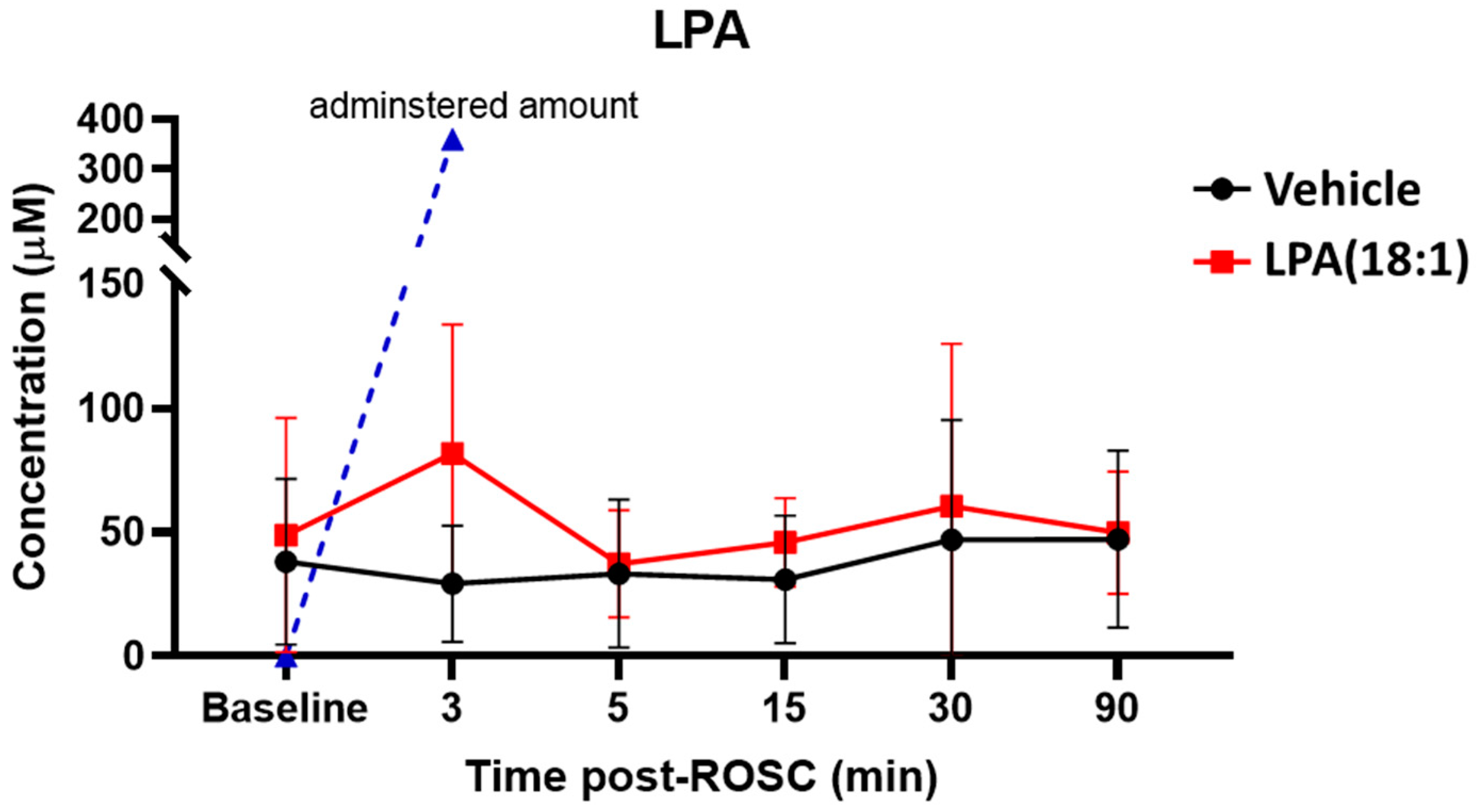
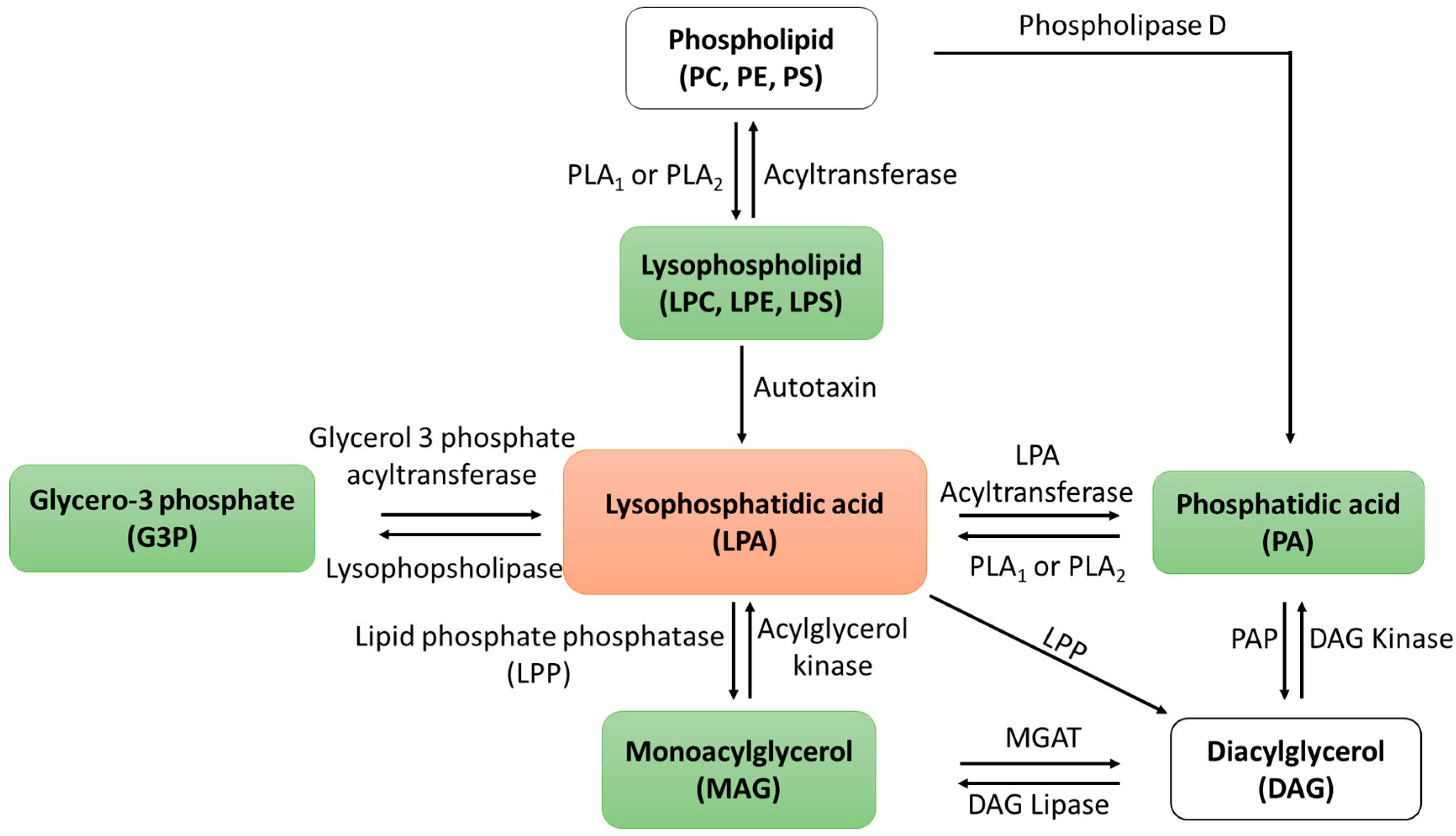
2.2. Changes in LPA Levels Post-CA
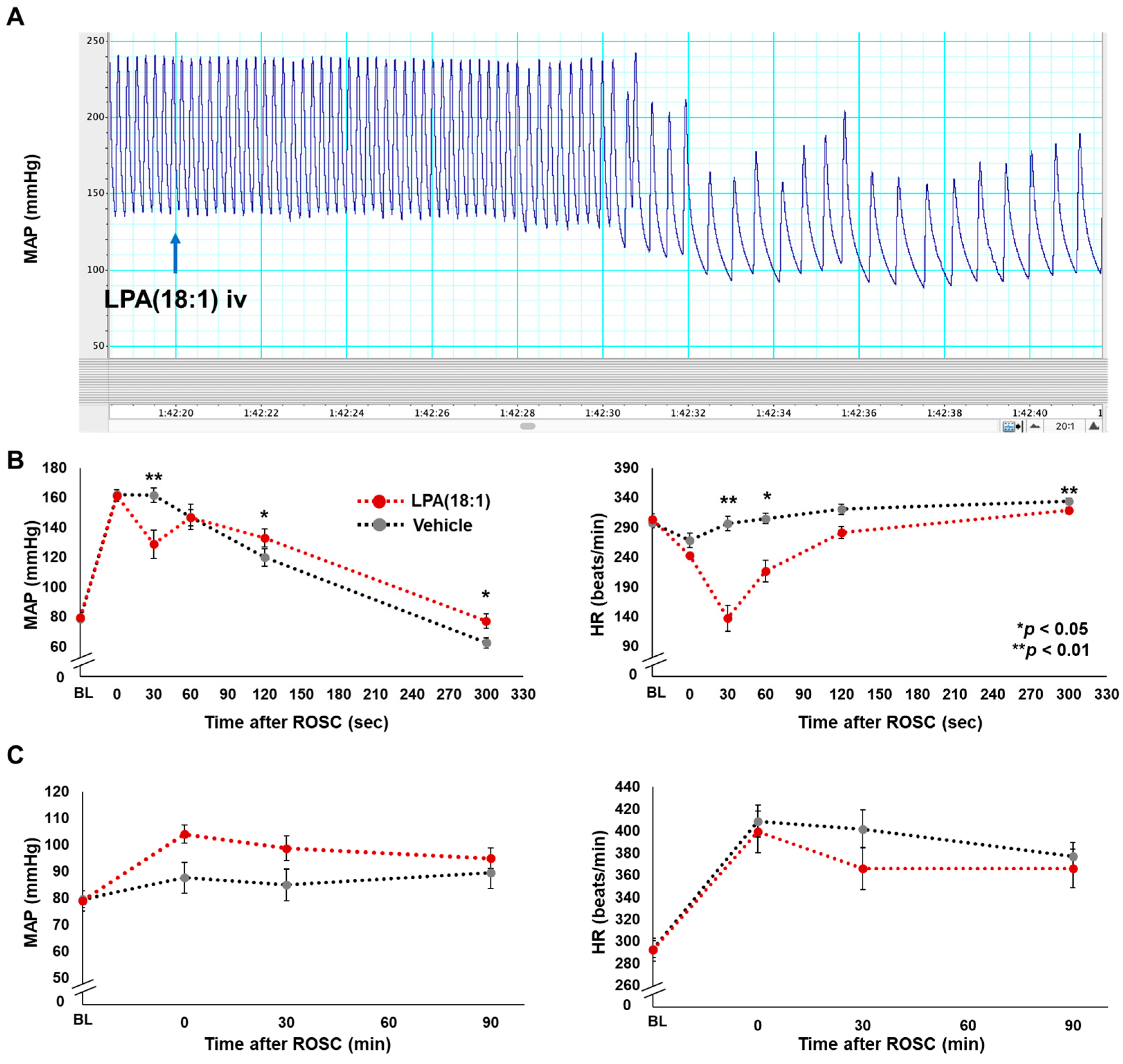
2.3. Effects of LPA (18:1) on Hemodynamics Post-CA
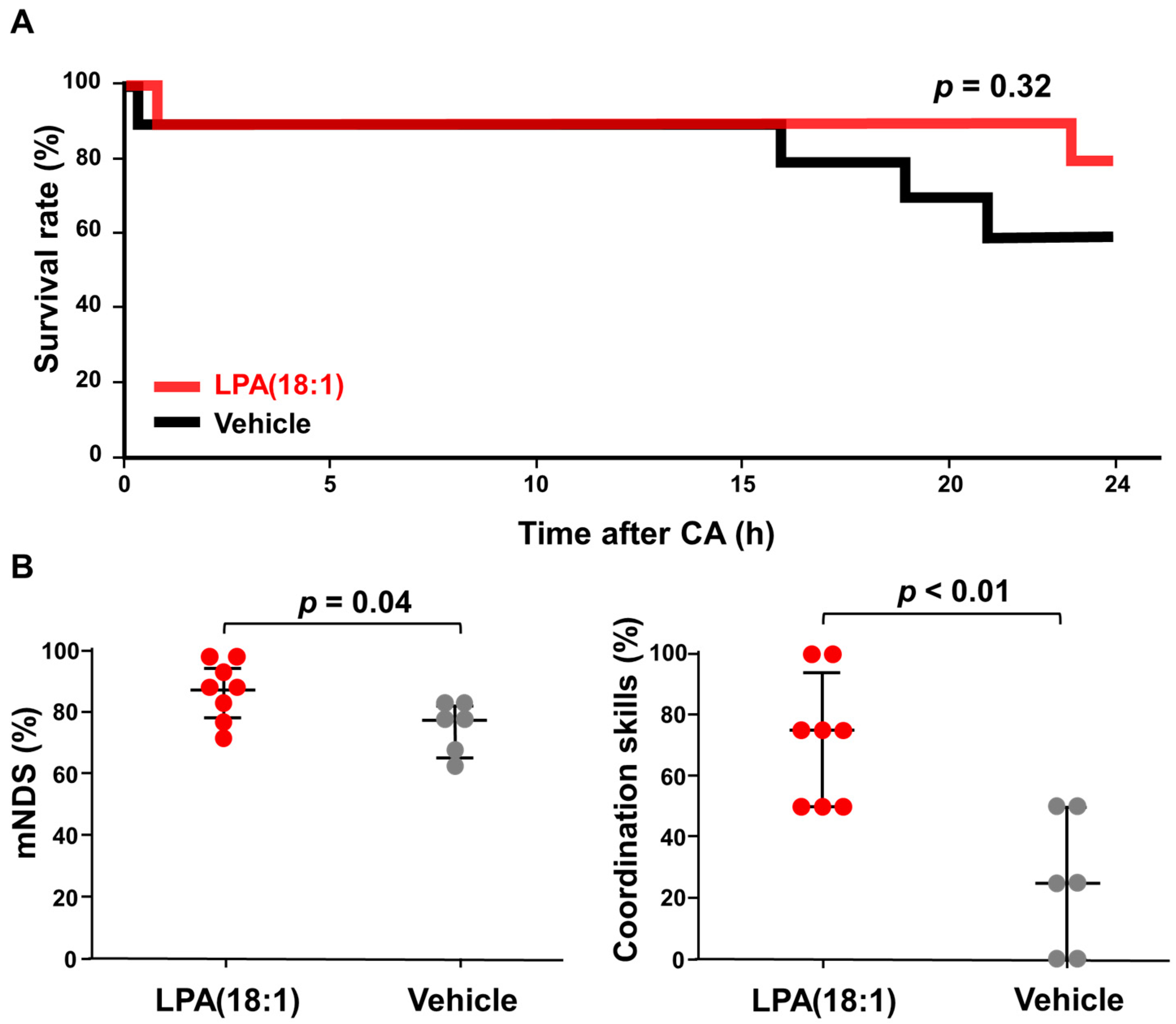
2.4. Effects of LPA (18:1) on Brain Function Post-CA
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
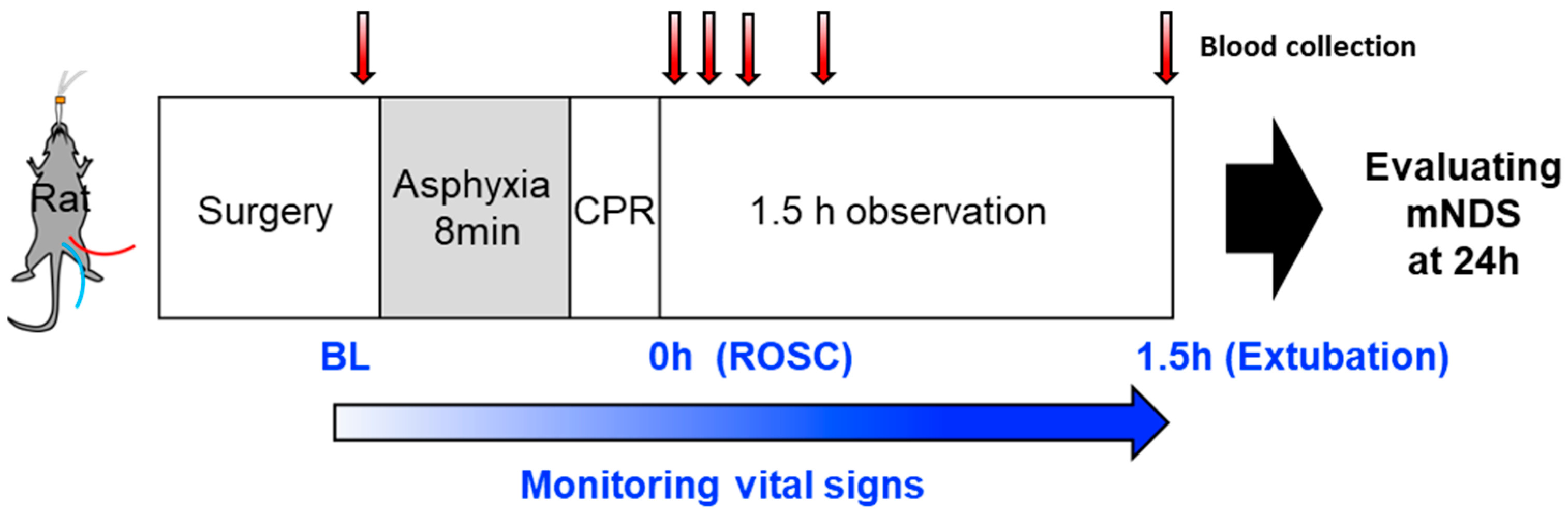
4.2. Animal Preparation
4.3. LPA Administration
4.4. Hemodynamics and Survival Outcomes
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Thel, M.C.; Armstrong, A.L.; McNulty, S.E.; Califf, R.M.; O’Connor, C.M. Randomised trial of magnesium in in-hospital cardiac arrest. Duke Internal Medicine Housestaff. Lancet 1997, 350, 1272–1276. [Google Scholar] [CrossRef] [PubMed]
- Argaud, L.; Cour, M.; Dubien, P.Y.; Giraud, F.; Jossan, C.; Riche, B.; Hernu, R.; Darmon, M.; Poncelin, Y.; Tchenio, X.; et al. Effect of Cyclosporine in Nonshockable Out-of-Hospital Cardiac Arrest: The CYRUS Randomized Clinical Trial. JAMA Cardiol. 2016, 1, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Naito, H.; Nojima, T.; Fujisaki, N.; Tsukahara, K.; Yamamoto, H.; Yamada, T.; Aokage, T.; Yumoto, T.; Osako, T.; Nakao, A. Therapeutic strategies for ischemia reperfusion injury in emergency medicine. Acute Med. Surg. 2020, 7, e501. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.-Q.; Fang, X.-J.; Zhang, Y.; Qiao, J.-T. Neuroprotective effect of lysophosphatidic acid on AbetaP31-35-induced apoptosis in cultured cortical neurons. Sheng Li Xue Bao 2005, 57, 289–294. [Google Scholar] [PubMed]
- Fan, H.; Zingarelli, B.; Harris, V.; Tempel, G.E.; Halushka, P.V.; Cook, J.A. Lysophosphatidic acid inhibits bacterial endotoxin-induced pro-inflammatory response: Potential anti-inflammatory signaling pathways. Mol. Med. 2008, 14, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Crack, P.J.; Zhang, M.; Morganti-Kossmann, M.C.; Morris, A.J.; Wojciak, J.M.; Fleming, J.K.; Karve, I.; Wright, D.; Sashindranath, M.; Goldshmit, Y.; et al. Anti-lysophosphatidic acid antibodies improve traumatic brain injury outcomes. J. Neuroinflamm. 2014, 11, 37. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-G.; Yu, Z.-C.; Yu, Y.-P.; Ju, W.-P.; Wang, D.-Z.; Zhan, X.; Wu, X.-J.; Zhou, L. Lysophosphatidic Acid Level and the Incidence of Silent Brain Infarction in Patients with Nonvalvular Atrial Fibrillation. Int. J. Mol. Sci. 2010, 11, 3988–3998. [Google Scholar] [CrossRef]
- Zeng, X.; Luo, Z.; Wu, J.; Zhou, J.; Shan, Y.; Zhu, Y.; Yan, G.; Lin, Y.; Wang, C. Cerebral Ischemia-Reperfusion Injury: Lysophosphatidic Acid Mediates Inflammation by Decreasing the Expression of Liver X Receptor. J. Mol. Neurosci. 2020, 70, 1376–1384. [Google Scholar] [CrossRef]
- Sarker, M.H.; Hu, D.-E.; Fraser, P.A. Regulation of Cerebromicrovascular Permeability by Lysophosphatidic Acid. Microcirculation 2010, 17, 39–46. [Google Scholar] [CrossRef]
- Schulze, C.; Smales, C.; Rubin, L.L.; Staddon, J.M. Lysophosphatidic Acid Increases Tight Junction Permeability in Cultured Brain Endothelial Cells. J. Neurochem. 1997, 68, 991–1000. [Google Scholar] [CrossRef]
- On, N.H.; Savant, S.; Toews, M.; Miller, D.W. Rapid and Reversible Enhancement of Blood–Brain Barrier Permeability Using Lysophosphatidic Acid. J. Cereb. Blood Flow Metab. 2013, 33, 1944–1954. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, J.; Tang, J.; Li, Y.-Y.; Gu, Y.; Yu, Y.; Xiong, J.; Zhao, X.; Zhang, Z.; Li, T.-T.; et al. Lysophosphatidic acid induces neuronal cell death via activation of asparagine endopeptidase in cerebral ischemia-reperfusion injury. Exp. Neurol. 2018, 306, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chi, O.Z.; Mellender, S.J.; Kiss, G.K.; Chiricolo, A.; Liu, X.; Patel, N.; Jacinto, E.; Weiss, H.R. Lysophosphatidic acid increased infarct size in the early stage of cerebral ischemia-reperfusion with increased BBB permeability. J. Stroke Cerebrovasc. Dis. 2020, 29, 105029. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, C.; Caramujo, M.J. The Various Roles of Fatty Acids. Molecules 2018, 23, 2583. [Google Scholar] [CrossRef] [PubMed]
- Glick, N.R.; Fischer, M.H. The Role of Essential Fatty Acids in Human Health. J. Evidence-Based Complement. Altern. Med. 2013, 18, 268–289. [Google Scholar] [CrossRef]
- Song, J.; Kim, Y.-S.; Lee, D.H.; Lee, S.H.; Park, H.J.; Lee, D.; Kim, H. Neuroprotective effects of oleic acid in rodent models of cerebral ischaemia. Sci. Rep. 2019, 9, 10732. [Google Scholar] [CrossRef]
- Sakurai, K.; Shen, C.; Shiraishi, I.; Inamura, N.; Hisatsune, T. Consumption of Oleic Acid on the Preservation of Cognitive Functions in Japanese Elderly Individuals. Nutrients 2021, 13, 284. [Google Scholar] [CrossRef]
- Cury-Boaventura, M.F.; Gorjão, R.; de Lima, T.M.; Newsholme, P.; Curi, R. Comparative toxicity of oleic and linoleic acid on human lymphocytes. Life Sci. 2006, 78, 1448–1456. [Google Scholar] [CrossRef]
- Choi, J.; Yin, T.; Shinozaki, K.; Lampe, J.W.; Stevens, J.F.; Becker, L.B.; Kim, J. Comprehensive analysis of phospholipids in the brain, heart, kidney, and liver: Brain phospholipids are least enriched with polyunsaturated fatty acids. Mol. Cell. Biochem. 2018, 442, 187–201. [Google Scholar] [CrossRef]
- Katz, L.; Ebmeyer, U.; Safar, P.; Radovsky, A.; Neumar, R. Outcome model of asphyxial cardiac arrest in rats. J. Cereb. Blood Flow Metab. 1995, 15, 1032–1039. [Google Scholar] [CrossRef]
- Jendzjowsky, N.G.; Roy, A.; Barioni, N.O.; Kelly, M.M.; Green FH, Y.; Wyatt, C.N.; Pye, R.L.; Tenorio-Lopes, L.; Wilson RJ, A. Preventing acute asthmatic symptoms by targeting a neuronal mechanism involving carotid body lysophos-phatidic acid receptors. Nat. Commun. 2018, 9, 4030. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Hasse, S.; Zhao, C.; Bourgoin, S.G. Targeting the autotaxin—Lysophosphatidic acid receptor axis in car-diovascular diseases. Biochem. Pharmacol. 2019, 164, 74–81. [Google Scholar] [CrossRef]
- Sullivan, E.M.; Pennington, E.R.; Sparagna, G.C.; Torres, M.J.; Neufer, P.D.; Harris, M.; Washington, J.; Anderson, E.J.; Zeczycki, T.N.; Brown, D.A.; et al. Docosahexaenoic acid lowers cardiac mitochondrial enzyme activity by replacing linoleic acid in the phospholipidome. J. Biol. Chem. 2018, 293, 466–483. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, C.M.; Le, C.H.; deMooy, A.B.; Nelson, C.B.; Chicco, A.J. Inhibition of delta-6 desaturase reverses car-diolipin remodeling and prevents contractile dysfunction in the aged mouse heart without altering mitochondrial respiratory function. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harslof, L.B.; Ciappolino, V.; Agostoni, C. DHA Effects in Brain Devel-opment and Function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Sugasini, D.; Thomas, R.; Yalagala PC, R.; Tai, L.M.; Subbaiah, P.V. Dietary docosahexaenoic acid (DHA) as lyso-phosphatidylcholine, but not as free acid, enriches brain DHA and improves memory in adult mice. Sci. Rep. 2017, 7, 11263. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Yin, T.; Shinozaki, K.; Lampe, J.W.; Becker, L.B. DHA-supplemented diet increases the survival of rats fol-lowing asphyxia-induced cardiac arrest and cardiopulmonary bypass resuscitation. Sci. Rep. 2016, 6, 36545. [Google Scholar] [CrossRef]
- Nishikimi, M.; Yagi, T.; Shoaib, M.B.; Takegawa, R.; Rasul, R.M.; Hayashida, K.; Okuma, Y.; Yin, T.; Choudhary, R.C.; Becker, L.B.M.; et al. Phospholipid Screening Postcardiac Arrest Detects Decreased Plasma Lysophosphatidylcholine: Supplementation as a New Therapeutic Approach. Crit. Care Med. 2022, 50, e199–e208. [Google Scholar] [CrossRef]
- Shinozaki, K.; Becker, L.B.; Saeki, K.; Kim, J.; Yin, T.; Da, T.; Lampe, J.W. Dissociated Oxygen Consumption and Carbon Dioxide Production in the Post–Cardiac Arrest Rat: A Novel Metabolic Phenotype. J. Am. Hear. Assoc. 2018, 7, e007721. [Google Scholar] [CrossRef]
- Neumar, R.W.; Bircher, N.G.; Sim, K.M.; Xiao, F.; Zadach, K.S.; Radovsky, A.; Katz, L.; Ebmeyer, E.; Safar, P. Ep-inephrine and sodium bicarbonate during CPR following asphyxial cardiac arrest in rats. Resuscitation 1995, 29, 249–263. [Google Scholar] [CrossRef]
- Han, F.; Boller, M.; Guo, W.; Merchant, R.M.; Lampe, J.W.; Smith, T.M.; Becker, L.B. A rodent model of emergency cardiopulmonary bypass resuscitation with different temperatures after asphyxial cardiac arrest. Resuscitation 2010, 81, 93–99. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Vehicle iv (n = 10) | 18:1 LPA iv (n = 10) | p-Value |
|---|---|---|---|
| Body weight, g | 436 ± 9.9 | 427 ± 8.4 | 0.79 |
| Surgical procedure time, min | 33.7 ± 1.9 | 32.6 ± 1.5 | 0.97 |
| Before CA | |||
| MAP, mmHg | 76.6 ± 2.9 | 79.6 ± 3.4 | 0.85 |
| HR, bpm | 298 ± 7.3 | 293 ± 9.5 | 0.85 |
| RR, times/min | 45.8 ± 0.1 | 45.9 ± 0.1 | 0.54 |
| BT, °C | 36.3 ± 0.2 | 36.3 ± 0.1 | 0.76 |
| Until CA time, s | 111 ± 13.6 | 118 ± 11.7 | 0.82 |
| CPR time, s | 70 ± 3.8 | 69 ± 3.2 | 0.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishikimi, M.; Choudhary, R.C.; Shoaib, M.; Yagi, T.; Becker, L.B.; Kim, J. Neurological Improvement via Lysophosphatidic Acid Administration in a Rodent Model of Cardiac Arrest-Induced Brain Injury. Int. J. Mol. Sci. 2023, 24, 17451. https://doi.org/10.3390/ijms242417451
Nishikimi M, Choudhary RC, Shoaib M, Yagi T, Becker LB, Kim J. Neurological Improvement via Lysophosphatidic Acid Administration in a Rodent Model of Cardiac Arrest-Induced Brain Injury. International Journal of Molecular Sciences. 2023; 24(24):17451. https://doi.org/10.3390/ijms242417451
Chicago/Turabian StyleNishikimi, Mitsuaki, Rishabh C. Choudhary, Muhammad Shoaib, Tsukasa Yagi, Lance B. Becker, and Junhwan Kim. 2023. "Neurological Improvement via Lysophosphatidic Acid Administration in a Rodent Model of Cardiac Arrest-Induced Brain Injury" International Journal of Molecular Sciences 24, no. 24: 17451. https://doi.org/10.3390/ijms242417451