
Distinct Roles for COMPASS Core Subunits Set1, Trx, and Trr in the Epigenetic Regulation of Drosophila Heart Development
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
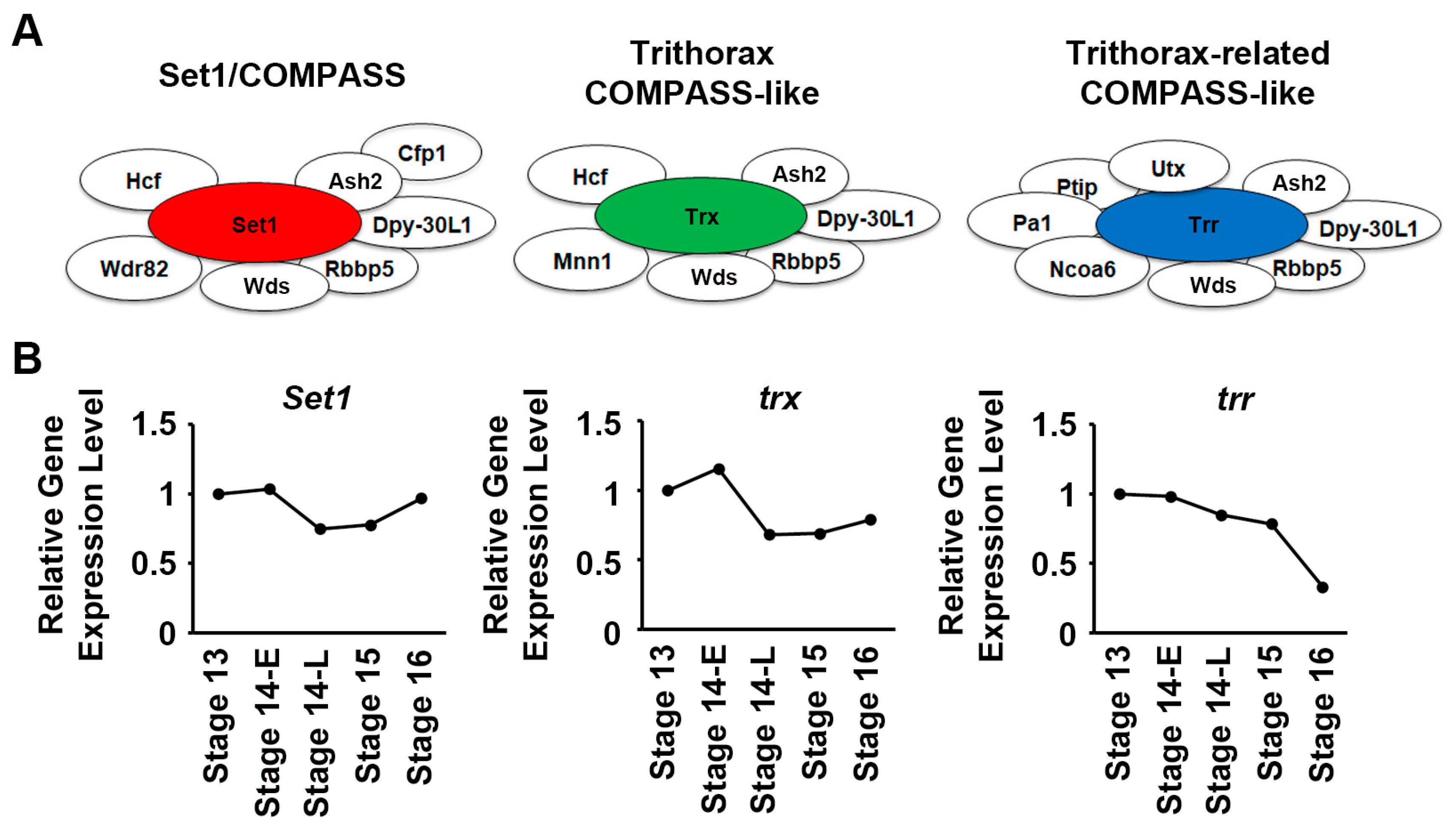
2.1. Temporal Expression Patterns of Set1, Trx, and Trr across Embryonic Heart Developmental Stages in Flies
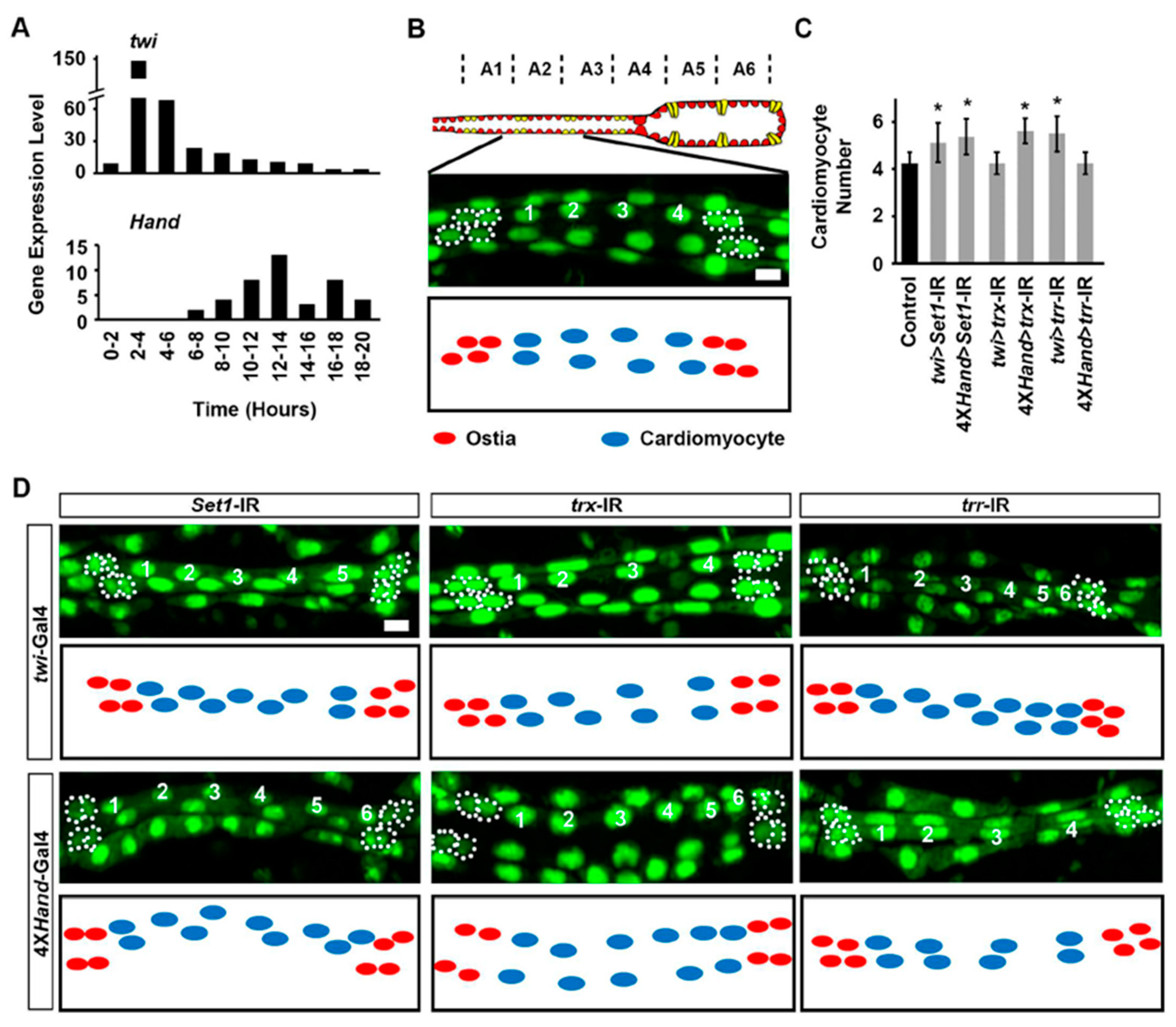
2.2. Set1, Trx, and Trr Silencing Impacted Larval Heart Development at Different Larval Stages

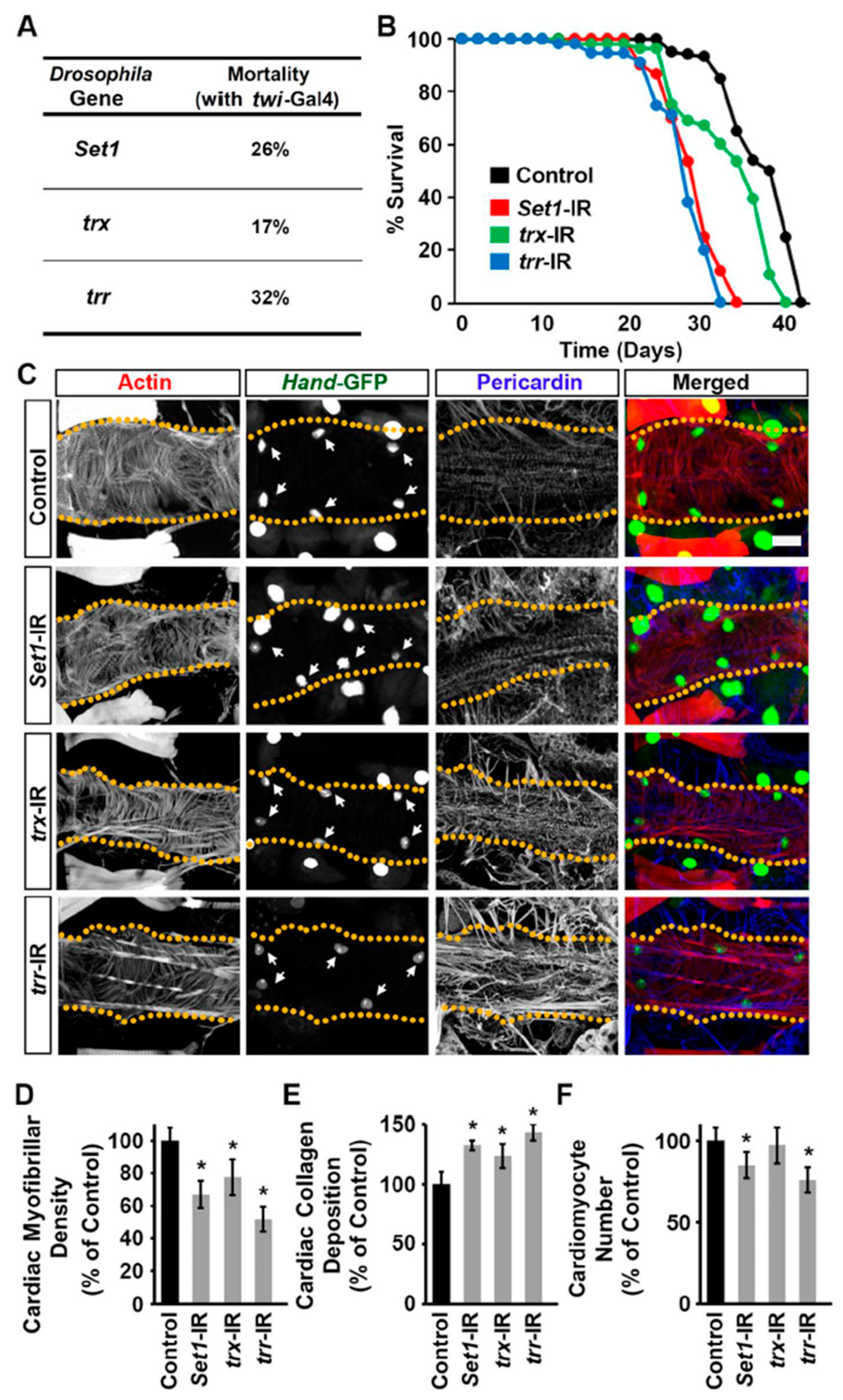
2.3. Set1, Trx, and Trr Silencing Impacted Survival and Cardiac Defects in Adult Drosophila

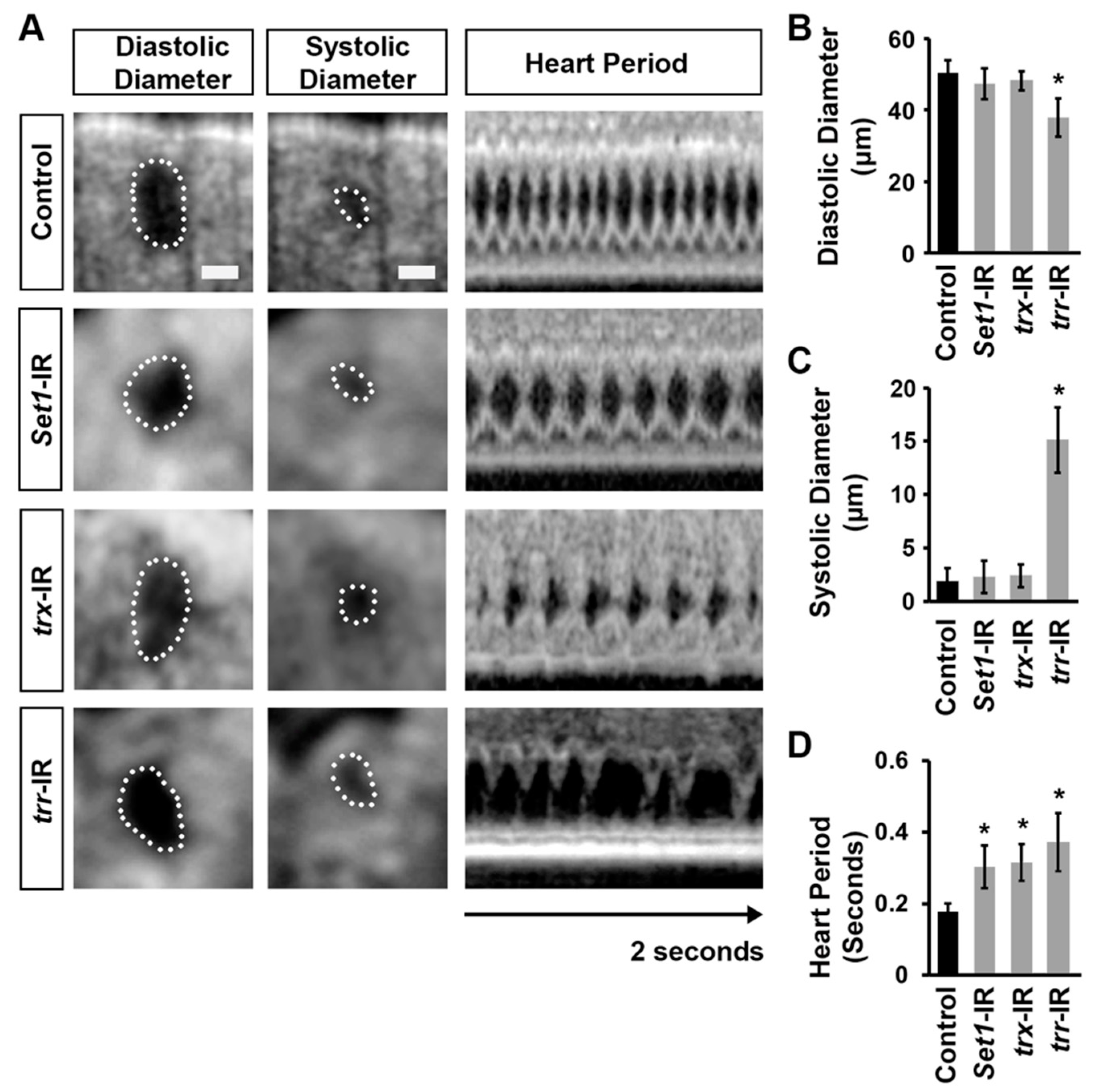
2.4. Set1, Trx, and Trr Silencing Induced Cardiac Dysfunction

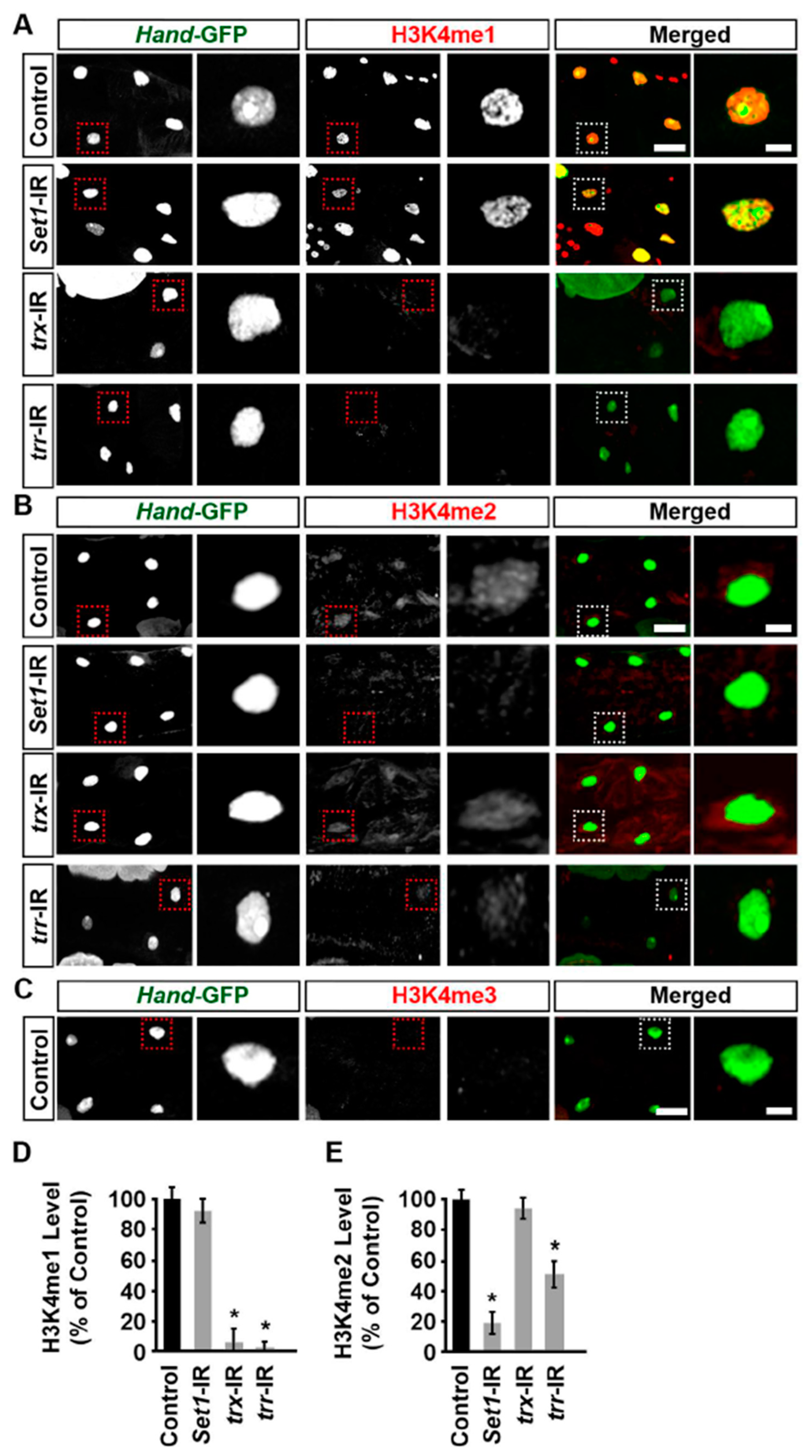
2.5. Set1, Trx, and Trr Regulated H3K4 Mono- and Dimethylation Levels in the Drosophila Heart

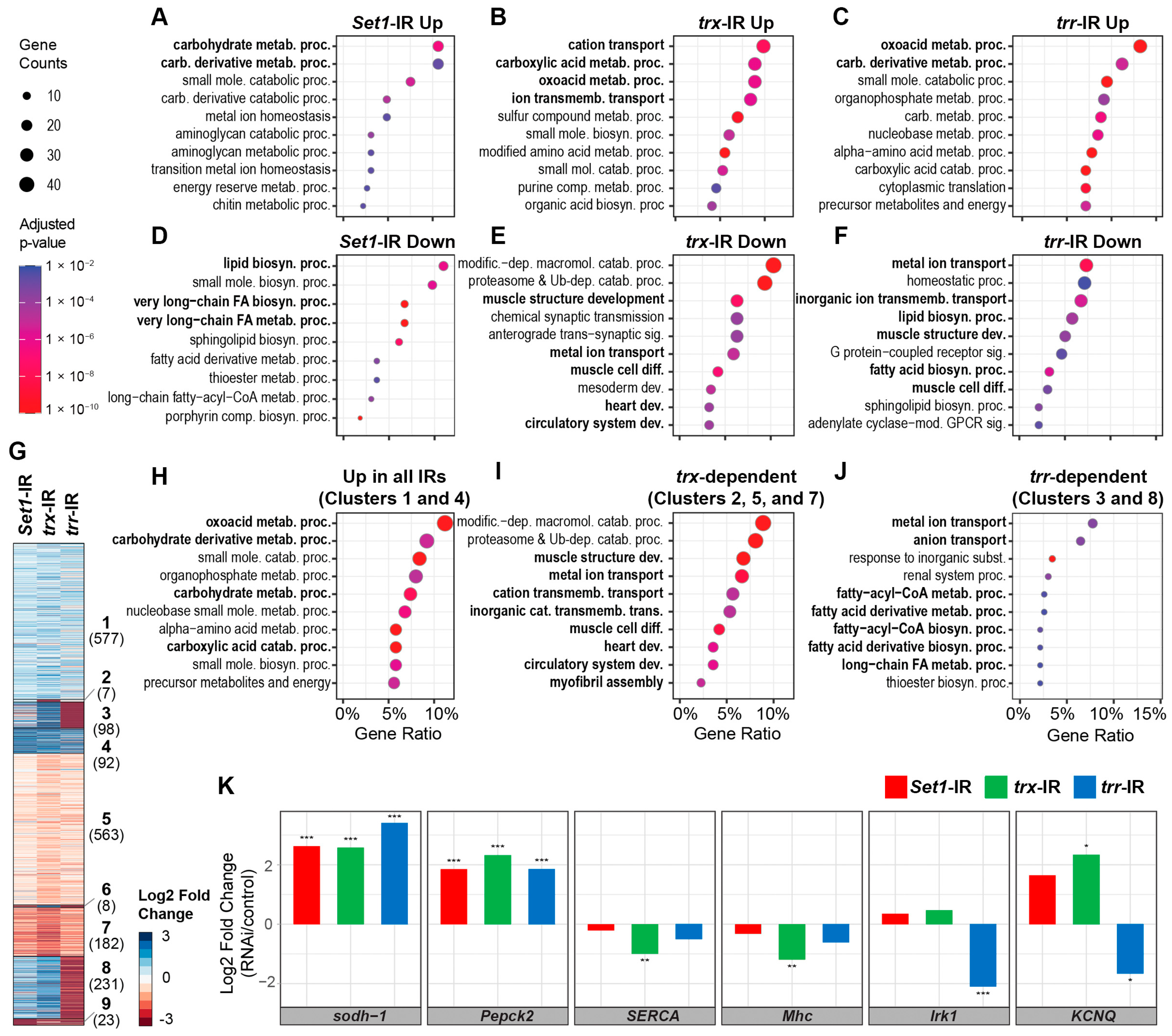
2.6. Silencing Set1, Trx, or Trr Led to Differential Transcriptomic Responses in the Drosophila Heart

3. Discussion
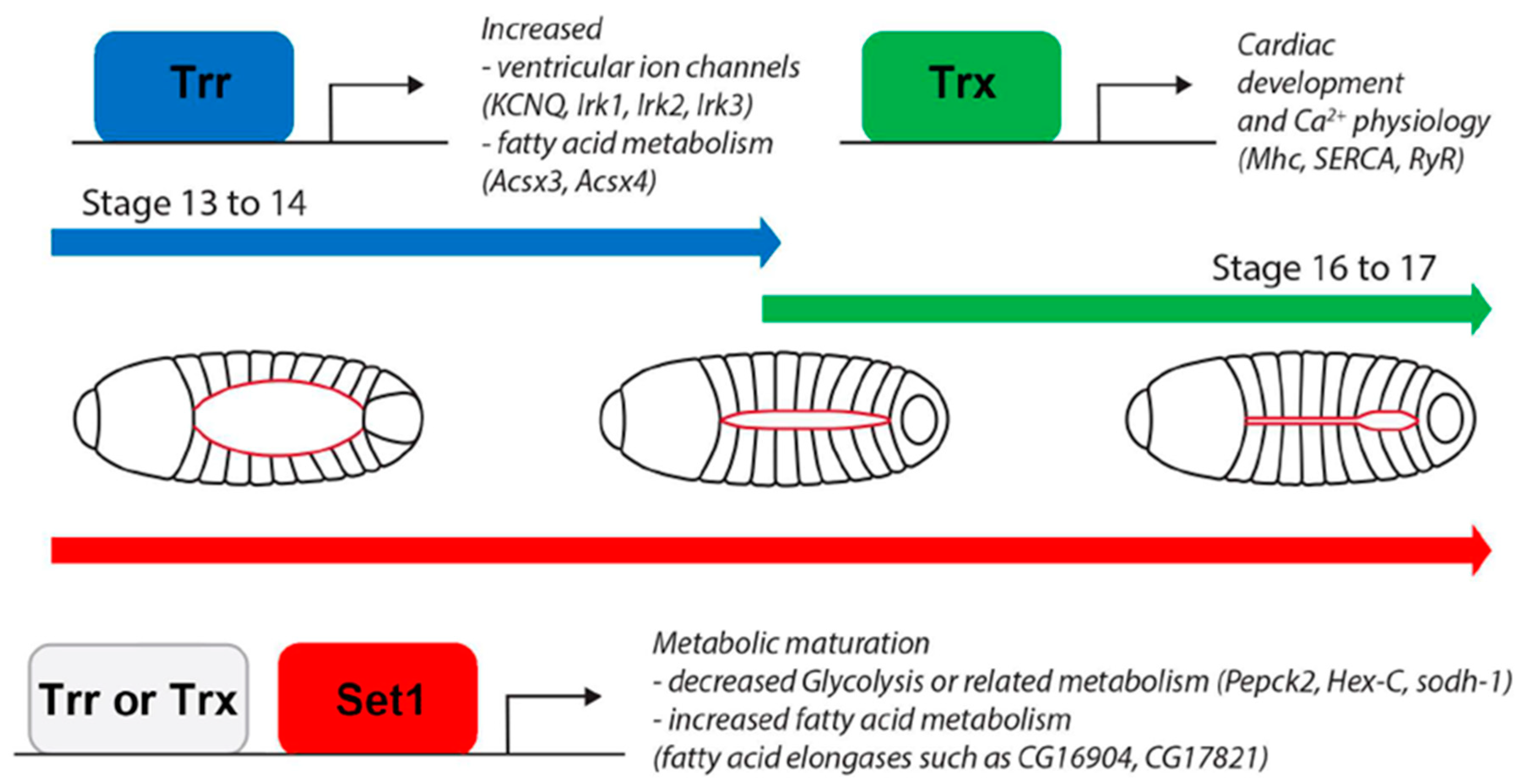
3.1. Set1, Trx, and Trr Distinctly Contribute to Methylation during Drosophila Heart Development
3.2. COMPASS Complex-Mediated Methylation during Heart Development and Disease
4. Materials and Methods
4.1. Drosophila Lines
4.2. Live Imaging of Drosophila Larva
4.3. Immunochemistry
4.4. Heart Structural Analysis and Quantitation
4.5. Lethality at Eclosion
4.6. Adult Drosophila Survival Assay
4.7. Optical Coherence Tomography (OCT)
4.8. Methylation Levels
4.9. RNA Extraction and Sequencing of Drosophila Cardiomyocytes
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, P.; Hang, C.T.; Yang, J.; Chang, C.-P. Chromatin Remodeling in Cardiovascular Development and Physiology. Circ. Res. 2011, 108, 378–396. [Google Scholar] [CrossRef]
- Ohtani, K.; Dimmeler, S. Epigenetic Regulation of Cardiovascular Differentiation. Cardiovasc. Res. 2011, 90, 404–412. [Google Scholar] [CrossRef]
- van Weerd, J.H.; Koshiba-Takeuchi, K.; Kwon, C.; Takeuchi, J.K. Epigenetic Factors and Cardiac Development. Cardiovasc. Res. 2011, 91, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-Y.; van de Leemput, J.; Han, Z. The Roles of Histone Lysine Methyltransferases in Heart Development and Disease. J. Cardiovasc. Dev. Dis. 2023, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Collins, B.E.; Greer, C.B.; Coleman, B.C.; Sweatt, J.D. Histone H3 Lysine K4 Methylation and Its Role in Learning and Memory. Epigenet. Chromatin 2019, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, S.; Choi, M.; Wakimoto, H.; Ma, L.; Jiang, J.; Overton, J.D.; Romano-Adesman, A.; Bjornson, R.D.; Breitbart, R.E.; Brown, K.K.; et al. De Novo Mutations in Histone-Modifying Genes in Congenital Heart Disease. Nature 2013, 498, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Shilatifard, A. The COMPASS Family of Histone H3K4 Methylases: Mechanisms of Regulation in Development and Disease Pathogenesis. Annu. Rev. Biochem. 2012, 81, 65–95. [Google Scholar] [CrossRef] [PubMed]
- Mohan, M.; Herz, H.-M.; Smith, E.R.; Zhang, Y.; Jackson, J.; Washburn, M.P.; Florens, L.; Eissenberg, J.C.; Shilatifard, A. The COMPASS Family of H3K4 Methylases in Drosophila. Mol. Cell. Biol. 2011, 31, 4310–4318. [Google Scholar] [CrossRef]
- Takahashi, Y.-H.; Westfield, G.H.; Oleskie, A.N.; Trievel, R.C.; Shilatifard, A.; Skiniotis, G. Structural Analysis of the Core COMPASS Family of Histone H3K4 Methylases from Yeast to Human. Proc. Natl. Acad. Sci. USA 2011, 108, 20526–20531. [Google Scholar] [CrossRef]
- Huang, W.; Zhu, J.-Y.; Fu, Y.; van de Leemput, J.; Han, Z. Lpt, Trr, and Hcf Regulate Histone Mono- and Dimethylation That Are Essential for Drosophila Heart Development. Dev. Biol. 2022, 490, 53–65. [Google Scholar] [CrossRef]
- Szulik, M.W.; Davis, K.; Bakhtina, A.; Azarcon, P.; Bia, R.; Horiuchi, E.; Franklin, S. Transcriptional Regulation by Methyltransferases and Their Role in the Heart: Highlighting Novel Emerging Functionality. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H847–H865. [Google Scholar] [CrossRef]
- Qian, C.; Zhou, M.-M. SET Domain Protein Lysine Methyltransferases: Structure, Specificity and Catalysis. Cell. Mol. Life Sci. 2006, 63, 2755–2763. [Google Scholar] [CrossRef]
- Fagan, R.J.; Dingwall, A.K. COMPASS Ascending: Emerging Clues Regarding the Roles of MLL3/KMT2C and MLL2/KMT2D Proteins in Cancer. Cancer Lett. 2019, 458, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Herz, H.-M.; Hu, D.; Shilatifard, A. Enhancer Malfunction in Cancer. Mol. Cell 2014, 53, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.; Ferdman, D.; Copel, J.; Scheinost, D.; Shabanova, V.; Brueckner, M.; Khokha, M.K.; Ment, L.R. De Novo Damaging Variants Associated with Congenital Heart Diseases Contribute to the Connectome. Sci. Rep. 2020, 10, 7046. [Google Scholar] [CrossRef] [PubMed]
- Jinxiu, L.; Shuimei, L.; Ming, X.; Jonathan, L.C.; Xiangju, L.; Wenyuan, D. Wiedemann-Steiner Syndrome with a de Novo Mutation in KMT2A: A Case Report. Medicine 2020, 99, e19813. [Google Scholar] [CrossRef] [PubMed]
- Borland, S.; Tenin, G.; Williams, S.; Monaghan, R.; Baxter, M.; Ray, D.; Abraham, S.; Keavney, B. BS9 KMT2C—A Tetralogy of Fallot Candidate Gene. Heart 2019, 105, A145–A146. [Google Scholar]
- Ang, S.-Y.; Uebersohn, A.; Spencer, C.I.; Huang, Y.; Lee, J.-E.; Ge, K.; Bruneau, B.G. KMT2D Regulates Specific Programs in Heart Development via Histone H3 Lysine 4 Di-Methylation. Development 2016, 143, 810–821. [Google Scholar] [CrossRef]
- Serrano, M.d.L.A.; Demarest, B.L.; Tone-Pah-Hote, T.; Tristani-Firouzi, M.; Yost, H.J. Inhibition of Notch Signaling Rescues Cardiovascular Development in Kabuki Syndrome. PLoS Biol. 2019, 17, e3000087. [Google Scholar] [CrossRef]
- Schwenty-Lara, J.; Nürnberger, A.; Borchers, A. Loss of Function of Kmt2d, a Gene Mutated in Kabuki Syndrome, Affects Heart Development in Xenopus Laevis. Dev. Dyn. 2019, 248, 465–476. [Google Scholar] [CrossRef]
- Van Laarhoven, P.M.; Neitzel, L.R.; Quintana, A.M.; Geiger, E.A.; Zackai, E.H.; Clouthier, D.E.; Artinger, K.B.; Ming, J.E.; Shaikh, T.H. Kabuki Syndrome Genes KMT2D and KDM6A: Functional Analyses Demonstrate Critical Roles in Craniofacial, Heart and Brain Development. Hum. Mol. Genet. 2015, 24, 4443–4453. [Google Scholar] [CrossRef]
- Bokinni, Y. Kabuki Syndrome Revisited. J. Hum. Genet. 2012, 57, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Digilio, M.C.; Gnazzo, M.; Lepri, F.; Dentici, M.L.; Pisaneschi, E.; Baban, A.; Passarelli, C.; Capolino, R.; Angioni, A.; Novelli, A.; et al. Congenital Heart Defects in Molecularly Proven Kabuki Syndrome Patients. Am. J. Med. Genet. A 2017, 173, 2912–2922. [Google Scholar] [CrossRef] [PubMed]
- Souidi, A.; Jagla, K. Drosophila Heart as a Model for Cardiac Development and Diseases. Cells 2021, 10, 3078. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-Y.; Fu, Y.; Nettleton, M.; Richman, A.; Han, Z. High Throughput in Vivo Functional Validation of Candidate Congenital Heart Disease Genes in Drosophila. eLife 2017, 6, e22617. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Fu, Y.; Lee, H.; Zhao, Y.; Yang, W.; van de Leemput, J.; Han, Z. Single-Cell Profiling of the Developing Embryonic Heart in Drosophila. Development 2023, 150, dev201936. [Google Scholar] [CrossRef] [PubMed]
- modENCODE Consortium; Roy, S.; Ernst, J.; Kharchenko, P.V.; Kheradpour, P.; Negre, N.; Eaton, M.L.; Landolin, J.M.; Bristow, C.A.; Ma, L.; et al. Identification of Functional Elements and Regulatory Circuits by Drosophila ModENCODE. Science 2010, 330, 1787–1797. [Google Scholar] [PubMed]
- Graveley, B.R.; Brooks, A.N.; Carlson, J.W.; Duff, M.O.; Landolin, J.M.; Yang, L.; Artieri, C.G.; van Baren, M.J.; Boley, N.; Booth, B.W.; et al. The Developmental Transcriptome of Drosophila Melanogaster. Nature 2011, 471, 473–479. [Google Scholar] [CrossRef]
- Vaughan, L.; Marley, R.; Miellet, S.; Hartley, P.S. The Impact of SPARC on Age-Related Cardiac Dysfunction and Fibrosis in Drosophila. Exp. Gerontol. 2018, 109, 59–66. [Google Scholar] [CrossRef]
- Hughes, C.J.R.; Turner, S.; Andrews, R.M.; Vitkin, A.; Jacobs, J.R. Matrix Metalloproteinases Regulate ECM Accumulation but Not Larval Heart Growth in Drosophila Melanogaster. J. Mol. Cell. Cardiol. 2020, 140, 42–55. [Google Scholar] [CrossRef]
- Gupta, H.P.; Fatima, M.-U.; Pandey, R.; Ravi Ram, K. Adult Exposure of Atrazine Alone or in Combination with Carbohydrate Diet Hastens the Onset/Progression of Type 2 Diabetes in Drosophila. Life Sci. 2023, 316, 121370. [Google Scholar] [CrossRef]
- Ren, J.; Zeng, Q.; Wu, H.; Liu, X.; Guida, M.C.; Huang, W.; Zhai, Y.; Li, J.; Ocorr, K.; Bodmer, R.; et al. Deacetylase-Dependent and -Independent Role of HDAC3 in Cardiomyopathy. J. Cell. Physiol. 2023, 238, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Tristani-Firouzi, M.; Jensen, J.L.; Donaldson, M.R.; Sansone, V.; Meola, G.; Hahn, A.; Bendahhou, S.; Kwiecinski, H.; Fidzianska, A.; Plaster, N.; et al. Functional and Clinical Characterization of KCNJ2 Mutations Associated with LQT7 (Andersen Syndrome). J. Clin. Investig. 2002, 110, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Ocorr, K.; Reeves, N.L.; Wessells, R.J.; Fink, M.; Chen, H.-S.V.; Akasaka, T.; Yasuda, S.; Metzger, J.M.; Giles, W.; Posakony, J.W.; et al. KCNQ Potassium Channel Mutations Cause Cardiac Arrhythmias in Drosophila That Mimic the Effects of Aging. Proc. Natl. Acad. Sci. USA 2007, 104, 3943–3948. [Google Scholar] [CrossRef]
- Kurtz, P.; Jones, A.E.; Tiwari, B.; Link, N.; Wylie, A.; Tracy, C.; Krämer, H.; Abrams, J.M. Drosophila P53 Directs Nonapoptotic Programs in Postmitotic Tissue. Mol. Biol. Cell 2019, 30, 1339–1351. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Tate, C.M.; You, J.-S.; Skalnik, D.G. Identification and Characterization of the Human Set1B Histone H3-Lys4 Methyltransferase Complex. J. Biol. Chem. 2007, 282, 13419–13428. [Google Scholar] [CrossRef]
- Shinsky, S.A.; Monteith, K.E.; Viggiano, S.; Cosgrove, M.S. Biochemical Reconstitution and Phylogenetic Comparison of Human SET1 Family Core Complexes Involved in Histone Methylation. J. Biol. Chem. 2015, 290, 6361–6375. [Google Scholar] [CrossRef]
- Higgs, M.R.; Sato, K.; Reynolds, J.J.; Begum, S.; Bayley, R.; Goula, A.; Vernet, A.; Paquin, K.L.; Skalnik, D.G.; Kobayashi, W.; et al. Histone Methylation by SETD1A Protects Nascent DNA through the Nucleosome Chaperone Activity of FANCD2. Mol. Cell 2018, 71, 25–41.e6. [Google Scholar] [CrossRef]
- Jones, W.D.; Dafou, D.; McEntagart, M.; Woollard, W.J.; Elmslie, F.V.; Holder-Espinasse, M.; Irving, M.; Saggar, A.K.; Smithson, S.; Trembath, R.C.; et al. De Novo Mutations in MLL Cause Wiedemann-Steiner Syndrome. Am. J. Hum. Genet. 2012, 91, 358–364. [Google Scholar] [CrossRef]
- Miyake, N.; Tsurusaki, Y.; Koshimizu, E.; Okamoto, N.; Kosho, T.; Brown, N.J.; Tan, T.Y.; Yap, P.J.J.; Suzumura, H.; Tanaka, T.; et al. Delineation of Clinical Features in Wiedemann-Steiner Syndrome Caused by KMT2A Mutations. Clin. Genet. 2016, 89, 115–119. [Google Scholar] [CrossRef]
- Sheppard, S.E.; Campbell, I.M.; Harr, M.H.; Gold, N.; Li, D.; Bjornsson, H.T.; Cohen, J.S.; Fahrner, J.A.; Fatemi, A.; Harris, J.R.; et al. Expanding the Genotypic and Phenotypic Spectrum in a Diverse Cohort of 104 Individuals with Wiedemann-Steiner Syndrome. Am. J. Med. Genet. A 2021, 185, 1649–1665. [Google Scholar] [CrossRef]
- Jiang, D.-S.; Yi, X.; Li, R.; Su, Y.-S.; Wang, J.; Chen, M.-L.; Liu, L.-G.; Hu, M.; Cheng, C.; Zheng, P.; et al. The Histone Methyltransferase Mixed Lineage Leukemia (MLL) 3 May Play a Potential Role on Clinical Dilated Cardiomyopathy. Mol. Med. 2017, 23, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-E.; Wang, C.; Xu, S.; Cho, Y.-W.; Wang, L.; Feng, X.; Baldridge, A.; Sartorelli, V.; Zhuang, L.; Peng, W.; et al. H3K4 Mono- and Di-Methyltransferase MLL4 Is Required for Enhancer Activation during Cell Differentiation. eLife 2013, 2, e01503. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.B.; Bigham, A.W.; Buckingham, K.J.; Hannibal, M.C.; McMillin, M.J.; Gildersleeve, H.I.; Beck, A.E.; Tabor, H.K.; Cooper, G.M.; Mefford, H.C.; et al. Exome Sequencing Identifies MLL2 Mutations as a Cause of Kabuki Syndrome. Nat. Genet. 2010, 42, 790–793. [Google Scholar] [CrossRef] [PubMed]
- Li, A.H.; Hanchard, N.A.; Furthner, D.; Fernbach, S.; Azamian, M.; Nicosia, A.; Rosenfeld, J.; Muzny, D.; D’Alessandro, L.C.A.; Morris, S.; et al. Whole Exome Sequencing in 342 Congenital Cardiac Left Sided Lesion Cases Reveals Extensive Genetic Heterogeneity and Complex Inheritance Patterns. Genome Med. 2017, 9, 95. [Google Scholar] [CrossRef]
- Cuvertino, S.; Hartill, V.; Colyer, A.; Garner, T.; Nair, N.; Al-Gazali, L.; Canham, N.; Faundes, V.; Flinter, F.; Hertecant, J.; et al. A Restricted Spectrum of Missense KMT2D Variants Cause a Multiple Malformations Disorder Distinct from Kabuki Syndrome. Genet. Med. 2020, 22, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Shi, X.; Yu, C.; Shao, X.; Zhou, H.; Li, X.; Chang, C.; Lai, K.S.; Ma, J.; Zhang, R. Identification of Novel Candidate Genes in Heterotaxy Syndrome Patients with Congenital Heart Diseases by Whole Exome Sequencing. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165906. [Google Scholar] [CrossRef]
- Sun, H.; Yi, T.; Hao, X.; Yan, H.; Wang, J.; Li, Q.; Gu, X.; Zhou, X.; Wang, S.; Wang, X.; et al. Contribution of Single-Gene Defects to Congenital Cardiac Left-Sided Lesions in the Prenatal Setting. Ultrasound Obstet. Gynecol. 2020, 56, 225–232. [Google Scholar] [CrossRef]
- Luo, S.; Chen, L.; Wei, W.; Tan, L.; Zhang, M.; Duan, Z.; Cao, J.; Zhou, Y.; Zhou, A.; He, X. Prenatal Genetic Diagnosis in Three Fetuses With Left Heart Hypoplasia (LHH) From Three Unrelated Families. Front. Cardiovasc. Med. 2021, 8, 631374. [Google Scholar] [CrossRef]
- Wan, X.; Liu, L.; Ding, X.; Zhou, P.; Yuan, X.; Zhou, Z.; Hu, P.; Zhou, H.; Li, Q.; Zhang, S.; et al. Mll2 Controls Cardiac Lineage Differentiation of Mouse Embryonic Stem Cells by Promoting H3K4me3 Deposition at Cardiac-Specific Genes. Stem Cell Rev. Rep. 2014, 10, 643–652. [Google Scholar] [CrossRef]
- Chen, L.; Hassani Nia, F.; Stauber, T. Ion Channels and Transporters in Muscle Cell Differentiation. Int. J. Mol. Sci. 2021, 22, 13615. [Google Scholar] [CrossRef]
- Han, Z.; Olson, E.N. Hand Is a Direct Target of Tinman and GATA Factors during Drosophila Cardiogenesis and Hematopoiesis. Development 2005, 132, 3525–3536. [Google Scholar] [CrossRef]
- Zhu, J.-Y.; Fu, Y.; Richman, A.; Han, Z. Validating Candidate Congenital Heart Disease Genes in Drosophila. Bio-Protocol 2017, 7, e2350. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Spletter, M.L.; Barz, C.; Yeroslaviz, A.; Zhang, X.; Lemke, S.B.; Bonnard, A.; Brunner, E.; Cardone, G.; Basler, K.; Habermann, B.H.; et al. A Transcriptomics Resource Reveals a Transcriptional Transition during Ordered Sarcomere Morphogenesis in Flight Muscle. eLife 2018, 7, e34058. [Google Scholar] [CrossRef] [PubMed]
- Yelbuz, T.M.; Choma, M.A.; Thrane, L.; Kirby, M.L.; Izatt, J.A. Optical Coherence Tomography: A New High-Resolution Imaging Technology to Study Cardiac Development in Chick Embryos. Circulation 2002, 106, 2771–2774. [Google Scholar] [CrossRef] [PubMed]
- Choma, M.A.; Izatt, S.D.; Wessells, R.J.; Bodmer, R.; Izatt, J.A. Images in Cardiovascular Medicine: In Vivo Imaging of the Adult Drosophila Melanogaster Heart with Real-Time Optical Coherence Tomography. Circulation 2006, 114, e35–e36. [Google Scholar] [CrossRef] [PubMed]
- Hoskins, R.A.; Carlson, J.W.; Wan, K.H.; Park, S.; Mendez, I.; Galle, S.E.; Booth, B.W.; Pfeiffer, B.D.; George, R.A.; Svirskas, R.; et al. The Release 6 Reference Sequence of the Drosophila Melanogaster Genome. Genome Res. 2015, 25, 445–458. [Google Scholar] [CrossRef]
- Howe, K.L.; Achuthan, P.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J.; et al. Ensembl 2021. Nucleic Acids Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Gramates, L.S.; Agapite, J.; Attrill, H.; Calvi, B.R.; Crosby, M.A.; Dos Santos, G.; Goodman, J.L.; Goutte-Gattat, D.; Jenkins, V.K.; Kaufman, T.; et al. FlyBase: A Guided Tour of Highlighted Features. Genetics 2022, 220, iyac035. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-Y.; Wang, G.; Huang, X.; Lee, H.; Lee, J.-G.; Yang, P.; van de Leemput, J.; Huang, W.; Kane, M.A.; Yang, P.; et al. SARS-CoV-2 Nsp6 Damages Drosophila Heart and Mouse Cardiomyocytes through MGA/MAX Complex-Mediated Increased Glycolysis. Commun. Biol. 2022, 5, 1039. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.-y.; Lee, H.; Huang, X.; van de Leemput, J.; Han, Z. Distinct Roles for COMPASS Core Subunits Set1, Trx, and Trr in the Epigenetic Regulation of Drosophila Heart Development. Int. J. Mol. Sci. 2023, 24, 17314. https://doi.org/10.3390/ijms242417314
Zhu J-y, Lee H, Huang X, van de Leemput J, Han Z. Distinct Roles for COMPASS Core Subunits Set1, Trx, and Trr in the Epigenetic Regulation of Drosophila Heart Development. International Journal of Molecular Sciences. 2023; 24(24):17314. https://doi.org/10.3390/ijms242417314
Chicago/Turabian StyleZhu, Jun-yi, Hangnoh Lee, Xiaohu Huang, Joyce van de Leemput, and Zhe Han. 2023. "Distinct Roles for COMPASS Core Subunits Set1, Trx, and Trr in the Epigenetic Regulation of Drosophila Heart Development" International Journal of Molecular Sciences 24, no. 24: 17314. https://doi.org/10.3390/ijms242417314