
Kiwifruit Monodehydroascorbate Reductase 3 Gene Negatively Regulates the Accumulation of Ascorbic Acid in Fruit of Transgenic Tomato Plants
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Kiwifruit MDHAR Family Members and Their Sequence Features
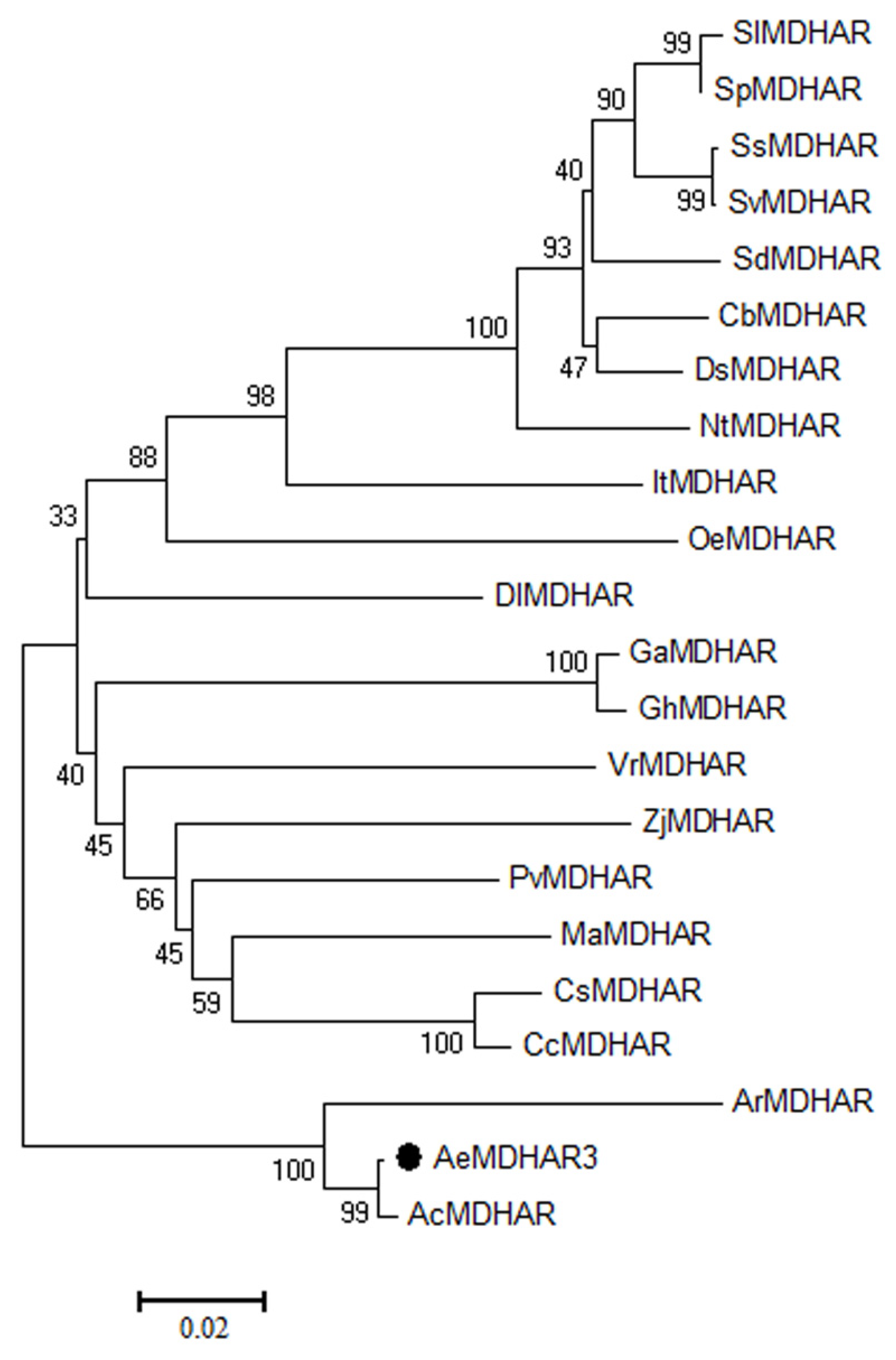
2.2. Phylogenetic Relationships of MDHAR Proteins for Kiwifruit and Other Species
2.3. Structures of AeMDHAR Genes and the Conserved Motifs of AeMDHAR Proteins
2.4. Synteny Relationships of MDHAR Genes within Kiwifruit Genome and between Genomes of Kiwifruit and Arabidopsis
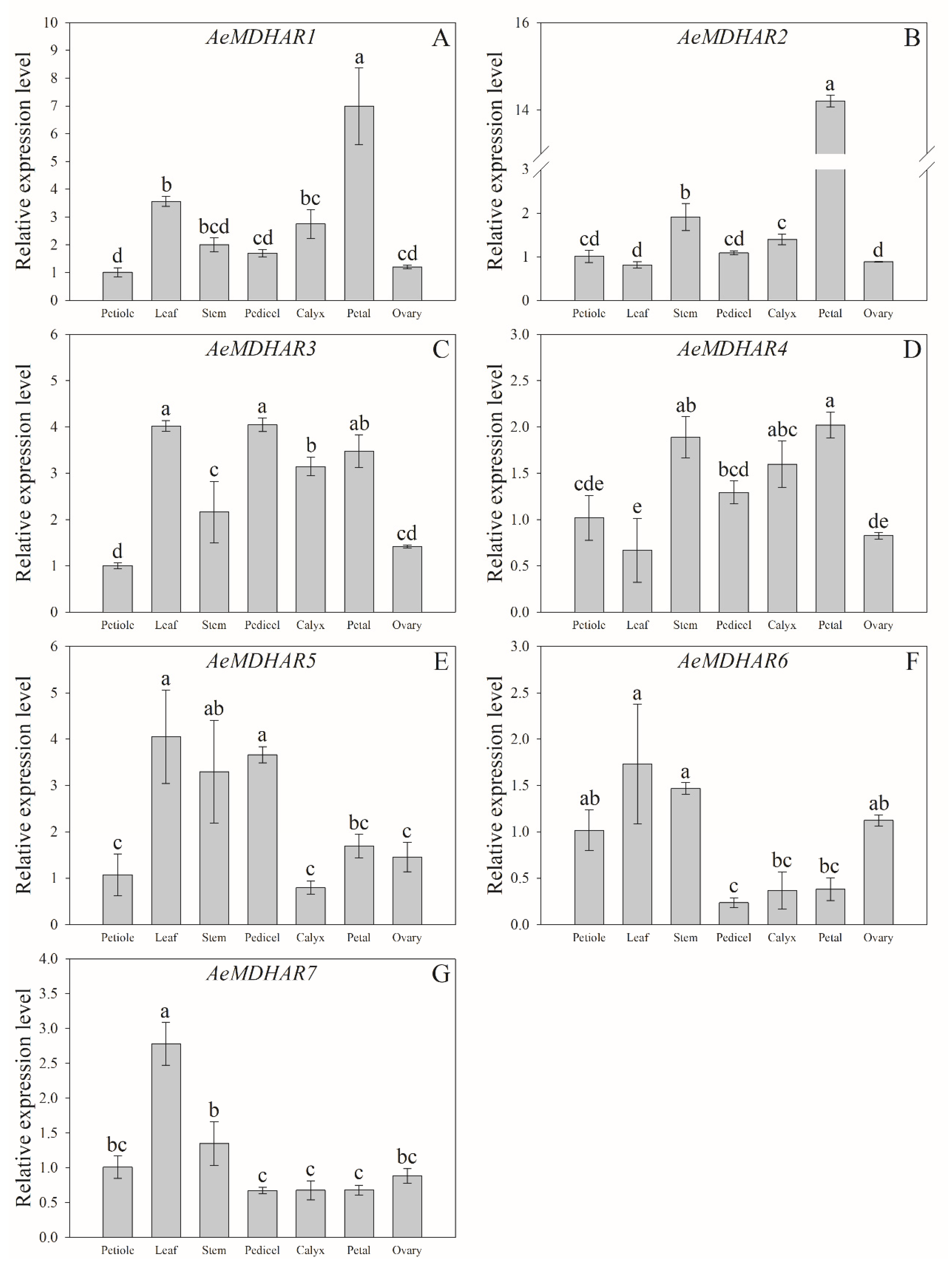
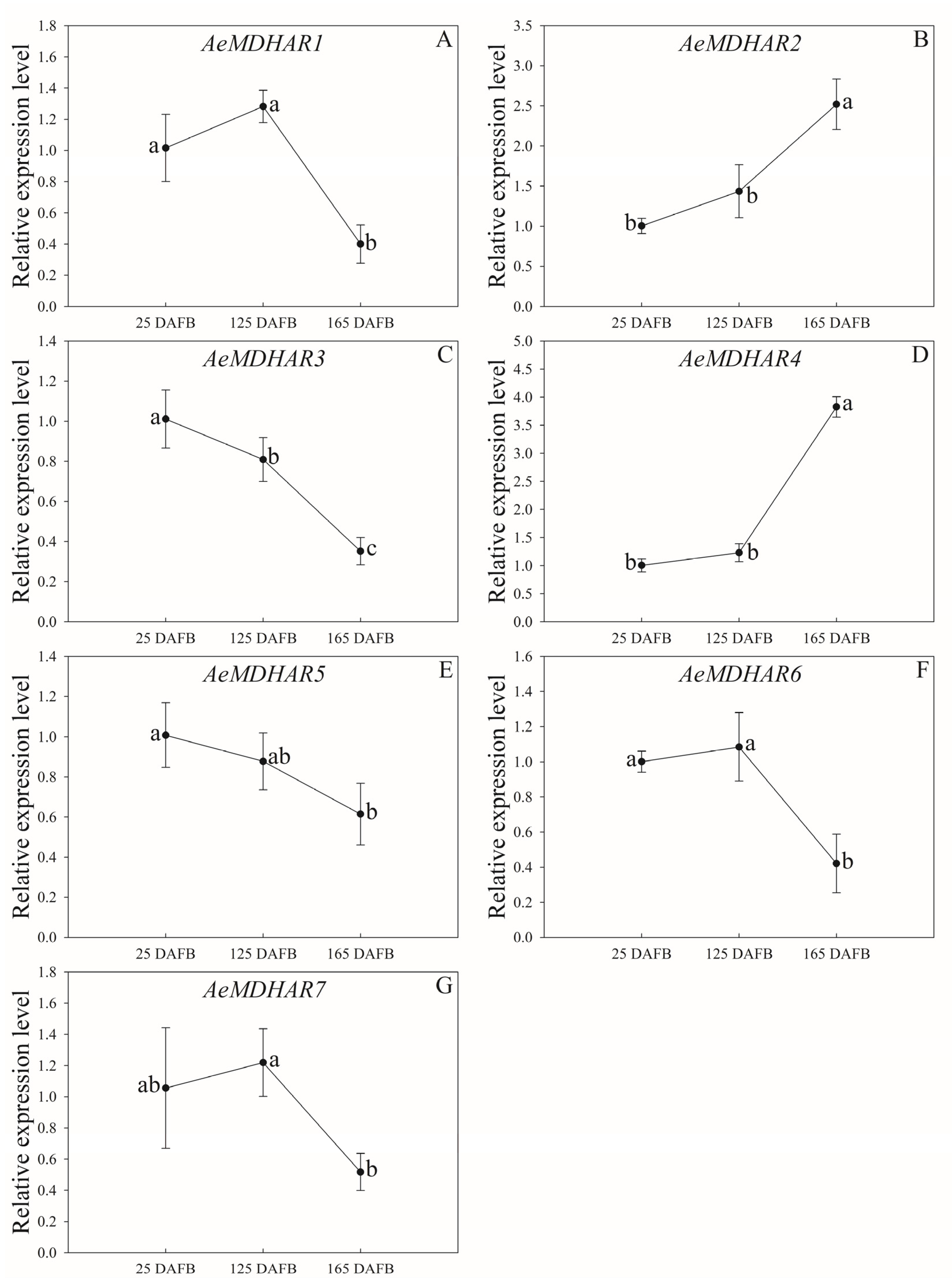
2.5. Relative Expressions of Kiwifruit MDHAR Genes in Different Tissues and in Fruits with Different Developmental Stages
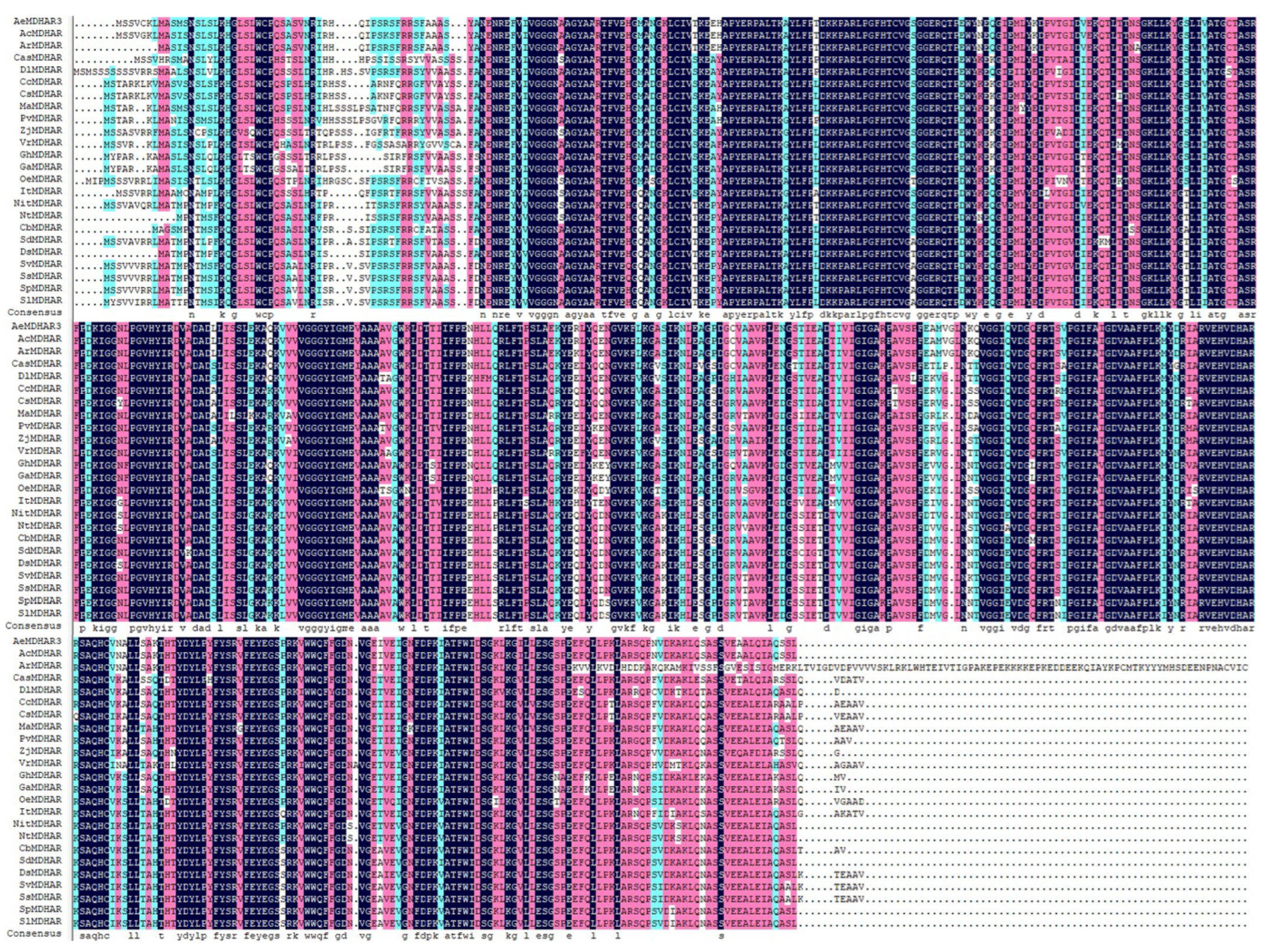
2.6. Molecular Features, Phylogenetic Relationships, and Sequence Similarities of AeMDHAR3 and Homologous Proteins from Other Species
2.7. Expression of AeMDHAR3 and Activity of MDHAR in Transgenic Tomato Lines
2.8. Contents of AsA and DHA in Transgenic Tomato Lines
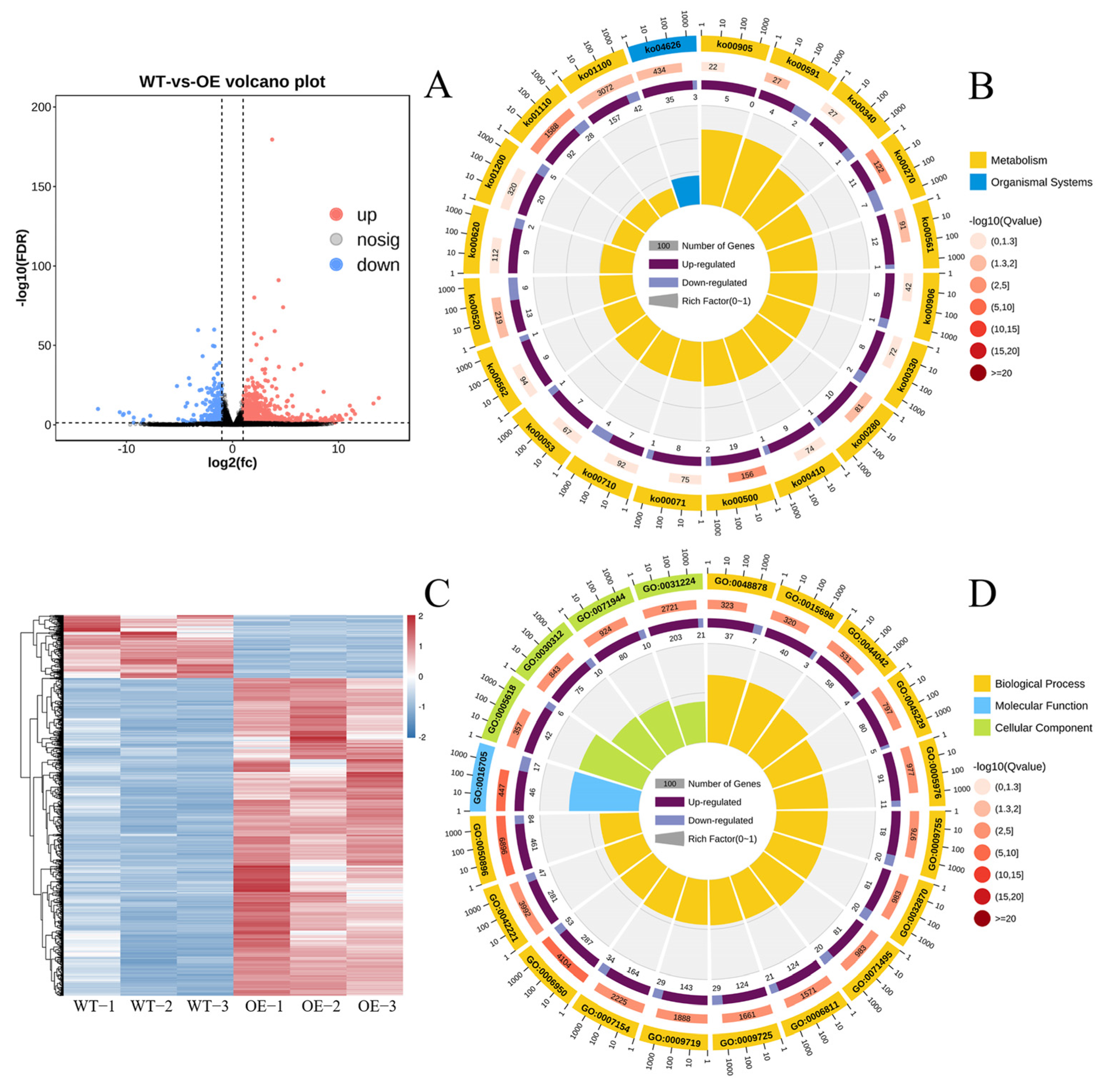
2.9. Differently Expressed Genes and Their GO Enrichment and KEGG Pathway Enrichment
2.10. Results of Expression Verification of Related Differently Expressed Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Genome-Wide Identification of MDHAR Protein Family Members in Kiwifruit (Actinidia eriantha)
4.3. Phylogenetic Analysis for MDHAR Proteins of Kiwifruit and Other Species
4.4. Analyses of Genomic Structure and Sequence Features for MDHAR Genes
4.5. Analysis of Chromosomal Location and Synteny for MDHAR Genes
4.6. Analysis of Relative Expression Levels for Genes in Kiwifruit or Tomato
4.7. Generation of Transgenic Tomato Lines Over-Expressing Kiwifruit AeMDHAR3 Gene
4.8. Analyses of Expression, Enzymatic Activity, and Contents of AsA and DHA in Transgenic Tomato Lines
4.9. Transcriptome Sequencing and Identification of Differentially Expressed Genes
4.10. Analyses of Gene Ontology Enrichment and KEGG Pathway Enrichment for the Differentially Expressed genes
4.11. Expression Verification for Differentially Expressed Genes via Quantitative Real-Time PCR Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smirnoff, N. Vitamin C: The metabolism and functions of ascorbic acid in plants. Adv. Plant Pathol. 2011, 59, 107–177. [Google Scholar]
- Mellidou, I.; Kanellis, A.K. Genetic control of ascorbic acid biosynthesis and recycling in horticultural crops. Front. Chem. 2017, 5, 50. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, D.S. Overview of species needing dietary vitamin C. J. Zoo Anim. Med. 1980, 11, 88–91. [Google Scholar] [CrossRef]
- Jia, D.; Xu, Z.; Chen, L.; Huang, Q.; Huang, C.; Tao, J.; Qu, X.; Xu, X. Analysis of organic acid metabolism reveals citric acid and malic acid play major roles in determining acid quality during the development of kiwifruit (Actinidia eriantha). J. Sci. Food Agric. 2023, 103, 6055–6069. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xie, J.; Wang, F.; Zhong, J.; Liu, Y.; Li, G.; Peng, S. Comparison of ascorbate metabolism in fruits of two citrus species with obvious difference in ascorbate content in pulp. J. Plant Physiol. 2011, 168, 2196–2205. [Google Scholar] [CrossRef] [PubMed]
- Gallie, D.R. The role of L-ascorbic acid recycling in responding to environmental stress and in promoting plant growth. J. Exp. Bot. 2013, 64, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Agius, F.; González-Lamothe, R.; Caballero, J.L.; MuñozBlanco, J.; Botella, M.A.; Valpuesta, V. Engineering increased vitamin C levels in plants by overexpression of a D-galacturonic acid reductase. Nat. Biotechnol. 2003, 21, 177–181. [Google Scholar] [CrossRef]
- Wolucka, B.A.; van Montagu, M. GDP-mannose 30′,50′-epimerase forms GDP-L-gulose, a putative intermediate for the de novo biosynthesis of vitamin C in plants. J. Biol. Chem. 2003, 278, 47483–47490. [Google Scholar] [CrossRef]
- Lorence, A.; Chevone, B.I.; Mendes, P.; Nessler, C.L. myo-inositol oxygenase offers a possible entry point into plant ascorbate biosynthesis. Plant Physiol. 2004, 134, 1200–1205. [Google Scholar] [CrossRef]
- Chen, Z.; Young, T.E.; Ling, J.; Chang, S.C.; Gallie, D.R. Increasing vitamin C content of plants through enhanced ascorbate recycling. Proc. Natl. Acad. Sci. USA 2003, 100, 3525–3530. [Google Scholar] [CrossRef]
- Wang, Z.; Xiao, Y.; Chen, W.; Tang, K.; Zhang, L. Increased vitamin C content accompanied by an enhanced recycling pathway confers oxidative stress tolerance in Arabidopsis. J. Integr. Plant Biol. 2010, 52, 400–409. [Google Scholar] [CrossRef]
- Haroldsen, V.M.; Chi-Ham, C.L.; Kulkarni, S.; Lorence, A.; Bennett, A.B. Constitutively expressed DHAR and MDHAR influence fruit, but not foliar ascorbate levels in tomato. Plant Physiol. Biochem. 2011, 49, 1244–1249. [Google Scholar] [CrossRef] [PubMed]
- Bulley, S.M.; Rassam, M.; Hoser, D.; Otto, W.; Schünemann, N.; Wright, M.; MacRae, E.; Gleave, A.; Laing, W. Gene expression studies in kiwifruit and gene over-expression in Arabidopsis indicates that GDP-L-galactose guanyltransferase is a major control point of vitamin C biosynthesis. J. Exp. Bot. 2009, 60, 765–778. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhong, Y.; Zheng, J.; Ali, M.; Liu, G.; Zheng, X. L-ascorbic acid metabolism in an ascorbate-rich kiwifruit (Actinidia. Eriantha Benth.) cv. ‘White’ during postharvest. Plant Physiol. Biochem. 2018, 124, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, Y.; Liao, G.; Xu, X.; Jia, D.; Zhong, M.; Wang, H.; Ye, B. Transcriptome and metabolome reveal AsA regulatory network between metabolites and genes after fruit shading by bagging in kiwifruit (Actinidia eriantha). Sci. Hortic. 2022, 302, 111184. [Google Scholar] [CrossRef]
- Liao, G.; Xu, Q.; Allan, A.C.; Xu, X. L-Ascorbic acid metabolism and regulation in fruit crops. Plant Physiol. 2023, 192, 1684–1695. [Google Scholar] [CrossRef] [PubMed]
- Alós, E.; Rodrigo, M.J.; Zacarías, L. Differential transcriptional regulation of L-ascorbic acid content in peel and pulp of citrus fruits during development and maturation. Planta 2014, 239, 1113–1128. [Google Scholar] [CrossRef]
- Mellidou, I.; Keulemans, J.; Kanellis, A.K.; Davey, M.W. Regulation of fruit ascorbic acid concentrations during ripening in high and low vitamin C tomato cultivars. BMC Plant Biol. 2012, 12, 239. [Google Scholar] [CrossRef]
- Huang, H.; Gong, J.; Wang, S.; He, Z.; Zhang, Z.; Li, J. Genetic diversity in the genus Acitinidia. Biodivers. Sci. 2000, 8, 1–12. [Google Scholar]
- Bulley, S.; Laing, W. The regulation of ascorbate biosynthesis. Curr. Opin. Plant Biol. 2016, 33, 15–22. [Google Scholar] [CrossRef]
- Sano, S.; Tao, S.; Endo, Y.; Inaba, T.; Hossain, M.A.; Miyake, C.; Matsuo, M.; Aoki, H.; Asada, K.; Saito, K. Purification and cDNA cloning of chloroplastic monodehydroascorbate reductase from spinach. Biosci. Biotechnol. Biochem. 2005, 69, 762–772. [Google Scholar] [CrossRef]
- Murthy, S.S.; Zilinskas, B.A. Molecular cloning and characterization of a cDNA encoding pea monodehydroascorbate reductase. J. Biol. Chem. 1994, 269, 31129–31133. [Google Scholar] [CrossRef]
- Eastmond, P.J. MONODEHYROASCORBATE REDUCTASE4 is required for seed storage oil hydrolysis and postgerminative growth in Arabidopsis. Plant Cell 2007, 19, 1376–1387. [Google Scholar] [CrossRef]
- Jiang, Z.; Huang, Q.; Jia, D.; Zhong, M.; Tao, J.; Liao, G.; Huang, C.; Xu, X. Characterization of organic acid metabolism and expression of related genes during fruit development of Actinidia eriantha ‘Ganmi 6’. Plants 2020, 9, 332. [Google Scholar] [CrossRef]
- Li, F.; Wu, Q.; Sun, Y.; Wang, L.; Yang, X.; Meng, Q. Overexpression of chloroplastic monodehydroascorbate reductase enhanced tolerance to temperature and methyl viologen-mediated oxidative stresses. Physiol. Plantarum. 2010, 139, 421–434. [Google Scholar] [CrossRef]
- Eltayeb, A.E.; Kawano, N.; Badawi, G.H.; Kaminaka, H.; Sanekata, T.; Shibahara, T.; Inanaga, S.; Tanaka, K. Overexpression of monodehydroascorbate reductase in transgenic tobacco confers enhanced tolerance to ozone, salt and polyethylene glycol stresses. Planta 2007, 225, 1255–1264. [Google Scholar] [CrossRef]
- Ioannidi, E.; Kalamaki, M.S.; Engineer, C.; Pateraki, I.; Alexandrou, D.; Mellidou, I.; Giovannonni, J.; Kanellis, A.K. Expression profiling of ascorbic acid-related genes during tomato fruit development and ripening and in response to stress conditions. J. Exp. Bot. 2009, 60, 663–678. [Google Scholar] [CrossRef]
- Gest, N.; Garchery, C.; Gautier, H.; Jiménez, A.; Stevens, R. Light-dependent regulation of ascorbate in tomato by a monodehydroascorbate reductase localized in peroxisomes and the cytosol. Plant Biotechnol. J. 2013, 11, 344–354. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef]
- Noshi, M.; Yamada, H.; Hatanaka, R.; Tanabe, N.; Tamoi, M.; Shigeoka, S. Arabidopsis dehydroascorbate reductase 1 and 2 modulate redox states of ascorbate-glutathione cycle in the cytosol in response to photooxidative stress. Biosci. Biotechnol. Biochem. 2016, 81, 523–533. [Google Scholar] [CrossRef]
- Truffault, V.; Gest, N.; Garchery, C.; Florian, A.; Fernie, A.R.; Gautier, H.; Stevens, R.G. Reduction of MDHAR activity in cherry tomato suppresses growth and yield and MDHAR activity is correlated with sugar levels under high light. Plant Cell Environ. 2016, 39, 1279–1292. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Jiang, Z.; Fu, H.; Chen, L.; Liao, G.; He, Y.; Huang, C.; Xu, X. Genome-wide identification and comprehensive analysis of NAC family genes involved in fruit development in kiwifruit (Actinidia). BMC Plant Biol. 2021, 21, 44. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Li, W.; Han, G.; Yang, Y.; Ma, Y.; Li, H.; Zhang, Z. Development of a seedling clone with high regeneration capacity and susceptibility to Agrobacterium in apple. Sci. Hortic. 2013, 164, 202–208. [Google Scholar] [CrossRef]
- Chen, Z.; Gallie, D.R. Dehydroascorbate reductase affects leaf growth, development, and function. Plant Physiol. 2006, 142, 775–787. [Google Scholar] [CrossRef]
- Mishra, N.P.; Mishra, R.K.; Singhal, G.S. Changes in the activities of anti-oxidant enzymes during exposure of intact wheat leaves to strong visible light at different temperatures in the presence of protein synthesis inhibitors. Plant Physiol. 1993, 102, 903–910. [Google Scholar] [CrossRef]
- Cao, J.; Jiang, W.; Zhao, Y. Experiment Guidance of Postharvest Physiology and Biochemistry of Fruits and Vegetables; China Light Industry Press: Beijing, China, 2007. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Jia, D.; Yi, S.; Huang, Q.; Liu, Q.; He, Y.; Liao, G.; Huang, C.; Xu, X. Comprehensive analysis of a red-peel kiwi berry mutant reveals key genes are responsible for anthocyanin biosynthesis in fruit. Sci. Hortic. 2023, 309, 111682. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Genome ID | Chromosomal Location (Direction) | CDS Length (bp) | Amino Acid Number | Molecular Weight (Da) | Isoelectric Point |
|---|---|---|---|---|---|---|
| AeMDHAR1 | DTZ79_05g12560 | Chr05: 23539803−23546779 (+) | 1410 | 469 | 50,584.19 | 8.07 |
| AeMDHAR2 | DTZ79_12g06930 | Chr12: 9554720−9560779 (−) | 1395 | 464 | 51,413.14 | 7.09 |
| AeMDHAR3 | DTZ79_15g00570 | Chr15: 590471−598741 (−) | 1440 | 479 | 51,964.37 | 8.49 |
| AeMDHAR4 | DTZ79_20g04830 | Chr20: 6405439−6411609 (−) | 1506 | 501 | 54,574.62 | 5.69 |
| AeMDHAR5 | DTZ79_25g06420 | Chr25: 15667066−15679146 (−) | 1506 | 501 | 54,849.56 | 7.16 |
| AeMDHAR6 | DTZ79_27g01630 | Chr27: 1604069−1610795 (+) | 1407 | 468 | 50,616.09 | 7.6 |
| AeMDHAR7 | DTZ79_27g11730 | Chr27: 24128673−24134913 (−) | 1383 | 460 | 50,130.73 | 8.3 |
| Sample | Raw Reads | Clean Reads | Total Bases (Gb) | Filtered Bases (Gb) | Q20 (Percentage of Base with Qphred Value ≥ 20) | Q30 (Percentage of Base with Qphred Value ≥ p30) | GC (Percentage of GC Content) |
|---|---|---|---|---|---|---|---|
| WT−1 | 46,335,034 | 46,121,526 | 6.47 | 6.42 | 97.01 | 91.75 | 42.23 |
| WT−2 | 50,712,724 | 50,513,766 | 7.08 | 7.03 | 97.8 | 93.41 | 42.21 |
| WT−3 | 91,821,512 | 91,422,210 | 12.83 | 12.73 | 96.86 | 91.44 | 42.22 |
| OE−1−1 | 49,241,644 | 49,011,976 | 6.88 | 6.82 | 96.94 | 91.62 | 42.3 |
| OE−1−2 | 46,022,396 | 45,816,832 | 6.43 | 6.37 | 96.94 | 91.61 | 42.17 |
| OE−1−3 | 41,615,986 | 41,434,366 | 5.81 | 5.76 | 97.07 | 91.87 | 42.1 |
| Genome ID | Gene Name | Symbol | Description | KEGG Pathway |
|---|---|---|---|---|
| Solyc07g062140.3 | SlTPS1 | TPS1 | Trehalose-phosphate synthase 1 | ko00500/starch and sucrose metabolism |
| Solyc08g079820.3 | SlNUDT14 | NUDT14 | Predicted: nudix hydrolase 14, chloroplastic | ko00500/starch and sucrose metabolism |
| Solyc09g010080.3 | SlINV1 | INV1 | Beta-fructofuranosidase | ko00500/starch and sucrose metabolism |
| Solyc02g080300.3 | SlBGLU47 | BGLU47 | Predicted: beta-glucosidase 18 | ko00500/starch and sucrose metabolism |
| Solyc04g053120.3 | SlDPEP | DPEP | Predicted: 4-alpha-glucanotransferase, chloroplastic/amyloplastic | ko00500/starch and sucrose metabolism |
| Solyc03g121070.3 | SlHXK1 | HXK1 | Hexokinase | ko00500/starch and sucrose metabolism |
| Solyc07g056140.3 | SlAGPS1 | AGPS1 | Glucose-1-phosphate adenylyltransferase small subunit, chloroplastic | ko00500/starch and sucrose metabolism |
| Solyc04g078900.3 | SlCYP707A4 | CYP707A4 | ABA 8’-hydroxylase | ko00906/carotenoid biosynthesis |
| Solyc01g109930.4 | SlHISN7 | HISN7 | Predicted: bifunctional phosphatase IMPL2, chloroplastic | ko00053/ascorbate and aldarate metabolism |
| Solyc05g005700.4 | SlALDH2B7 | ALDH2B7 | Aldehyde dehydrogenase family 2 member B7d | ko00053/ascorbate and aldarate metabolism |
| Solyc02g084640.4 | SlALDH3F1 | ALDH3F1 | Predicted: aldehyde dehydrogenase family 3 member F1 | ko00053/ascorbate and aldarate metabolism |
| Solyc01g111510.3 | SlAPX3 | APX3 | L-ascorbate peroxidase 3, peroxisomal | ko00053/ascorbate and aldarate metabolism |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, D.; Gao, H.; He, Y.; Liao, G.; Lin, L.; Huang, C.; Xu, X. Kiwifruit Monodehydroascorbate Reductase 3 Gene Negatively Regulates the Accumulation of Ascorbic Acid in Fruit of Transgenic Tomato Plants. Int. J. Mol. Sci. 2023, 24, 17182. https://doi.org/10.3390/ijms242417182
Jia D, Gao H, He Y, Liao G, Lin L, Huang C, Xu X. Kiwifruit Monodehydroascorbate Reductase 3 Gene Negatively Regulates the Accumulation of Ascorbic Acid in Fruit of Transgenic Tomato Plants. International Journal of Molecular Sciences. 2023; 24(24):17182. https://doi.org/10.3390/ijms242417182
Chicago/Turabian StyleJia, Dongfeng, Huan Gao, Yanqun He, Guanglian Liao, Liting Lin, Chunhui Huang, and Xiaobiao Xu. 2023. "Kiwifruit Monodehydroascorbate Reductase 3 Gene Negatively Regulates the Accumulation of Ascorbic Acid in Fruit of Transgenic Tomato Plants" International Journal of Molecular Sciences 24, no. 24: 17182. https://doi.org/10.3390/ijms242417182