
Dynamics of Humic Acid, Silicon, and Biochar under Heavy Metal, Drought, and Salinity with Special Reference to Phytohormones, Antioxidants, and Melatonin Synthesis in Rice
 ,
,  and
and
Abstract
:1. Introduction
2. Results
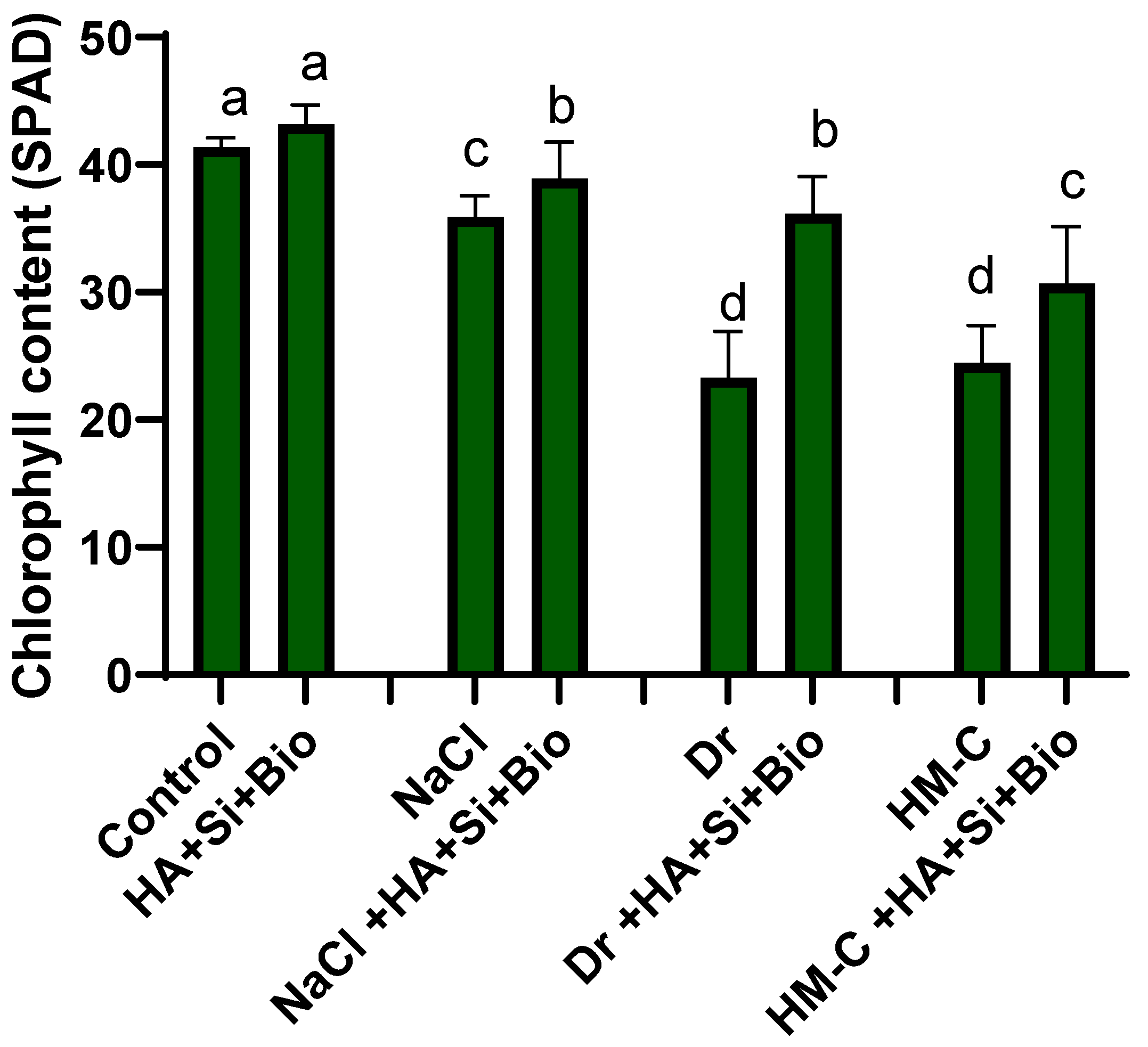
2.1. Morphological Characteristics

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | TLN | SW (g·pot−1 F.W.) | RW (g·pot−1 F.W.) | SL (cm) |
|---|---|---|---|---|
| No-Stress Condition (NS) | ||||
| Control | 15 ± 0.86 ab | 46.02 ± 0.26 a | 32.94 ± 0.54 b | 61.2 ± 0.43 a |
| HA + Si + biochar | 16.33 ± 0.57 a | 48.63 ± 0.96 a | 34.81 ± 1.07 a | 61.8 ± 0.44 a |
| Salt-Stressed Condition (SS) | ||||
| NaCl | 11 ± 1.32 c | 27.92 ± 0.34 d | 18.12 ± 0.49 e | 59.46 ± 0.20 b |
| NaCl (HA + Si + biochar) | 14 ± 0.5 b | 36.15 ± 0.34 c | 27.99 ± 0.39 c | 61.36 ± 0.8 a |
| Drought-Stressed Condition (DS) | ||||
| Drought | 14.5 ± 0.5 b | 34.92 ± 2.30 c | 25.35 ± 0.37 d | 59.2 ± 0.45 b |
| Drought (HA + Si + biochar) | 15.66 ± 0.76 ab | 41.61 ± 1.98 b | 31.92 ± 1.43 b | 61.7 ± 0.43 a |
| Combined Heavy Metal Stress Condition (HM-C) | ||||
| HM-C | 10.66 ± 1.6 c | 23.16 ± 0.34 e | 19.2 ± 1.25 e | 56.16 ± 0.32 c |
| HM-C + HA + Si + biochar | 11.66 ± 1.44 c | 27.28 ± 2.82 d | 24.16 ± 1.92 d | 56.13 ± 0.37 c |

2.2. Physiochemical Analysis
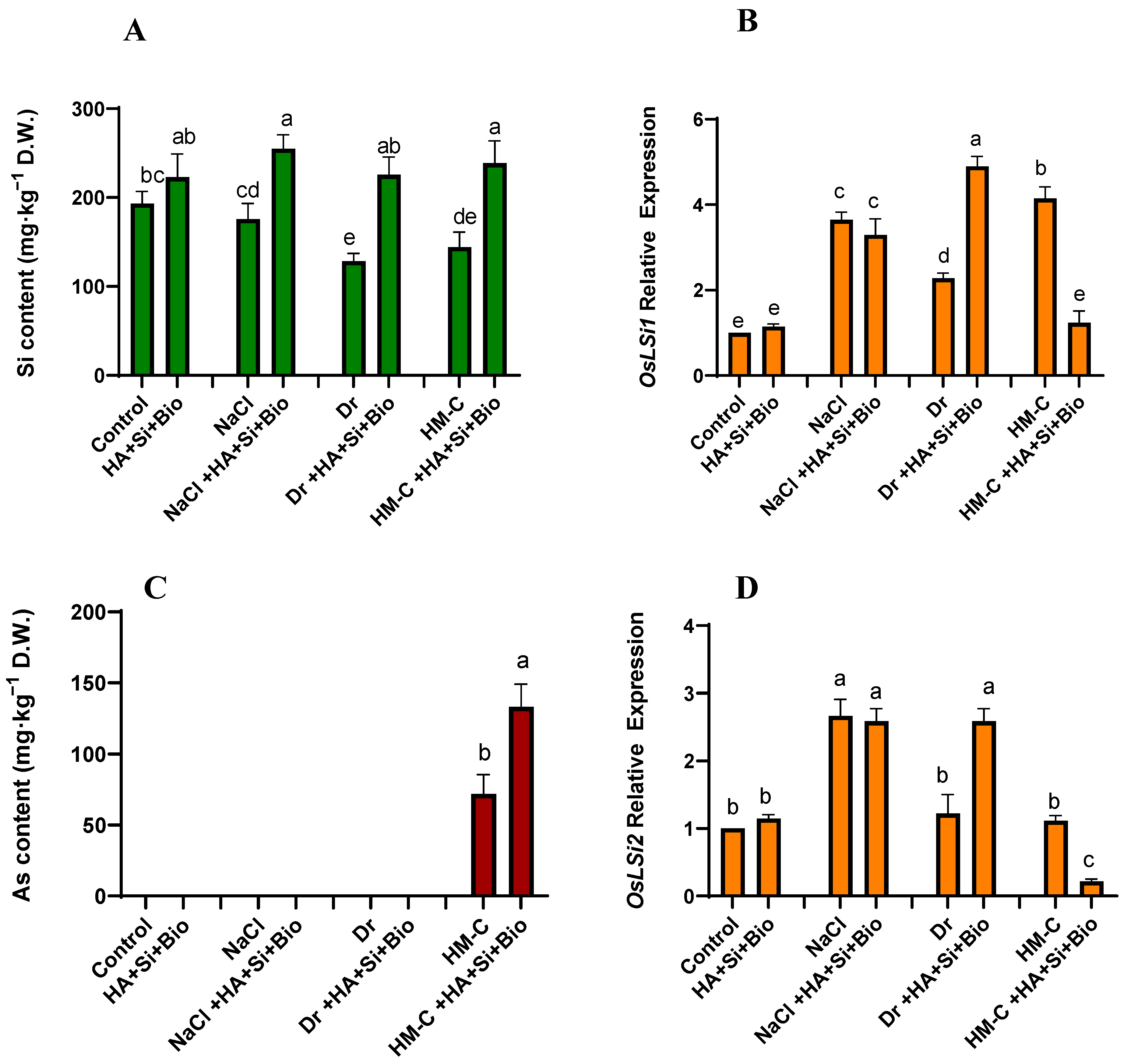
2.2.1. Quantification of Si and As and the Expression of Their Transporter Genes OsLsi1 and OsLsi2
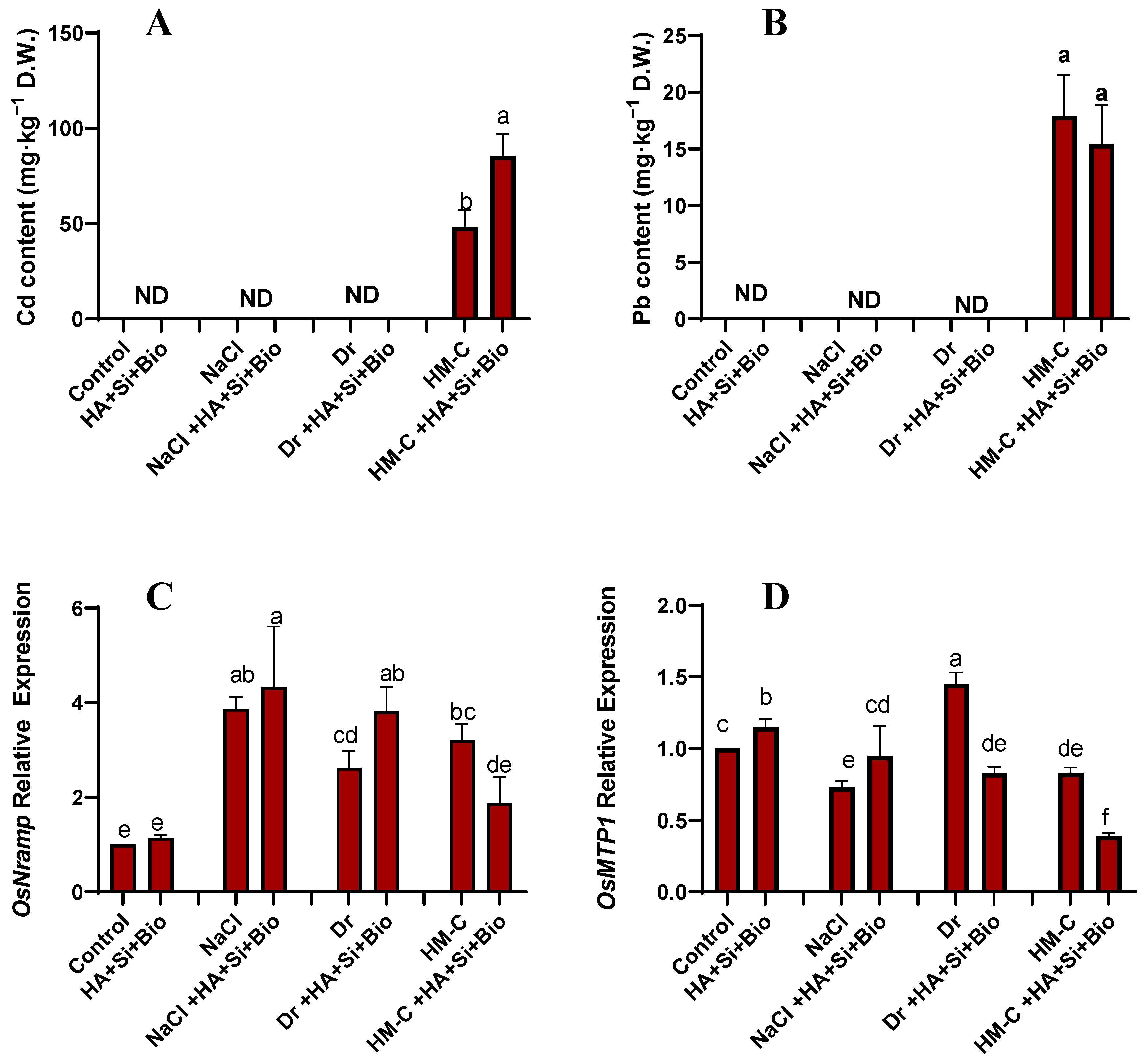
2.2.2. Quantification of Cd and Pb and the Expression of Their Related Genes OsMTP1 and OsNramp
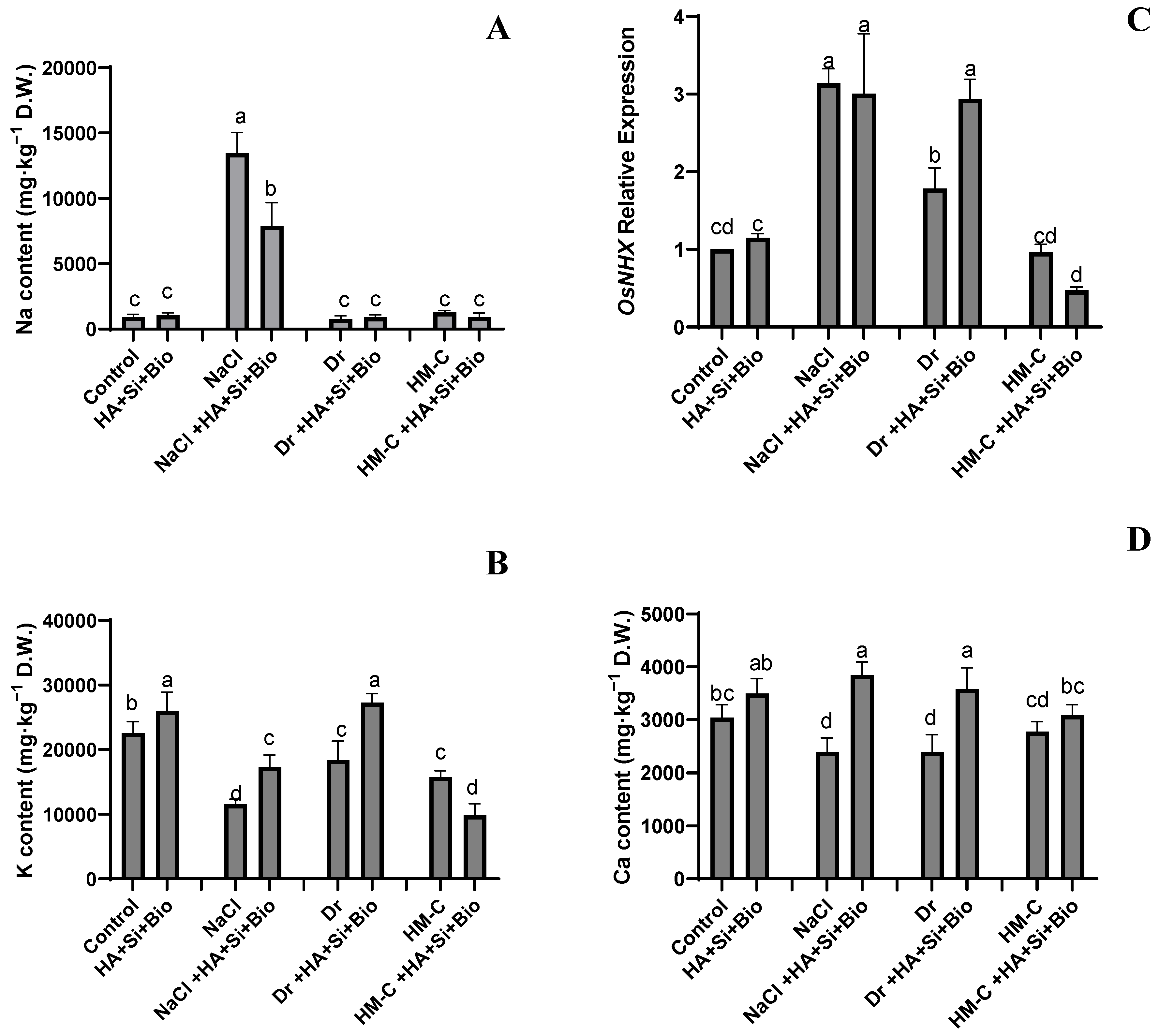
2.2.3. Analysis of Na+/K+/Ca2+ and Its Regulator OsNHX1 Expression
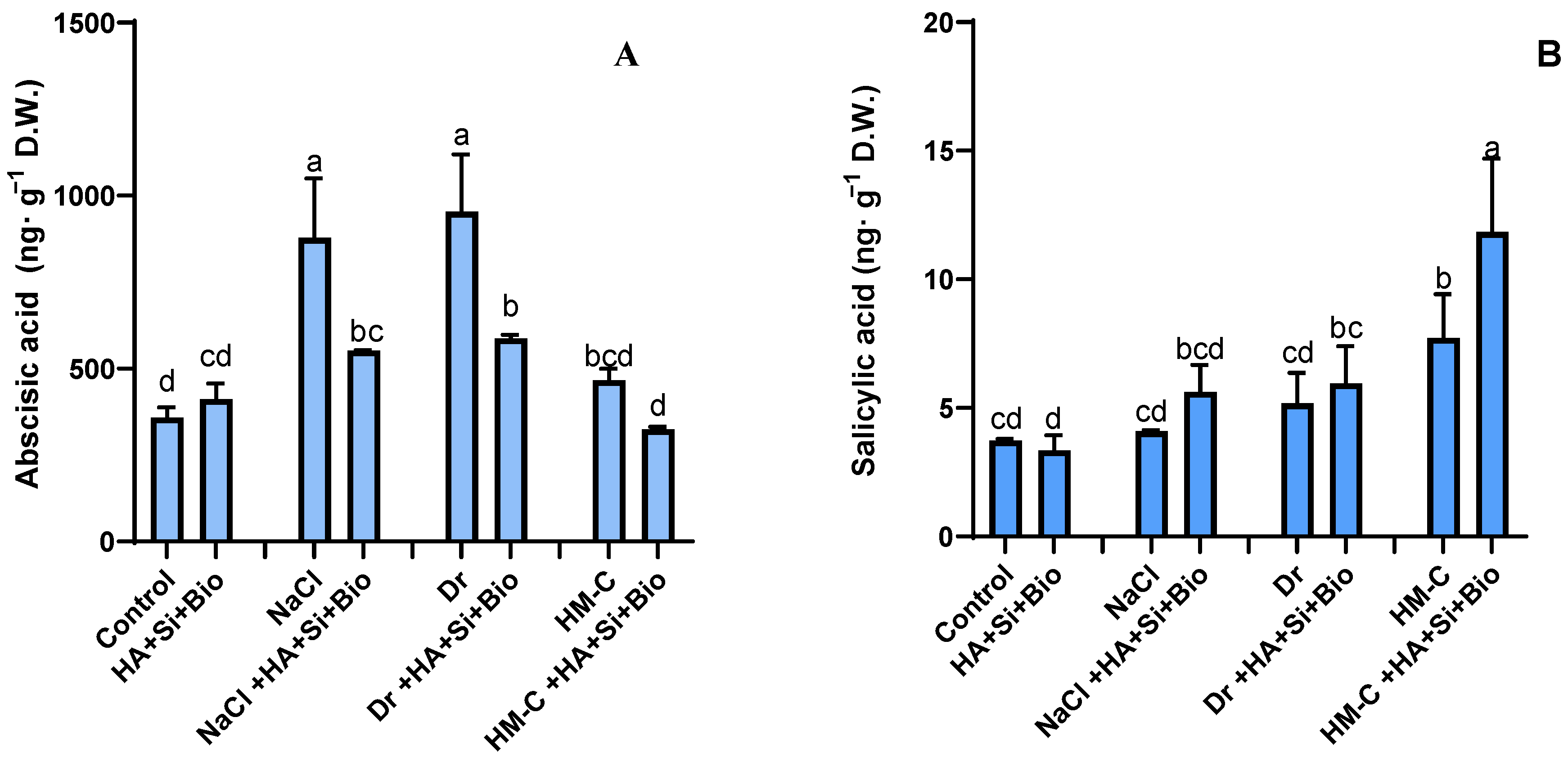
2.2.4. Analysis of Endogenous Phytohormones (ABA and SA)
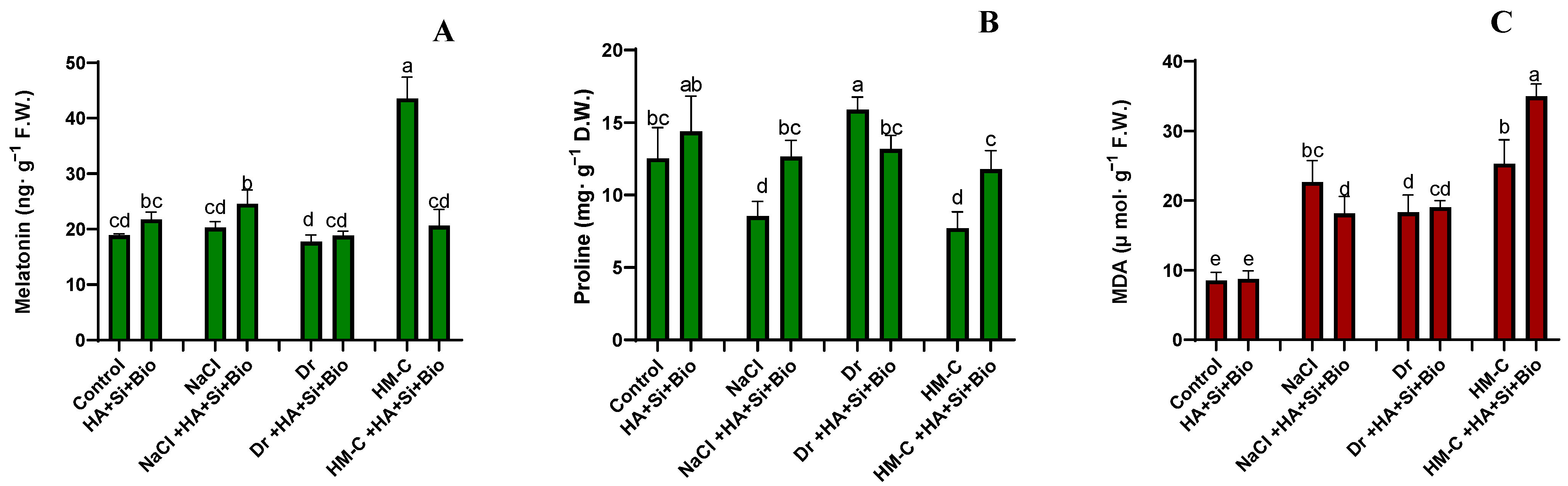
2.2.5. Quantification of the Melatonin Proline and Malondialdehyde (MDA) Levels
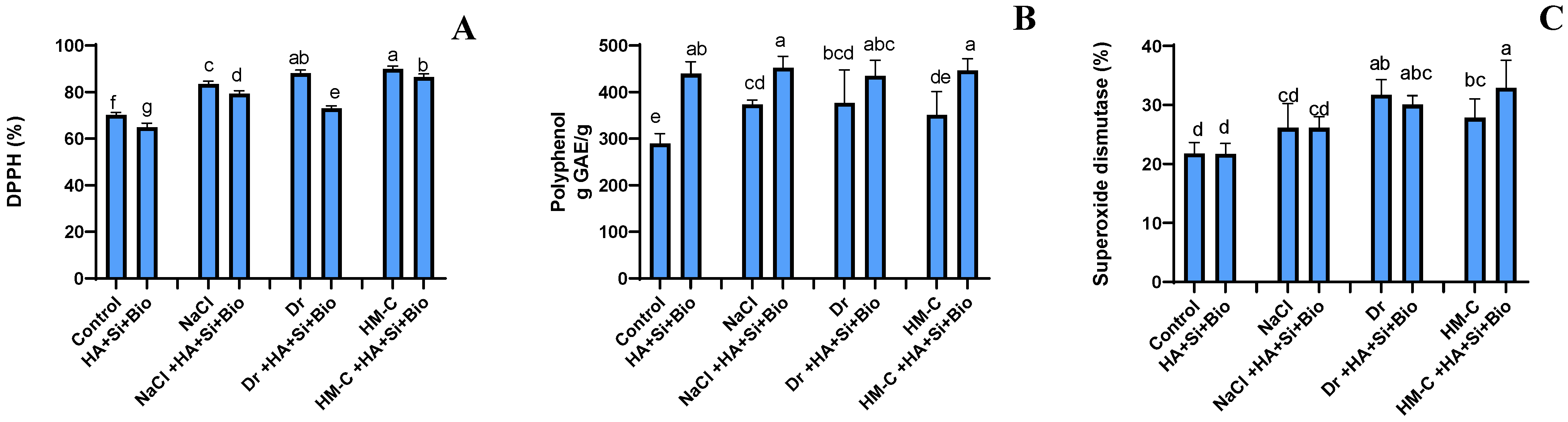
2.2.6. Antioxidant Activity Assay
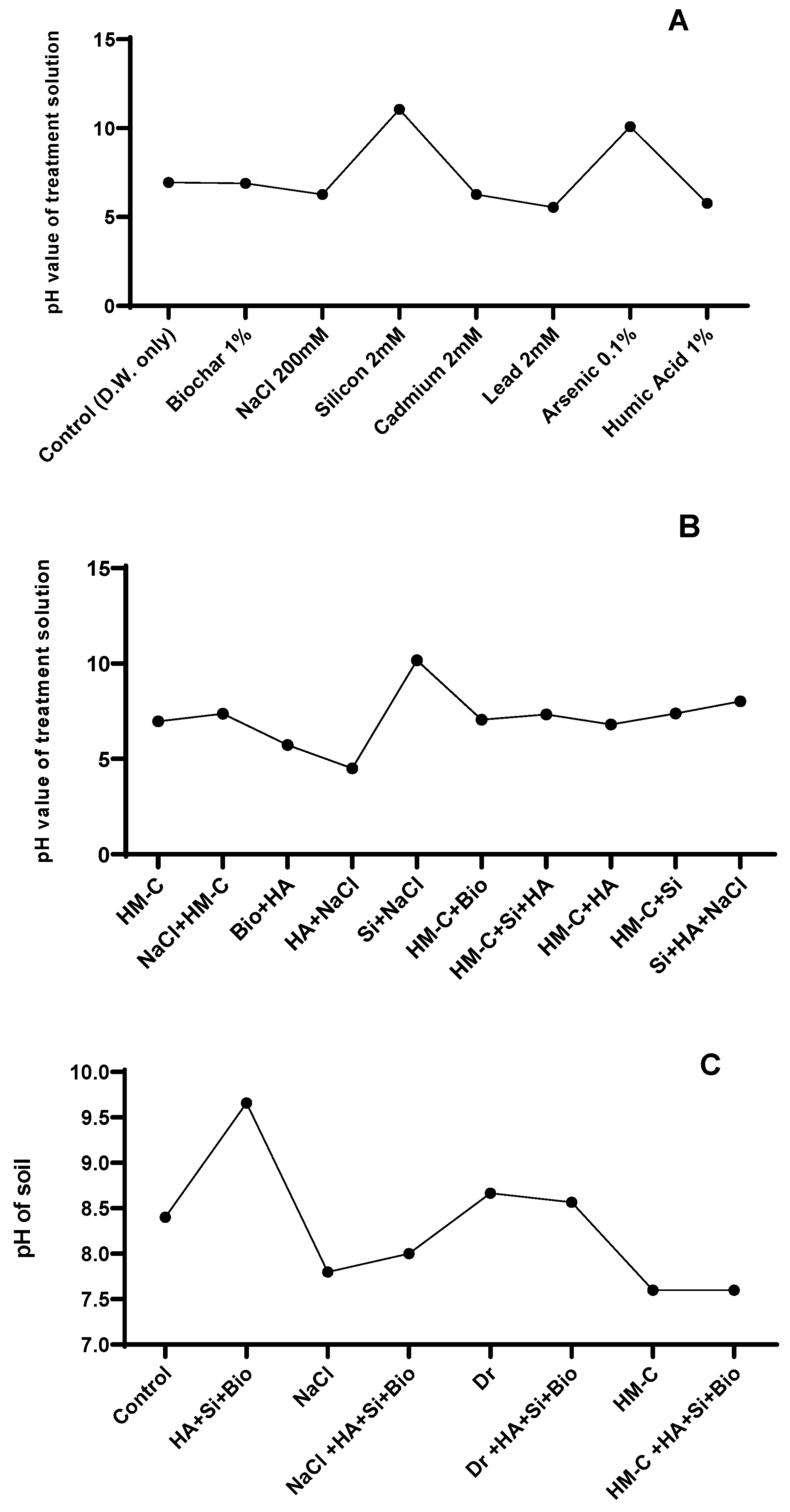
2.2.7. pH of the Soil and Treatment Solution
3. Discussion
4. Materials and Methods
4.1. Experimental Work Plan
4.2. Plant Experiments
4.2.1. Materials
4.2.2. HA, Si, and Biochar Screening
4.2.3. pH Level Measurement of the Treatment Solutions
4.2.4. Plant Experiment with Different Stresses
4.3. Biochemical Analysis of Rice Plants
4.3.1. Endogenous Phytohormone Analysis
4.3.2. Mineral Elemental Analysis
4.3.3. Proline Quantification
4.3.4. Extent of the Lipid Peroxidation MDA Analysis
4.3.5. cDNA Synthesis and Real-Time PCR for the Analysis of the Metal Transporter and Melatonin Pathways
4.3.6. Melatonin Quantification
4.3.7. Antioxidant Activity Assay (DPPH and Polyphenol)
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Kumar, M.; Saifi, Z.; Krishnananda, S.D. Decoding the physiological response of plants to stress using deep learning for forecasting crop loss due to abiotic, biotic, and climatic variables. Sci. Rep. 2023, 13, 8598. [Google Scholar] [CrossRef]
- Wang, D.; Saleh, N.B.; Byro, A.; Zepp, R.; Sahle-Demessie, E.; Luxton, T.P.; Ho, K.T.; Burgess, R.M.; Flury, M.; White, J.C.; et al. Nano-enabled pesticides for sustainable agriculture and global food security. Nat. Nanotechnol. 2022, 17, 347–360. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.; Zhu, J.-K. Developing naturally stress-resistant crops for a sustainable agriculture. Nat. Plants 2018, 4, 989–996. [Google Scholar] [CrossRef]
- Kumar, G.; Basu, S.; Singla-Pareek, S.L.; Pareek, A. Unraveling the contribution of OsSOS2 in conferring salinity and drought tolerance in a high-yielding rice. Physiol. Plant. 2022, 174, e13638. [Google Scholar] [CrossRef]
- Yadav, S.; Irfan, M.; Ahmad, A.; Hayat, S. Causes of salinity and plant manifestations to salt stress: A review. J. Environ. Biol. 2011, 32, 667. [Google Scholar]
- Skuza, L.; Szućko-Kociuba, I.; Filip, E.; Bożek, I. Natural Molecular Mechanisms of Plant Hyperaccumulation and Hypertolerance towards Heavy Metals. Int. J. Mol. Sci. 2022, 23, 9335. [Google Scholar] [CrossRef]
- Paithankar, J.G.; Saini, S.; Dwivedi, S.; Sharma, A.; Chowdhuri, D.K. Heavy metal associated health hazards: An interplay of oxidative stress and signal transduction. Chemosphere 2021, 262, 128350. [Google Scholar] [CrossRef] [PubMed]
- Tóth, G.; Hermann, T.; Da Silva, M.R.; Montanarella, L. Heavy metals in agricultural soils of the European Union with implications for food safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Khan, E. Trophic transfer, bioaccumulation, and biomagnification of non-essential hazardous heavy metals and metalloids in food chains/webs—Concepts and implications for wildlife and human health. Hum. Ecol. Risk Assess. 2019, 25, 1353–1376. [Google Scholar] [CrossRef]
- Béné, C.; Barange, M.; Subasinghe, R.; Pinstrup-Andersen, P.; Merino, G.; Hemre, G.-I.; Williams, M. Feeding 9 billion by 2050–Putting fish back on the menu. Food Secur. 2015, 7, 261–274. [Google Scholar] [CrossRef]
- Premanandh, J. Factors affecting food security and contribution of modern technologies in food sustainability. J. Sci. Food Agric. 2011, 91, 2707–2714. [Google Scholar] [CrossRef]
- Li, Y.; Chu, Y.; Sun, H.; Bao, Q.; Huang, Y. Melatonin alleviates arsenite toxicity by decreasing the arsenic accumulation in cell protoplasts and increasing the antioxidant capacity in rice. Chemosphere 2023, 312, 137292. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Min, W.; Akhtar, M.; Lu, X.; Bai, X.; Zhang, Y.; Tian, L.; Li, P. Melatonin enhances drought tolerance in rice seedlings by modulating antioxidant systems, osmoregulation, and corresponding gene expression. Int. J. Mol. Sci. 2022, 23, 12075. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Ding, D.; Bai, D.; Zhu, Y.; Sun, W.; Sun, Y.; Zhang, D. Melatonin biosynthesis pathways in nature and its production in engineered microorganisms. Synth. Syst. Biotechnol. 2022, 7, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.-D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Pardo-Hernández, M.; López-Delacalle, M.; Rivero, R.M. ROS and NO Regulation by Melatonin Under Abiotic Stress in Plants. Antioxidants 2020, 9, 1078. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Burkhardt, S.; Manchester, L.C. Melatonin in Plants. Nutr. Rev. 2001, 59, 286–290. [Google Scholar] [CrossRef]
- Posmyk, M.M.; Janas, K.M. Melatonin in plants. Acta Physiol. Plant. 2009, 31, 1–11. [Google Scholar] [CrossRef]
- Murch, S.J.; Erland, L.A.E. A systematic review of melatonin in plants: An example of evolution of literature. Front. Plant. Sci. 2021, 12, 683047. [Google Scholar] [CrossRef]
- Ye, T.; Yin, X.; Yu, L.; Zheng, S.J.; Cai, W.J.; Wu, Y.; Feng, Y.Q. Metabolic analysis of the melatonin biosynthesis pathway using chemical labeling coupled with liquid chromatography-mass spectrometry. J. Pineal Res. 2019, 66, e12531. [Google Scholar] [CrossRef] [PubMed]
- Albacete, A.A.; Martínez-Andújar, C.; Pérez-Alfocea, F. Hormonal and metabolic regulation of source–sink relations under salinity and drought: From plant survival to crop yield stability. Biotechnol. Adv. 2014, 32, 12–30. [Google Scholar] [CrossRef]
- Etesami, H.; Li, Z.; Maathuis, F.J.M.; Cooke, J. The combined use of silicon and arbuscular mycorrhizas to mitigate salinity and drought stress in rice. Environ. Exp. Bot. 2022, 201, 104955. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and future prospects on the action mechanisms in alleviating biotic and abiotic stresses in plants. Ecotoxicol. Environ. Saf. 2018, 147, 881–896. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, F.; Moosa, A.; Nazir, M.M.; Ferrante, A.; Ashraf, M.; Nafees, M.; Chen, J.; Darras, A.; Siddique, K.H.M. Biochar: An emerging recipe for designing sustainable horticulture under climate change scenarios. Front. Plant. Sci. 2022, 13, 1018646. [Google Scholar] [CrossRef] [PubMed]
- Çimrin, K.M.; Türkmen, Ö.; Turan, M.; Tuncer, B. Phosphorus and humic acid application alleviate salinity stress of pepper seedling. Afr. J. Biotechnol. 2010, 9, 5845–5851. [Google Scholar]
- Khaleda, L.; Park, H.J.; Yun, D.-J.; Jeon, J.-R.; Kim, M.G.; Cha, J.-Y.; Kim, W.-Y. Humic acid confers high-affinity K+ transporter 1-mediated salinity stress tolerance in Arabidopsis. Mol. Cells 2017, 40, 966. [Google Scholar]
- Stevenson, F.J. Humus Chemistry: Genesis, Composition, Reactions; John Wiley & Sons: Hoboken, NJ, USA, 1994. [Google Scholar]
- Li, Y.; Fang, F.; Wei, J.; Wu, X.; Cui, R.; Li, G.; Zheng, F.; Tan, D. Humic Acid Fertilizer Improved Soil Properties and Soil Microbial Diversity of Continuous Cropping Peanut: A Three-Year Experiment. Sci. Rep. 2019, 9, 12014. [Google Scholar] [CrossRef] [PubMed]
- Ozfidan-Konakci, C.; Yildiztugay, E.; Bahtiyar, M.; Kucukoduk, M. The humic acid-induced changes in the water status, chlorophyll fluorescence and antioxidant defense systems of wheat leaves with cadmium stress. Ecotoxicol. Environ. Saf. 2018, 155, 66–75. [Google Scholar] [CrossRef]
- Genuino, D.A.D.; Bataller, B.G.; Capareda, S.C.; de Luna, M.D.G. Application of artificial neural network in the modeling and optimization of humic acid extraction from municipal solid waste biochar. J. Environ. Chem. Eng. 2017, 5, 4101–4107. [Google Scholar] [CrossRef]
- Konishi, N.; Mitani-Ueno, N.; Yamaji, N.; Ma, J.F. Polar localization of a rice silicon transporter requires isoleucine at both C- and N-termini as well as positively charged residues. Plant. Cell. 2023, 35, 2232–2250. [Google Scholar] [CrossRef]
- Ma, G. Food, eating behavior, and culture in Chinese society. J. Ethn. Foods 2015, 2, 195–199. [Google Scholar] [CrossRef]
- He, X.; Batáry, P.; Zou, Y.; Zhou, W.; Wang, G.; Liu, Z.; Bai, Y.; Gong, S.; Zhu, Z.; Settele, J. Agricultural diversification promotes sustainable and resilient global rice production. Nat. Food 2023, 4, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Soni, P.; Shivhare, R.; Kaur, A.; Bansal, S.; Sonah, H.; Deshmukh, R.; Giri, J.; Lata, C.; Ram, H. Reference gene identification for gene expression analysis in rice under different metal stress. J. Biotechnol. 2021, 332, 83–93. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 1–12. [Google Scholar] [CrossRef]
- Zhao, K.; Yang, Y.; Peng, H.; Zhang, L.; Zhou, Y.; Zhang, J.; Du, C.; Liu, J.; Lin, X.; Wang, N.; et al. Silicon fertilizers, humic acid and their impact on physicochemical properties, availability and distribution of heavy metals in soil and soil aggregates. Sci. Total. Environ. 2022, 822, 153483. [Google Scholar] [CrossRef]
- Wang, Y.; Xiao, X.; Zhang, K.; Chen, B. Effects of biochar amendment on the soil silicon cycle in a soil-rice ecosystem. Environ. Pollut. 2019, 248, 823–833. [Google Scholar] [CrossRef]
- Ali, S.; Rizwan, M.; Qayyum, M.F.; Ok, Y.S.; Ibrahim, M.; Riaz, M.; Arif, M.S.; Hafeez, F.; Al-Wabel, M.I.; Shahzad, A.N. Biochar soil amendment on alleviation of drought and salt stress in plants: A critical review. Environ. Sci. Pollut. Res. 2017, 24, 12700–12712. [Google Scholar] [CrossRef]
- Esringü, A.; Kaynar, D.; Turan, M.; Ercisli, S. Ameliorative Effect of Humic Acid and Plant Growth-Promoting Rhizobacteria (PGPR) on Hungarian Vetch Plants under Salinity Stress. Commun. Soil. Sci. Plant. Anal. 2016, 47, 602–618. [Google Scholar] [CrossRef]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Butt, M.; Irfan, M.; Rizwan, M.S.; Ali, H.; Cheema, M.A. Interactive Effect of Biochar and Silicon on Improving Morpho-Physiological and Biochemical Attributes of Maize by Reducing Drought Hazards. J. Soil. Sci. Plant. Nutr. 2020, 20, 1819–1826. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef]
- Khan, E.; Gupta, M. Arsenic–silicon priming of rice (Oryza sativa L.) seeds influence mineral nutrient uptake and biochemical responses through modulation of Lsi-1, Lsi-2, Lsi-6 and nutrient transporter genes. Sci. Rep. 2018, 8, 10301. [Google Scholar] [CrossRef]
- Kumar, N.; Dubey, A.K.; Upadhyay, A.K.; Gautam, A.; Ranjan, R.; Srikishna, S.; Sahu, N.; Behera, S.K.; Mallick, S. GABA accretion reduces Lsi-1 and Lsi-2 gene expressions and modulates physiological responses in Oryza sativa to provide tolerance towards arsenic. Sci. Rep. 2017, 7, 8786. [Google Scholar] [CrossRef]
- Pang, Z.; Yin, W.; Wang, Y.; Zeng, W.; Peng, H.; Liang, Y. Silicon-phosphorus pathway mitigates heavy metal stress by buffering rhizosphere acidification. Sci. Total. Environ. 2023, 904, 166887. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Cai, Z.; Xian, P.; Yang, Y.; Cheng, Y.; Lian, T.; Ma, Q.; Nian, H. Silicon-enhanced tolerance to cadmium toxicity in soybean by enhancing antioxidant defense capacity and changing cadmium distribution and transport. Ecotoxicol. Environ. Saf. 2022, 241, 113766. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-Z.; Fan, W.-J.; Lin, Y.-J. Analysis of gene expression profiles for metal tolerance protein in rice seedlings exposed to both the toxic hexavalent chromium and trivalent chromium. Int. Biodeterior. Biodegrad. 2018, 129, 102–108. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Nakanishi, H.; Nishizawa, N.K. Role of the iron transporter OsNRAMP1 in cadmium uptake and accumulation in rice. Plant. Signal. Behav. 2011, 6, 1813–1816. [Google Scholar] [CrossRef]
- Das, N.; Bhattacharya, S.; Maiti, M.K. Enhanced cadmium accumulation and tolerance in transgenic tobacco overexpressing rice metal tolerance protein gene OsMTP1 is promising for phytoremediation. Plant. Physiol. Biochem. 2016, 105, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Sharma, D.; Dwivedi, S.; Singh, M.; Tripathi, R.D.; Trivedi, P.K. Expression in Arabidopsis and cellular localization reveal involvement of rice NRAMP, OsNRAMP 1, in arsenic transport and tolerance. Plant Cell Environ. 2014, 37, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Nevo, Y.; Nelson, N. The NRAMP family of metal-ion transporters. Biochim. Biophys. Acta (BBA)-Mol. Cell. Res. 2006, 1763, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Yan, G.; Fan, X.; Zheng, W.; Gao, Z.; Yin, C.; Li, T.; Liang, Y. Silicon alleviates salt stress-induced potassium deficiency by promoting potassium uptake and translocation in rice (Oryza sativa L.). J. Plant. Physiol. 2021, 258, 153379. [Google Scholar] [CrossRef] [PubMed]
- Solis, C.A.; Yong, M.-T.; Zhou, M.; Venkataraman, G.; Shabala, L.; Holford, P.; Shabala, S.; Chen, Z.-H. Evolutionary Significance of NHX Family and NHX1 in Salinity Stress Adaptation in the Genus Oryza. Int. J. Mol. Sci. 2022, 23, 2092. [Google Scholar] [CrossRef] [PubMed]
- Saidimoradi, D.; Ghaderi, N.; Javadi, T. Salinity stress mitigation by humic acid application in strawberry (Fragaria x ananassa Duch.). Sci. Hortic. 2019, 256, 108594. [Google Scholar] [CrossRef]
- García, A.C.; Santos, L.A.; Izquierdo, F.G.; Rumjanek, V.M.; Castro, R.N.; dos Santos, F.S.; de Souza, L.G.A.; Berbara, R.L.L. Potentialities of vermicompost humic acids to alleviate water stress in rice plants (Oryza sativa L.). J. Geochem. Explor. 2014, 136, 48–54. [Google Scholar] [CrossRef]
- Hassanein, R.A.; Hussein, O.S.; Farag, I.A.; Hassan, Y.E.; Abdelkader, A.F.; Ibrahim, M. Salt-Stressed Coriander (Coriandrum sativum L.) Responses to Potassium Silicate, Humic Acid and Gamma Irradiation Pretreatments. Agronomy 2022, 12, 2268. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant. Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Li, J.; Liu, J.; Zhu, T.; Zhao, C.; Li, L.; Chen, M. The Role of Melatonin in Salt Stress Responses. Int. J. Mol. Sci. 2019, 20, 1735. [Google Scholar] [CrossRef]
- Altaf, M.A.; Hao, Y.; Shu, H.; Mumtaz, M.A.; Cheng, S.; Alyemeni, M.N.; Ahmad, P.; Wang, Z. Melatonin enhanced the heavy metal-stress tolerance of pepper by mitigating the oxidative damage and reducing the heavy metal accumulation. J. Hazard. Mater. 2023, 454, 131468. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, S.; Ma, L.; Kong, L.; Pan, S.; Tang, X.; Tian, H.; Duan, M.; Mo, Z. Effect of Exogenous Melatonin Application on the Grain Yield and Antioxidant Capacity in Aromatic Rice under Combined Lead–Cadmium Stress. Antioxidants 2022, 11, 776. [Google Scholar] [CrossRef]
- Lee, K.; Choi, G.H.; Back, K. Cadmium-induced melatonin synthesis in rice requires light, hydrogen peroxide, and nitric oxide: Key regulatory roles for tryptophan decarboxylase and caffeic acid O-methyltransferase. J. Pineal Res. 2017, 63, e12441. [Google Scholar] [CrossRef] [PubMed]
- Postiglione, A.E.; Muday, G.K. The role of ROS homeostasis in ABA-induced guard cell signaling. Front. Plant. Sci. 2020, 11, 968. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Zhang, L.; Fu, G.; Yang, Y.; Zhu, C.; Tao, L. Drought-induced proline accumulation is uninvolved with increased nitric oxide, which alleviates drought stress by decreasing transpiration in rice. J. Plant. Res. 2012, 125, 155–164. [Google Scholar] [CrossRef]
- Larkindale, J.; Huang, B. Thermotolerance and antioxidant systems in Agrostis stolonifera: Involvement of salicylic acid, abscisic acid, calcium, hydrogen peroxide, and ethylene. J. Plant. Physiol. 2004, 161, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhang, J. Cross-talk between calcium and reactive oxygen species originated from NADPH oxidase in abscisic acid-induced antioxidant defence in leaves of maize seedlings. Plant Cell Environ. 2003, 26, 929–939. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant. Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Sánchez, J.C.; García, R.F.; Cors, M.T.M. 1, 1-Diphenyl-2-picrylhydrazyl radical and superoxide anion scavenging activity of Rhizophora mangle (L.) bark. Pharmacogn. Res. 2010, 2, 279. [Google Scholar] [CrossRef]
- Adhikari, A.; Khan, M.A.; Imran, M.; Lee, K.-E.; Kang, S.-M.; Shin, J.Y.; Joo, G.-J.; Khan, M.; Yun, B.-W.; Lee, I.-J. The Combined Inoculation of Curvularia lunata AR11 and Biochar Stimulates Synthetic Silicon and Potassium Phosphate Use Efficiency, and Mitigates Salt and Drought Stresses in Rice. Front. Plant. Sci. 2022, 13, 816858. [Google Scholar] [CrossRef]
- Kim, L.-R.; Adhikari, A.; Kang, Y.; Gam, H.-J.; Kang, S.-M.; Kim, K.-Y.; Lee, I.-J. Investigation of Solanum carolinense Dominance and Phytotoxic Effect in Festuca arundinacea with Special Reference to Allelochemical Identification, Analysis of Phytohormones and Antioxidant Mechanisms. Agronomy 2022, 12, 1954. [Google Scholar] [CrossRef]
- Seskar, M.; Shulaev, V.; Raskin, I. Endogenous methyl salicylate in pathogen-inoculated tobacco plants. Plant. Physiol. 1998, 116, 387–392. [Google Scholar] [CrossRef]
- Gupta, P.; Rani, R.; Chandra, A.; Kumar, V. Potential applications of Pseudomonas sp. (strain CPSB21) to ameliorate Cr6+ stress and phytoremediation of tannery effluent contaminated agricultural soils. Sci. Rep. 2018, 8, 4860. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.-H.; Adhikari, A.; Imran, M.; Lee, D.-S.; Lee, C.-Y.; Kang, S.-M.; Lee, I.-J. Exogenous SA Applications Alleviate Salinity Stress via Physiological and Biochemical changes in St John’s Wort Plants. Plants 2023, 12, 310. [Google Scholar] [CrossRef]
- Wang, L.; Chen, W.J.; Wang, Q.; Eneji, A.E.; Li, Z.H.; Duan, L.S. Coronatine enhances chilling tolerance in cucumber (Cucumis sativus L.) seedlings by improving the antioxidative defence system. J. Agron. Crop. Sci. 2009, 195, 377–383. [Google Scholar]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Adhikari, B.; Dhungana, S.K.; Ali, M.W.; Adhikari, A.; Kim, I.-D.; Shin, D.-H. Resveratrol, total phenolic and flavonoid contents, and antioxidant potential of seeds and sprouts of Korean peanuts. Food Sci. Biotechnol. 2018, 27, 1275–1284. [Google Scholar] [CrossRef]







| No-Stress Condition (NS) | |
|---|---|
| T1 | Control |
| T2 | HA + Si + biochar |
| Salt-Stressed Condition (SS) | |
| T3 | NaCl only |
| T4 | NaCl (HA + Si + biochar) |
| Drought-Stressed Condition (DS) | |
| T5 | Dr only |
| T6 | Dr (HA + Si + biochar) |
| Combined Heavy Metal Stress Condition (HM-C) | |
| T7 | HM-C only |
| T8 | HM-C (HA + Si + biochar) |
| Experiment 1 (NaCl) | Experiment 2 (Arsenic) | Experiment 3 (Drought) | Experiment 4 (As, Pb, and Cd) |
|---|---|---|---|
| No-Stress Condition (NS) | |||
| Control | Control | Control | Control |
| HA | HA | HA | HA |
| Si | Biochar | Si | Si |
| HA + Si | HA + Biochar | HA + Si | HA + Si |
| Stressed Condition (SS, DS, and HM-C) | |||
| NaCl | As | Drought | HM-C |
| HA + NaCl | HA + As | HA + Drought | HA + HM-C |
| Si + NaCl | Biochar + AS | Si + Drought | Si + HM-C |
| HA + Si + NaCl | HA + Biochar + As | HA + Si + Drought | HA + Si + HM-C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adhikari, A.; Aneefi, A.G.; Sisuvanh, H.; Singkham, S.; Pius, M.V.; Akter, F.; Kwon, E.-H.; Kang, S.-M.; Woo, Y.-J.; Yun, B.-W.; et al. Dynamics of Humic Acid, Silicon, and Biochar under Heavy Metal, Drought, and Salinity with Special Reference to Phytohormones, Antioxidants, and Melatonin Synthesis in Rice. Int. J. Mol. Sci. 2023, 24, 17369. https://doi.org/10.3390/ijms242417369
Adhikari A, Aneefi AG, Sisuvanh H, Singkham S, Pius MV, Akter F, Kwon E-H, Kang S-M, Woo Y-J, Yun B-W, et al. Dynamics of Humic Acid, Silicon, and Biochar under Heavy Metal, Drought, and Salinity with Special Reference to Phytohormones, Antioxidants, and Melatonin Synthesis in Rice. International Journal of Molecular Sciences. 2023; 24(24):17369. https://doi.org/10.3390/ijms242417369
Chicago/Turabian StyleAdhikari, Arjun, Appiah Gregory Aneefi, Hairkham Sisuvanh, Santivong Singkham, Masele Valentine Pius, Farida Akter, Eun-Hae Kwon, Sang-Mo Kang, Youn-Ji Woo, Byung-Wook Yun, and et al. 2023. "Dynamics of Humic Acid, Silicon, and Biochar under Heavy Metal, Drought, and Salinity with Special Reference to Phytohormones, Antioxidants, and Melatonin Synthesis in Rice" International Journal of Molecular Sciences 24, no. 24: 17369. https://doi.org/10.3390/ijms242417369