
Genome-Wide Identification and Expression Analysis of the SWEET Gene Family in Capsicum annuum L.
 , ,
, ,
Abstract
:1. Introduction
2. Results
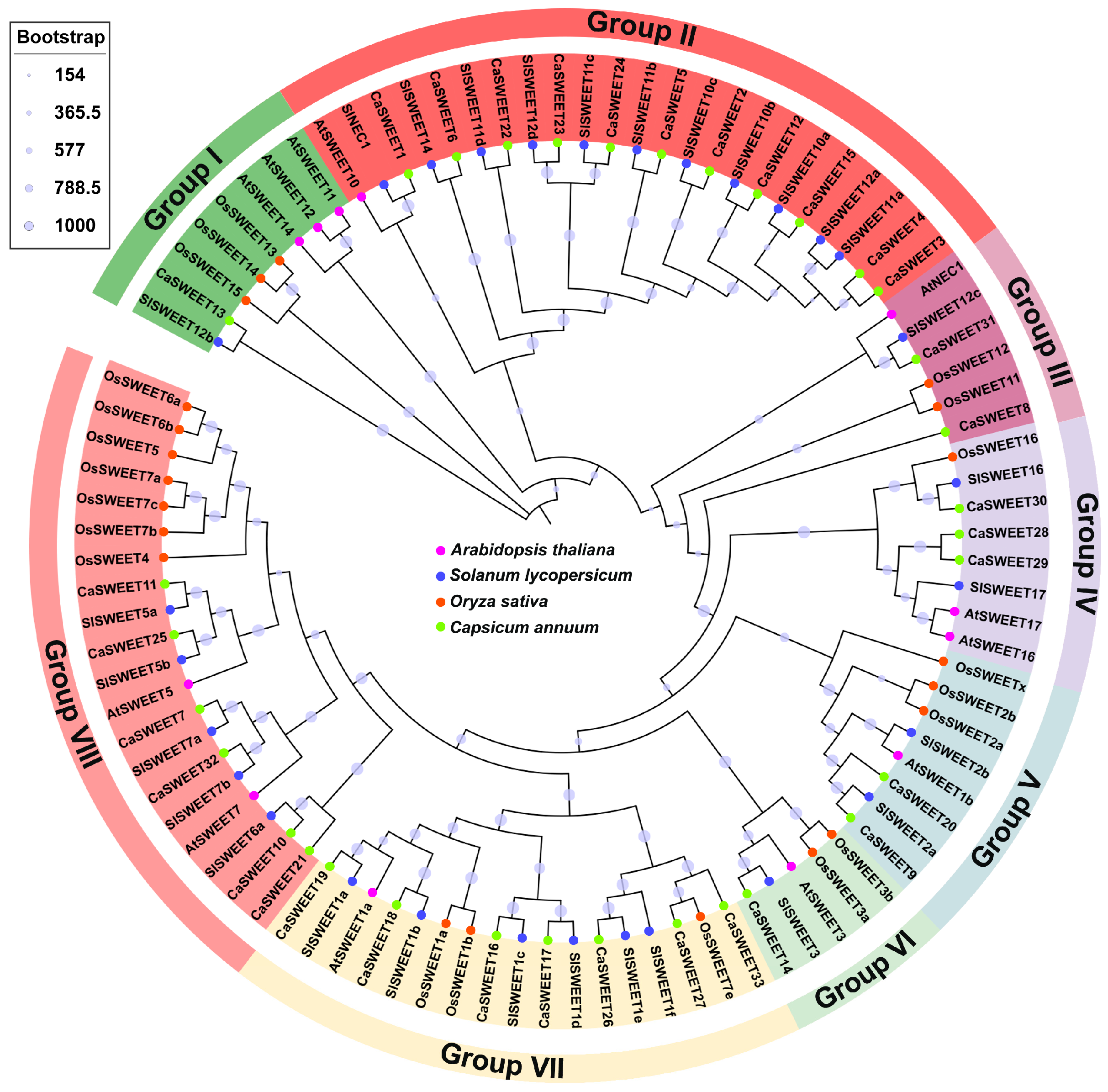
2.1. Identification and Phylogenetic Analysis of CaSWEETs
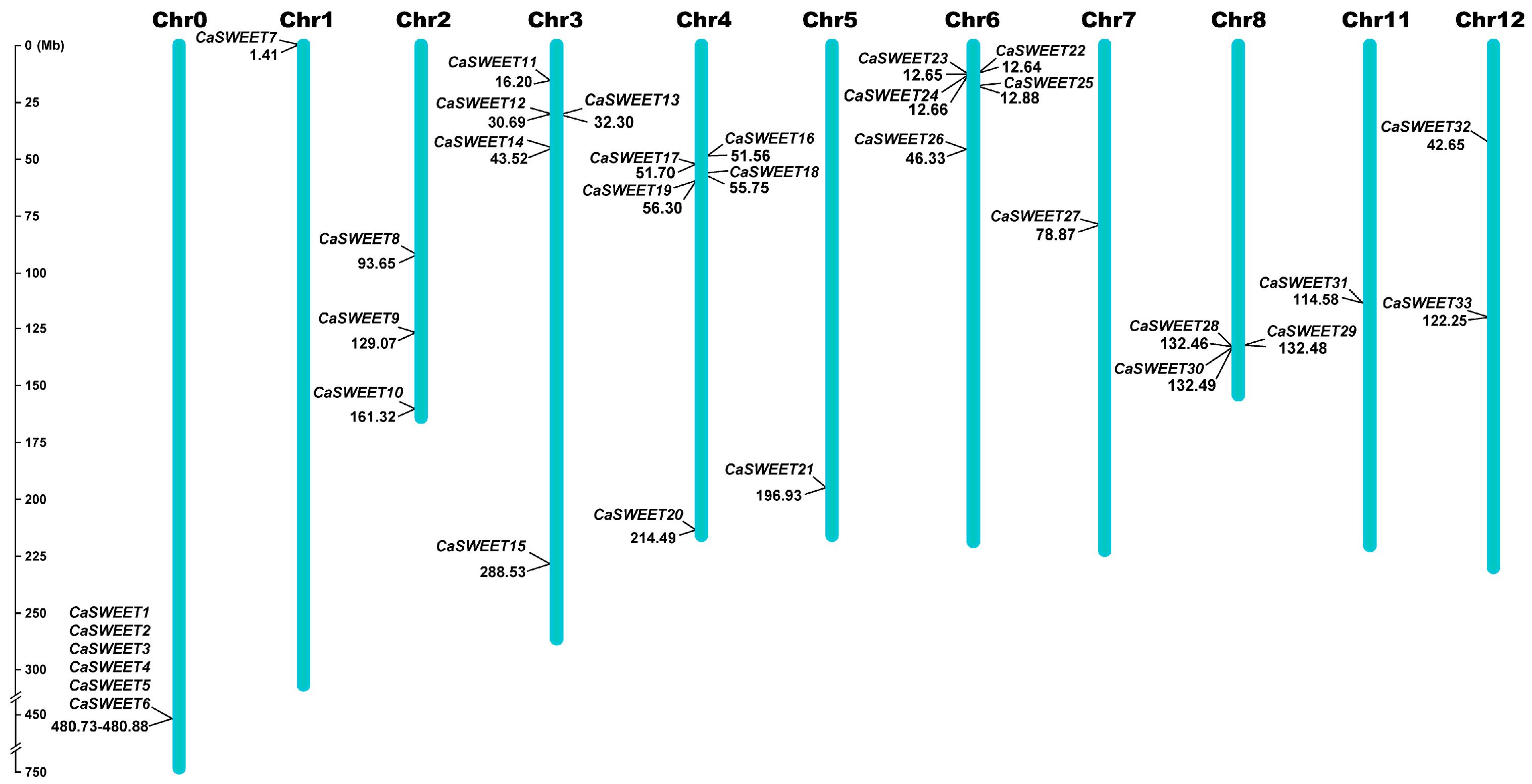
2.2. CaSWEETs Chromosomal Distribution
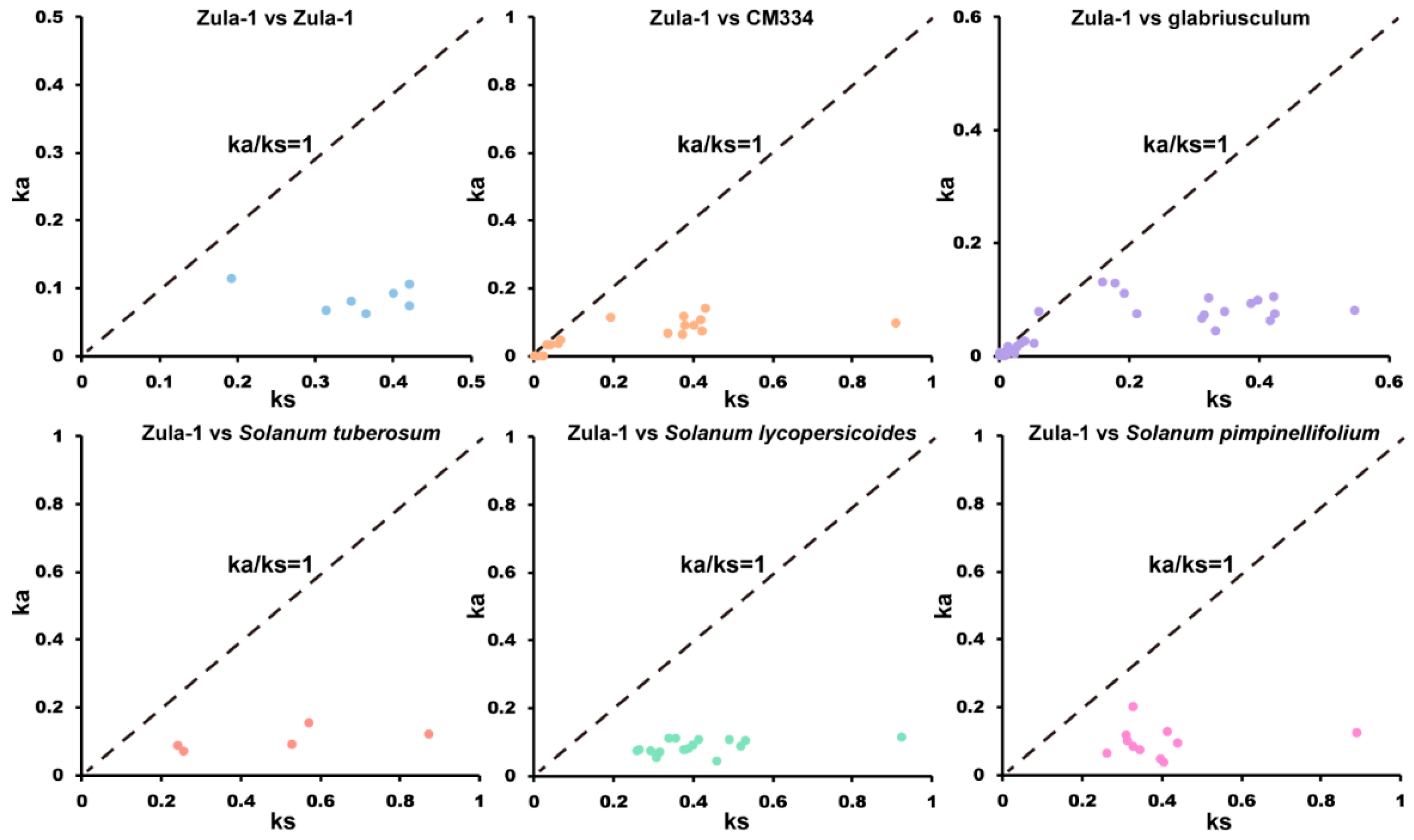
2.3. SWEET Family Members Exhibit Evolutionary Conservation
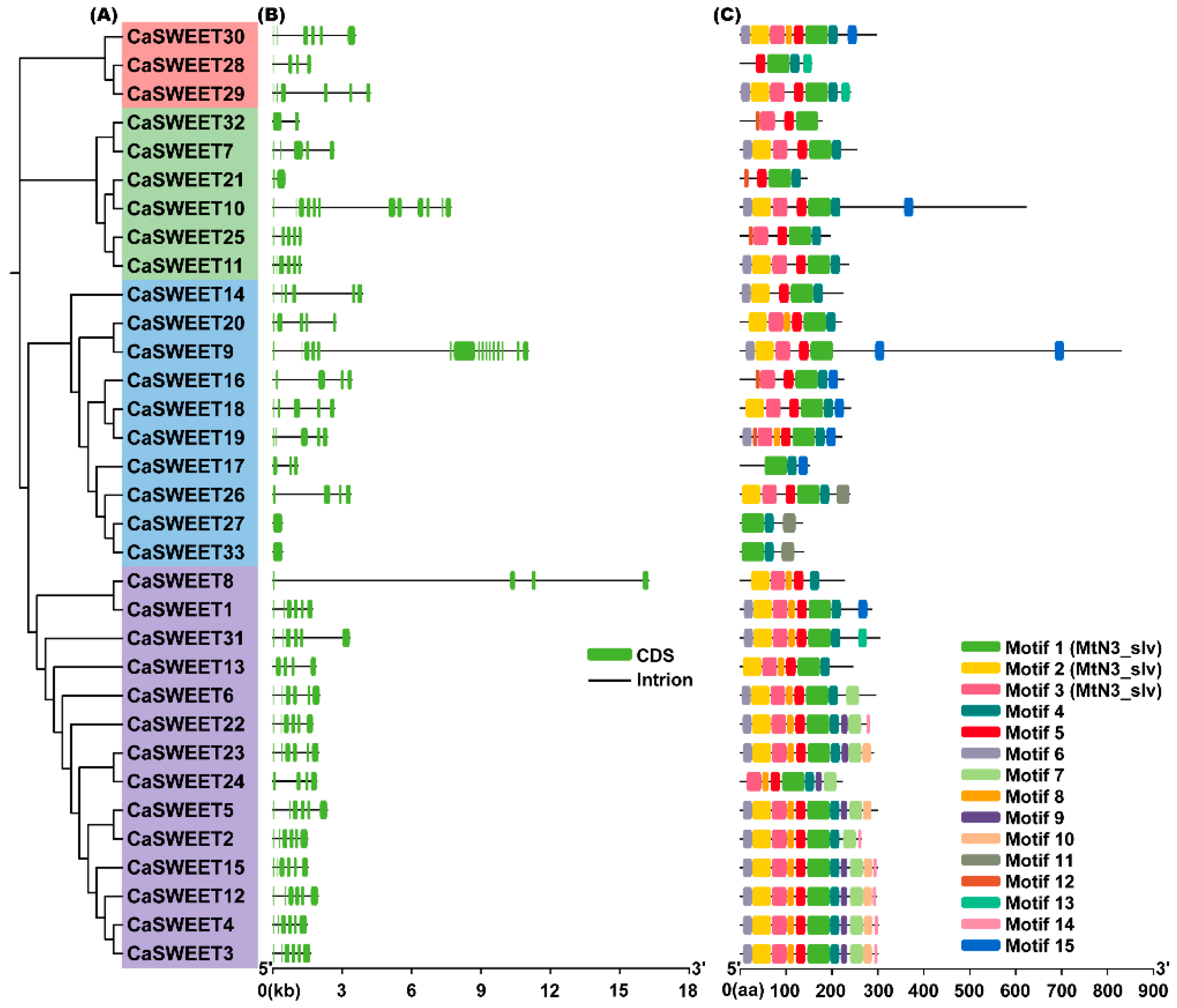
2.4. Gene Structure and Conserved Motif Analysis of CaSWEETs
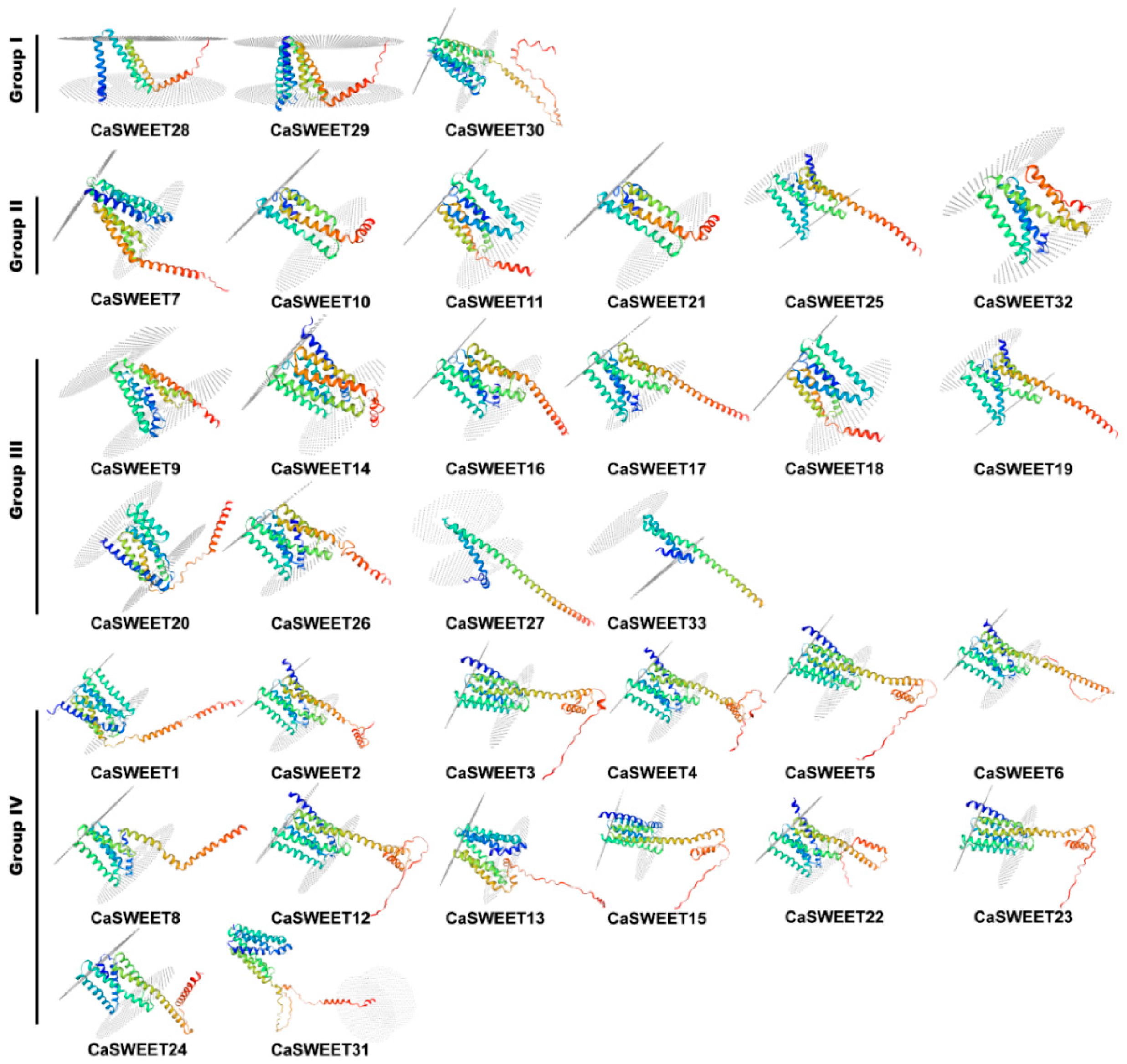
2.5. CaSWEET Protein Characterization and Three-Dimensional Structure Analysis
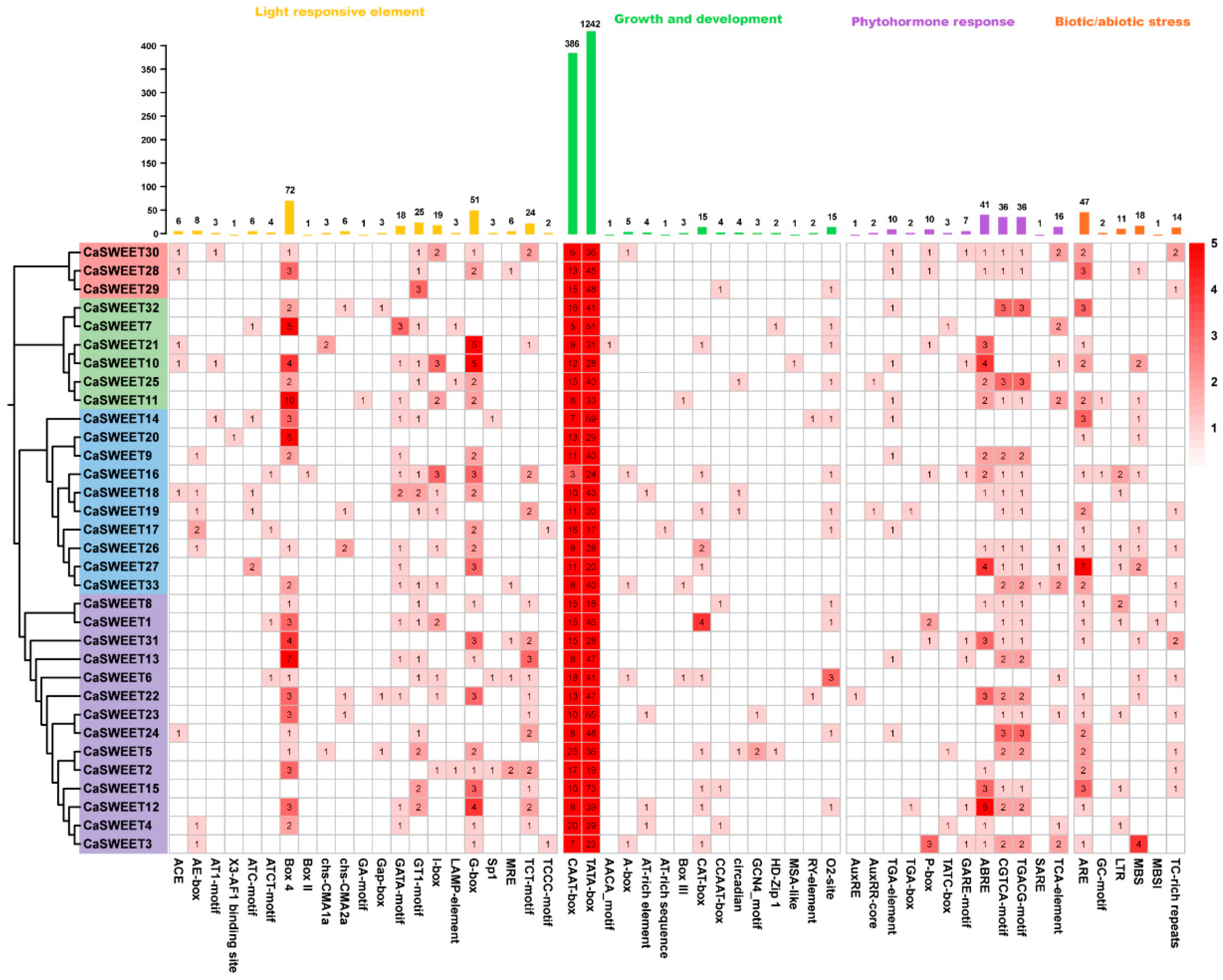
2.6. CaSWEETs Contain Versatility Cis-Regulatory Elements
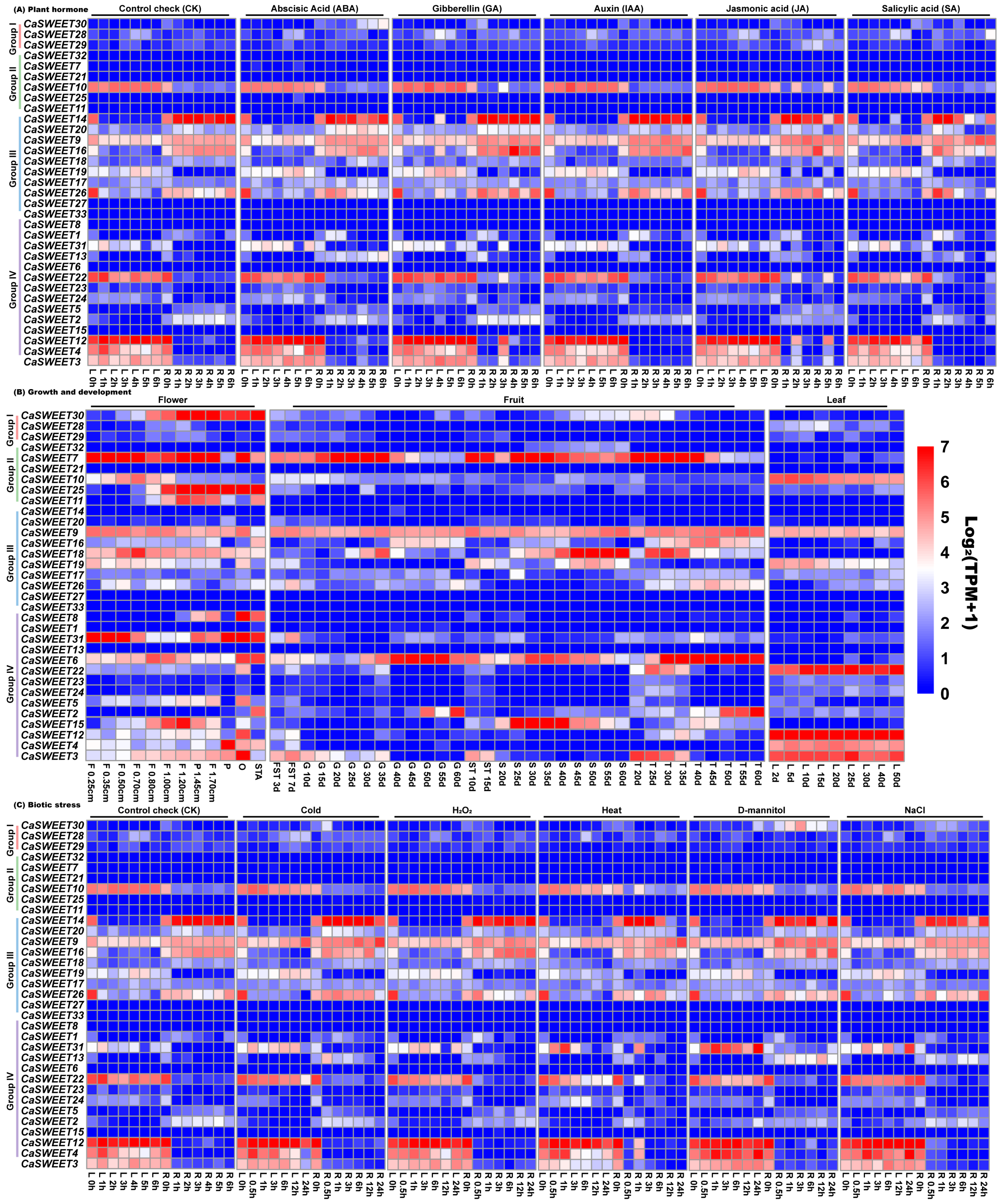
2.7. Transcriptomic Analysis Revealed Multiple CaSWEET Expression Patterns
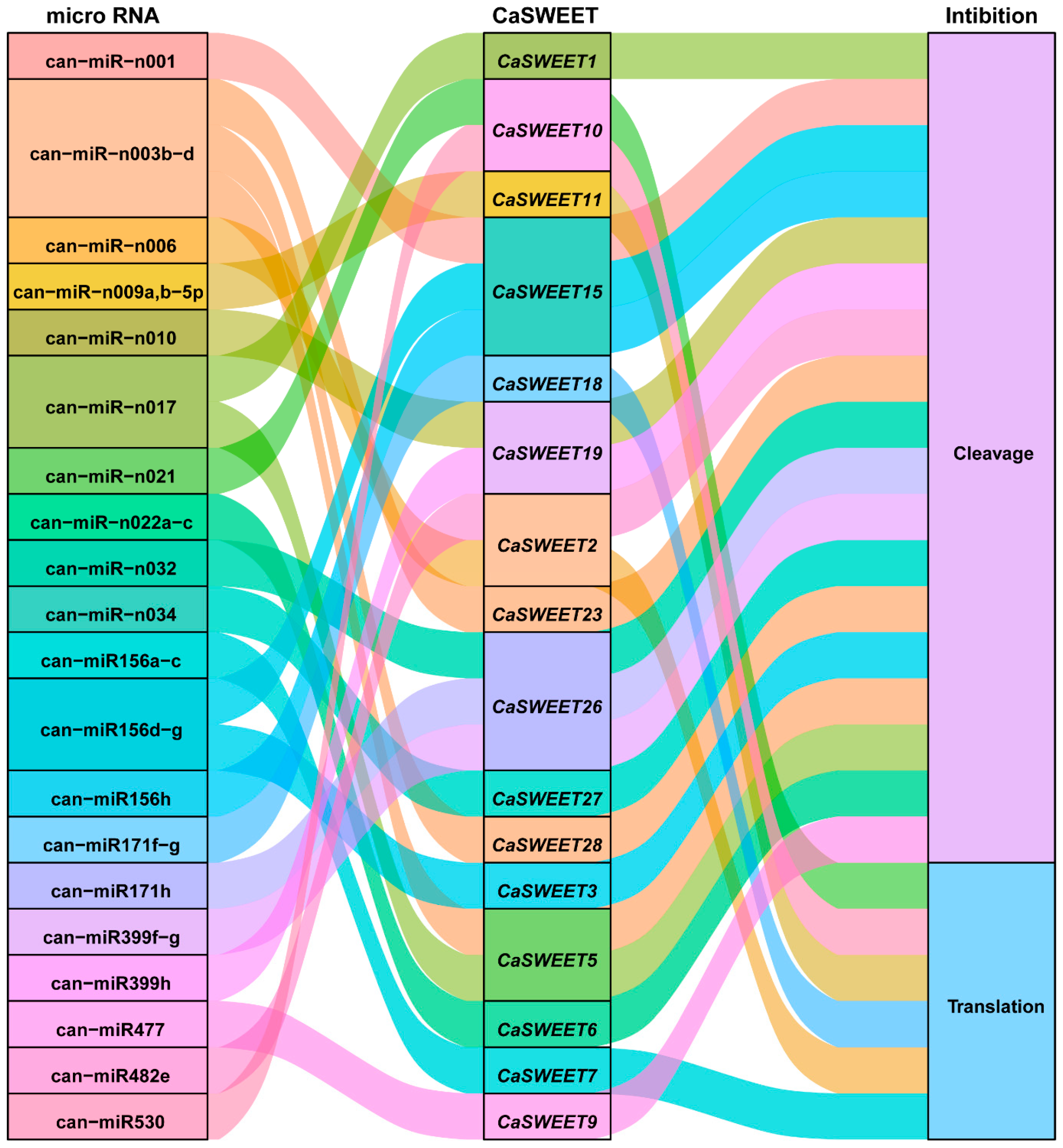
2.8. MicroRNA (miRNA) Regulates The Post-Transcriptional Expression of CaSWEETs
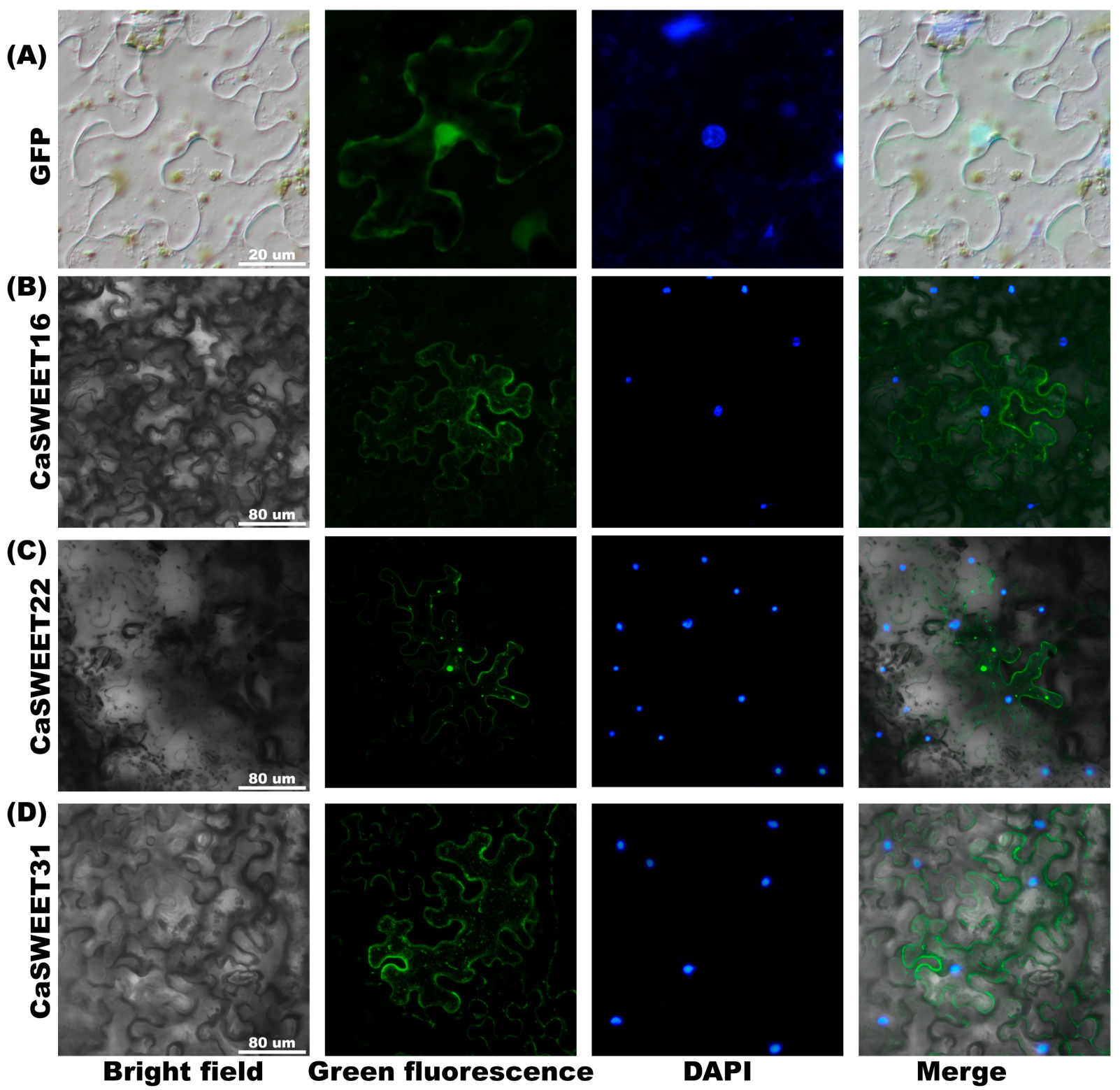
2.9. Subcellular Localization Analysis of CaSWEET16/22/31
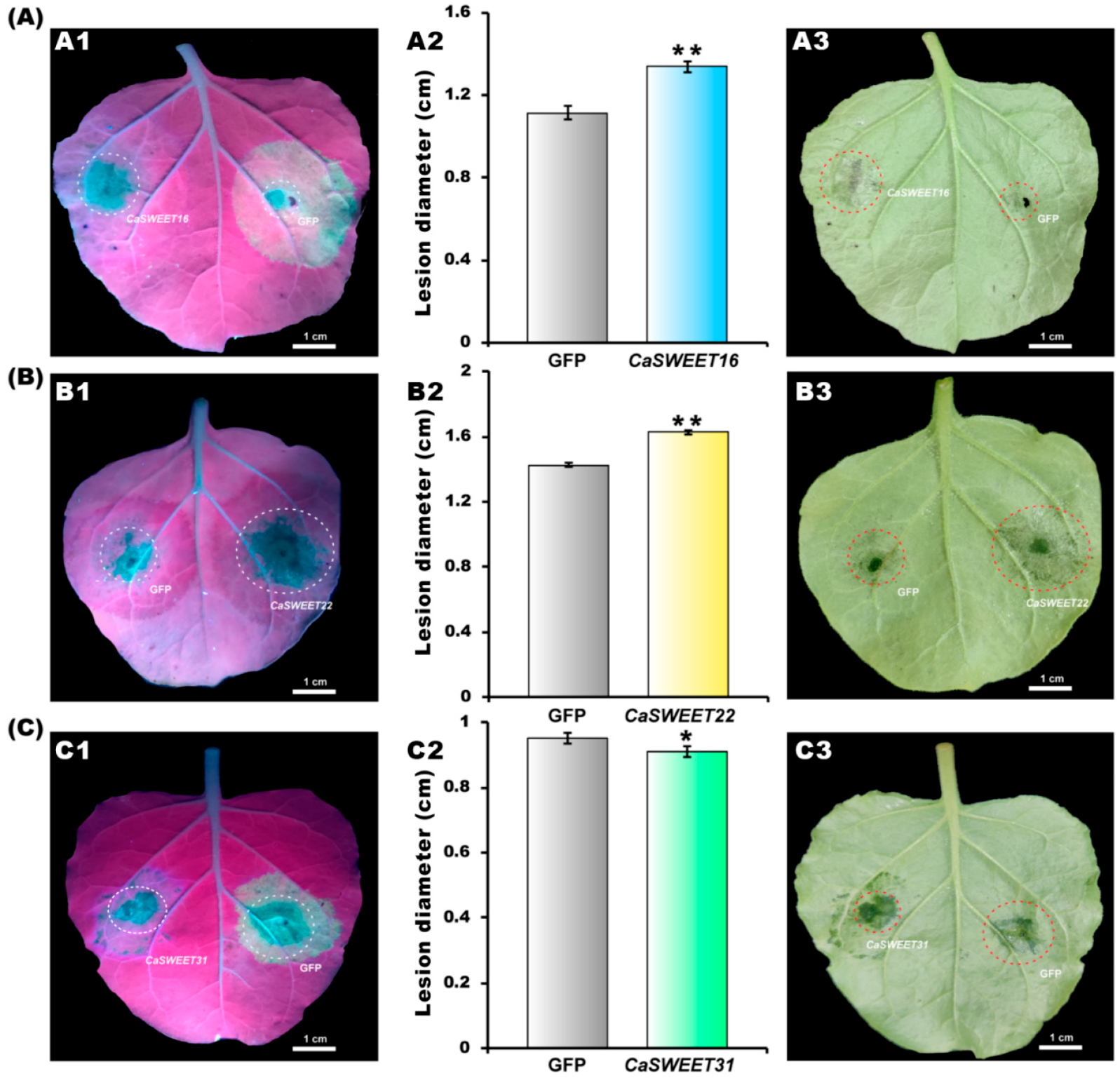
2.10. CaSWEET16/22 Promote and CaSWEET31 Represses Pathogen Colonization
3. Discussion
4. Materials and Methods
4.1. Identification of the SWEET Gene Family Members in Pepper
4.2. Phylogenetic Analysis of SWEET
4.3. Chromosomal Localization of CaSWEETs
4.4. Interspecific Evolutionary Analysis of SWEETs
4.5. CaSWEETS Gene Structure and Conserved Motif Analysis
4.6. Protein Characterization and Three-Dimensional Homology Modeling of CaSWEETs
4.7. CaSWEET Promoter Cis-Acting Elements Analysis
4.8. CaSWEET Expression Pattern Analysis
4.9. Prediction of miRNA Targeting Relationships with CaSWEETs
4.10. Subcellular Localization Analysis of CaSWEET16/22/31
4.11. Pathogen Inoculation Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Arora, H.; Sharma, A.; Sharma, S.; Haron, F.F.; Gafur, A.; Sayyed, R.Z.; Datta, R. Pythium damping-off and root rot of Capsicum annuum L.: Impacts, diagnosis, and management. Microorganisms 2021, 9, 823. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q. Analysis of Production Cost and Income of Pepper in Main Production Areas in China—Based on the Investigation of Five Provinces Such as Guizhou and Hunan; Agricultural University of Hebei: Baoding, China, 2018. [Google Scholar]
- Guo, M.; Liu, J.H.; Lu, J.P.; Zhai, Y.F.; Wang, H.; Gong, Z.H.; Wang, S.B.; Lu, M.H. Genome-wide analysis of the CaHsp20 gene family in pepper: Comprehensive sequence and expression profile analysis under heat stress. Front. Plant Sci. 2015, 6, 806. [Google Scholar] [CrossRef] [PubMed]
- Han, X.W.; Han, S.; Zhu, Y.X.; Wang, F.; Gao, S.H.; Liu, Y.Q.; Yin, J.L. Genome-wide identification and expression analysis of pepper DUF966 gene family. J. South. Agric. 2023, 54, 1341–1351. [Google Scholar]
- Huang, C.; Li, X.; Nan, N.; Li, S.Y.; Wang, J.D. Research progress of plant SWEET gene family. Chin. Agric. Sci. Bull. 2022, 38, 17–26. [Google Scholar]
- Yuan, M.; Wang, S.P. Rice MtN3/saliva/SWEET family genes and their homologs in cellular organisms. Mol. Plant 2013, 6, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.Y.; Han, J.X.; Han, X.X.; Jiang, J. Genome-wide identification; phylogeny, and expression analysis of the SWEET gene family in tomato. Gene 2015, 573, 261–272. [Google Scholar] [CrossRef]
- Feng, L.; Frommer, W.B. Structure and function of SemiSWEET and SWEET sugar transporters. Trends Biochem. Sci. 2015, 40, 480–486. [Google Scholar] [CrossRef]
- Feng, L.; Frommer, W.B. Evolution of transporters: The relationship of SWEETs, PQ-loop, and PnuC transporters. Trends Biochem. Sci. 2016, 41, 118–119. [Google Scholar] [CrossRef]
- Hu, L.P.; Zhang, F.; Xu, H.; Liu, G.M.; Wang, Y.Q.; He, H.J. Research advances in the structure, function and regulation of SWEET gene family in plants. Biotechnol. Bull. 2017, 33, 27–37. [Google Scholar]
- Chen, L.Q.; Qu, X.Q.; Hou, B.H.; Sosso, D.; Osorio, S.; Fernie, A.R.; Frommer, W.B. Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science 2012, 335, 207–211. [Google Scholar] [CrossRef]
- Chu, Z.H.; Yuan, M.; Yao, J.L.; Ge, X.J.; Yuan, B.; Xu, C.G.; Li, X.H.; Fu, B.Y.; Li, Z.K.; Bennetzen, J.L.; et al. Promoter mutations of an essential gene for pollen development result in disease resistance in rice. Genes Dev. 2006, 20, 1250–1255. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.R.; Huang, S.; Feng, Y.M.; Fu, R.M.; Tang, F.J.; Zheng, L.Y.; Li, P.J.; Chao, N.; Liu, L. SWEET transporters and their potential roles in response to abiotic and biotic stresses in mulberry. Beverage Plant Res. 2023, 3, 6. [Google Scholar] [CrossRef]
- Klemens, P.A.W.; Patzke, K.; Deitmer, J.; Spinner, L.; Le Hir, R.; Bellini, C.; Bedu, M.; Chardon, F.; Krapp, A.; Neuhaus, H.E. Overexpression of the vacuolar sugar carrier AtSWEET16 modifies germination, growth, and stress tolerance in Arabidopsis. Plant Physiol. 2013, 163, 1338–1352. [Google Scholar] [CrossRef] [PubMed]
- Xuan, C.Q.; Lan, G.P.; Si, F.F.; Zeng, Z.L.; Wang, C.X.; Yadav, V.; Wei, C.H.; Zhang, X. Systematic genome-wide study and expression analysis of SWEET gene family: Sugar transporter family contributes to biotic and abiotic stimuli in watermelon. Int. J. Mol. Sci. 2021, 22, 8407. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Sugio, A.; White, F.F. Os8N3 is a host disease-susceptibility gene for bacterial blight of rice. Proc. Natl. Acad. Sci. USA 2006, 103, 10503–10508. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, Z.Y.; Kumar, V.; Xu, X.F.; Yuan, D.P.; Zhu, X.F.; Li, T.Y.; Jia, B.L.; Xuan, Y.H. Genome-wide identification of the SWEET gene family in wheat. Gene 2018, 642, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Cohn, M.; Bart, R.S.; Shybut, M.; Dahlbeck, D.; Gomez, M.; Morbitzer, R.; Hou, B.-H.; Frommer, W.B.; Lahaye, T.; Staskawicz, B.J. Xanthomonas axonopodis virulence is promoted by a transcription activator-like effector-mediated induction of a SWEET sugar transporter in cassava. Mol. Plant-Microbe Interact. MPMI 2014, 27, 1186–1198. [Google Scholar] [CrossRef]
- Hu, L.P.; Zhang, F.; Song, S.H.; Tang, X.W.; Xu, H.; Liu, G.M.; Wang, Y.Q.; He, H.J. Genome-wide identification; characterization, and expression analysis of the SWEET gene family in cucumber. J. Integr. Agric. 2017, 16, 1486–1501. [Google Scholar] [CrossRef]
- Zhao, L.J.; Yao, J.B.; Chen, W.; Li, Y.; Lü, Y.J.; Guo, Y.; Wang, J.Y.; Yuan, L.; Liu, Z.Y.; Zhang, Y.S. A genome-wide analysis of SWEET gene family in cotton and their expressions under different stresses. J. Cotton Res. 2018, 1, 7. [Google Scholar] [CrossRef]
- Liu, J.H.; Peng, T.; Dai, W.S. Critical cis-Acting Elements and Interacting Transcription Factors: Key Players Associated with Abiotic Stress Responses in Plants. Plant Mol. Biol. Report. 2014, 32, 303–317. [Google Scholar] [CrossRef]
- Zou, X.X.; Hu, B.W.; Xiong, C.; Dai, X.Z.; Liu, F.; Ou, L.J.; Yang, B.Z.; Liu, Z.B.; Suo, H.; Xu, H.; et al. Review and prospects of pepper breeding for the past 60 years in China. Acta Hortic. Sin. 2022, 49, 2099–2118. [Google Scholar]
- Du, Y.L.; Li, W.J.; Geng, J.; Li, S.Q.; Zhang, W.J.; Liu, X.X.; Hu, M.H.; Zhang, Z.N.; Fan, Y.R.; Yuan, X.K.; et al. Genome-wide identification of the SWEET gene family in Phaseolus vulgaris L. and their patterns of expression under abiotic stress. J. Plant Interact. 2022, 17, 390–403. [Google Scholar] [CrossRef]
- Patil, G.; Valliyodan, B.; Deshmukh, R.; Prince, S.; Nicander, B.; Zhao, M.; Sonah, H.; Song, L.; Lin, L.; Chaudhary, J.; et al. Soybean (Glycine max) SWEET gene family: Insights through comparative genomics, transcriptome profiling and whole genome re-sequence analysis. BMC Genom. 2015, 16, 520. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Z.; Vang, S.; Yu, J.; Wong, G.K.-S.; Wang, J. Correlation between Ka/Ks and Ks is related to substitution model and evolutionary lineage. J. Mol. Evol. 2009, 68, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, J.; Lyu, M.; Liu, Z.; Mehmood, N.; Munir, N.; Ahmed, M.A.A.; Batool, W.; Aslam, M.M.; Yuan, Y.; Wu, B. Sugar and hormone dynamics and the expression profiles of SUT/SUC and SWEET dugar transporters during flower development in Petunia axillaris. Plants 2020, 9, 1770. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.F.; Huang, X.Y.; Zhu, J.; Gao, J.F.; Zhang, H.X.; Yang, Z.N. RUPTURED POLLEN GRAIN1, a member of the MtN3/saliva gene family, is crucial for exine pattern formation and cell integrity of microspores in Arabidopsis. Plant Physiol. 2008, 147, 852–863. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Osakabe, Y.; Mizoi, J.; Nakashima, K.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of an Arabidopsis thaliana abiotic stress-inducible facilitated diffusion transporter for monosaccharides. J. Biol. Chem. 2010, 285, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.; Porcheron, B.; Hennion, N.; Maurousset, L.; Lemoine, R.; Pourtau, N. Water Deficit Enhances C Export to the Roots in Arabidopsis thaliana Plants with Contribution of Sucrose Transporters in Both Shoot and Roots. Plant Physiol. 2016, 170, 1460–1479. [Google Scholar] [CrossRef]
- Jian, H.J.; Lu, K.; Yang, B.; Wang, T.Y.; Zhang, L.; Zhang, A.X.; Wang, J.; Liu, L.Z.; Qu, C.M.; Li, J.N. Genome-wide analysis and expression profiling of the SUC and SWEET gene families of sucrose transporters in oilseed rape (Brassica napus L.). Front. Plant Sci. 2016, 7, 1464. [Google Scholar] [CrossRef]
- Hu, B.; Wu, H.; Huang, W.F.; Song, J.B.; Zhou, Y.; Lin, Y.J. SWEET gene family in medicago truncatula: Genome-wide identification, expression and substrate specificity analysis. Plants 2019, 8, 338. [Google Scholar] [CrossRef]
- Yin, J.L.; Liu, M.Y.; Ma, D.F.; Wu, J.W.; Li, S.L.; Zhu, Y.X.; Han, B. Identification of circular RNAs and their targets during tomato fruit ripening. Postharvest Biol. Technol. 2018, 136, 90–98. [Google Scholar] [CrossRef]
- Köhler, R.H. GFP for in vivo imaging of subcellular structures in plant cells. Trends Plant Sci. 1998, 3, 317–320. [Google Scholar] [CrossRef]
- Yang, L.; Xu, M.; Koo, Y.; He, J.; Poethig, R.S. Sugar promotes vegetative phase change in Arabidopsis thaliana by repressing the expression of MIR156A and MIR156C. eLife 2013, 2, e00260. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Li, X. Advances in the study of plant SWEET gene family for disease development. Jiangsu J. Agric. Sci. 2022, 38, 1411–1420. [Google Scholar]
- Chong, J.; Piron, M.-C.; Meyer, S.; Merdinoglu, D.; Bertsch, C.; Mestre, P. The SWEET family of sugar transporters in grapevine: VvSWEET4 is involved in the interaction with Botrytis cinerea. J. Exp. Bot. 2014, 65, 6589–6601. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, C.; Han, X.; Wang, Z.Y.; Ma, L.; Yuan, D.P.; Wu, J.N.; Zhu, X.F.; Liu, J.M.; Li, D.P.; et al. Inhibition of OsSWEET11 function in mesophyll cells improves resistance of rice to sheath blight disease. Mol. Plant Pathol. 2018, 19, 2149–2161. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.X.; Zhang, Z.Q.; Ren, Z.Y.; Wang, X.X.; Sun, W.J.; Feng, H.J.; Zhao, J.J.; Zhang, F.; Li, W.; Ma, X.F.; et al. The GhSWEET42 glucose transporter participates in Verticillium dahliae infection in cotton. Front. Plant Sci. 2021, 12, 690754. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.N.; Zhang, H.; Zhang, Q.; Zhai, H.; Liu, Q.C.; He, S.Z. The plasma membrane-localized sucrose tansporter IbSWEET10 contributes to the resistance of Sweet potato to Fusarium oxysporum. Front. Plant Sci. 2017, 8, 197. [Google Scholar]
- Yuan, M.; Zhao, J.W.; Huang, R.Y.; Li, X.H.; Xiao, J.H.; Wang, S.P. Rice MtN3/saliva/SWEET gene family: Evolution, expression profiling, and sugar transport. J. Integr. Plant Biol. 2014, 56, 559–570. [Google Scholar] [CrossRef]
- Li, Y.T.; Liu, X.; Xiao, Y.X.; Wen, Y.; Li, K.K.; Ma, Z.L.; Yang, L.J.; Zhu, Y.X.; Yin, J.L. Genome-wide characterization and function analysis uncovered roles of wheat LIMs in responding to adverse stresses and TaLIM8-4D function as a susceptible gene. Plant Genome 2022, 15, e20246. [Google Scholar] [CrossRef]
- Chem, Y.K.; Li, C.H.; Zhang, B.; Yi, J.; Kong, C.Y.; Lei, C.X.; Gong, M. The Role of the Late Embryogenesis-Abundant (LEA) Protein Family in Development and the Abiotic Stress Response: A Comprehensive Expression Analysis of Potato (Solanum tuberosum). Genes 2019, 10, 148. [Google Scholar] [CrossRef] [PubMed]
- Gish, W.; States, D.J. Identification of protein coding regions by database similarity search. Nat. Genet. 1993, 3, 266–272. [Google Scholar] [CrossRef]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.E.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Chou, K.-C.; Shen, H.-B. Cell-PLoc: A package of Web servers for predicting subcellular localization of proteins in various organisms. Nat. Protoc. 2008, 3, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yu, H.Y.; Deng, Y.G.; Zheng, J.Y.; Liu, M.L.; Ou, L.J.; Yang, B.Z. PepperHub, a Pepper Informatics Hub for the chili pepper research community. Mol. Plant 2017, 10, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Chan, C.-K.K. Analysis of RNA-Seq Data Using TopHat and Cufflinks. In Methods in Molecular Biology; Humana Press: Clifton, NJ, USA, 2016; Volume 1374, pp. 339–361. [Google Scholar]
- Yuan, Y.; Yin, X.H.; Han, X.W.; Han, S.; Li, Y.T.; Ma, D.F.; Fang, Z.W.; Yin, J.L.; Gong, S.J. Genome-wide identification, characterization and expression analysis of the TaDUF724 gene family in wheat (Triticum aestivum). Int. J. Mol. Sci. 2023, 24, 14248. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.L.; Yan, J.H.; Hou, L.; Jiang, L.L.; Xian, W.R.; Guo, Q.Y. Identification and functional deciphering suggested the regulatory roles of long intergenic ncRNAs (lincRNAs) in increasing grafting pepper resistance to Phytophthora capsici. BMC Genom. 2021, 22, 868. [Google Scholar] [CrossRef]
- Yin, X.H.; Yuan, Y.; Han, X.W.; Han, S.; Li, Y.T.; Ma, D.F.; Fang, Z.W.; Gong, S.J.; Yin, J.L. Genome-wide identification; characterization, and expression profiling of TaDUF668 gene family in Triticum aestivum. Agronomy 2023, 13, 2178. [Google Scholar] [CrossRef]
- Dang, F.F.; Wang, Y.; She, J.J.; Lei, Y.F.; Liu, Z.Q.; Eulgem, T.; Lai, Y.; Lin, J.; Yu, L.; Lei, D.; et al. Overexpression of CaWRKY27, a subgroup IIe WRKY transcription factor of Capsicum annuum, positively regulates tobacco resistance to Ralstonia solanacearum infection. Physiol. Plantarum. 2014, 150, 397–411. [Google Scholar] [CrossRef]
- Yin, J.L.; Gu, B.; Huang, G.Y.; Tian, Y.E.; Quan, J.L.; Lindqvist-Kreuze, H.; Shan, W.X. Conserved RXLR effector genes of Phytophthora infestans expressed at the early stage of potato infection are suppressive to host defense. Front. Plant Sci. 2017, 8, 2155. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Len. | MW. | Pi. | Ins. | GRAVY | Sub. |
|---|---|---|---|---|---|---|---|
| Capana00g002536 | CaSWEET1 | 264 | 29.75 | 9.44 | 36.89 | 0.872 | Cell membrane |
| Capana00g002537 | CaSWEET2 | 296 | 33.35 | 9.37 | 29.38 | 0.604 | Cell membrane |
| Capana00g002538 | CaSWEET3 | 300 | 33.49 | 9.28 | 31.03 | 0.691 | Cell membrane |
| Capana00g002539 | CaSWEET4 | 301 | 33.48 | 9.18 | 31.45 | 0.704 | Cell membrane |
| Capana00g002540 | CaSWEET5 | 298 | 33.55 | 8.99 | 41.64 | 0.635 | Cell membrane Chloroplast |
| Capana00g002541 | CaSWEET6 | 295 | 33.39 | 8.91 | 35.61 | 0.744 | Cell membrane |
| Capana01g000092 | CaSWEET7 | 254 | 28.30 | 9.66 | 37.25 | 0.714 | Cell membrane |
| Capana02g000800 | CaSWEET8 | 227 | 25.48 | 9.41 | 32.43 | 0.47 | Cell membrane |
| Capana02g001619 | CaSWEET9 | 829 | 93.21 | 8.72 | 46.16 | −0.227 | Nucleus |
| Capana02g003486 | CaSWEET10 | 622 | 68.21 | 5.41 | 34.45 | 0.239 | Chloroplast |
| Capana03g000977 | CaSWEET11 | 236 | 26.76 | 9.47 | 35.85 | 0.765 | Cell membrane |
| Capana03g001611 | CaSWEET12 | 298 | 33.25 | 9.43 | 39.32 | 0.7 | Cell membrane |
| Capana03g001663 | CaSWEET13 | 245 | 27.41 | 8.88 | 31.38 | 0.558 | Cell membrane |
| Capana03g002056 | CaSWEET14 | 224 | 25.32 | 8.42 | 34.05 | 0.36 | Cell membrane |
| Capana03g003556 | CaSWEET15 | 286 | 32.91 | 7.65 | 37.24 | 0.416 | Cell membrane Chloroplast |
| Capana04g001408 | CaSWEET16 | 225 | 25.14 | 9.38 | 36.98 | 0.643 | Cell membrane Chloroplast Peroxisome |
| Capana04g001409 | CaSWEET17 | 150 | 16.92 | 8.47 | 37.62 | 0.515 | Chloroplast |
| Capana04g001457 | CaSWEET18 | 240 | 26.41 | 9.56 | 39.85 | 0.411 | Cell membrane Chloroplast Golgi apparatus Peroxisome |
| Capana04g001460 | CaSWEET19 | 221 | 24.05 | 9.71 | 27.53 | 0.654 | Cell membrane |
| Capana04g002805 | CaSWEET20 | 220 | 24.81 | 9.48 | 53.11 | 0.841 | Cell membrane |
| Capana05g002066 | CaSWEET21 | 146 | 15.97 | 8.96 | 38.11 | 0.838 | Cell membrane Chloroplast Golgi apparatus Peroxisome |
| Capana06g000792 | CaSWEET22 | 282 | 31.91 | 9.25 | 39.17 | 0.662 | Cell membrane |
| Capana06g000793 | CaSWEET23 | 290 | 32.91 | 8.08 | 45.41 | 0.685 | Cell membrane Chloroplast |
| Capana06g000794 | CaSWEET24 | 222 | 25.38 | 8.77 | 34.71 | 0.69 | Cell membrane Chloroplast |
| Capana06g000962 | CaSWEET25 | 195 | 21.95 | 7.78 | 33.69 | 0.835 | Cell membrane Chloroplast |
| Capana06g001685 | CaSWEET26 | 239 | 26.75 | 9.26 | 41.52 | 0.535 | Cell membrane |
| Capana07g000747 | CaSWEET27 | 135 | 15.59 | 9.3 | 44.94 | 0.015 | Chloroplast |
| Capana08g001545 | CaSWEET28 | 156 | 17.22 | 9.42 | 38.59 | 0.585 | Cell membrane |
| Capana08g001547 | CaSWEET29 | 240 | 26.62 | 6.72 | 38.43 | 0.604 | Cell membrane |
| Capana08g001548 | CaSWEET30 | 296 | 32.56 | 9.67 | 32.29 | 0.319 | Cell membrane Chloroplast |
| Capana11g001074 | CaSWEET31 | 304 | 33.96 | 7.64 | 40.67 | 0.649 | Cell membrane |
| Capana12g001058 | CaSWEET32 | 178 | 20.16 | 9.87 | 46.67 | 0.942 | Cell membrane |
| Capana12g001557 | CaSWEET33 | 138 | 15.66 | 8.83 | 36.89 | 0.162 | Cell membrane |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, X.; Han, S.; Zhu, Y.; Liu, Y.; Gao, S.; Yin, J.; Wang, F.; Yao, M. Genome-Wide Identification and Expression Analysis of the SWEET Gene Family in Capsicum annuum L. Int. J. Mol. Sci. 2023, 24, 17408. https://doi.org/10.3390/ijms242417408
Han X, Han S, Zhu Y, Liu Y, Gao S, Yin J, Wang F, Yao M. Genome-Wide Identification and Expression Analysis of the SWEET Gene Family in Capsicum annuum L. International Journal of Molecular Sciences. 2023; 24(24):17408. https://doi.org/10.3390/ijms242417408
Chicago/Turabian StyleHan, Xiaowen, Shuo Han, Yongxing Zhu, Yiqing Liu, Shenghua Gao, Junliang Yin, Fei Wang, and Minghua Yao. 2023. "Genome-Wide Identification and Expression Analysis of the SWEET Gene Family in Capsicum annuum L." International Journal of Molecular Sciences 24, no. 24: 17408. https://doi.org/10.3390/ijms242417408