
Glucocorticoid Nanoparticles Show Full Therapeutic Efficacy in a Mouse Model of Acute Lung Injury and Concomitantly Reduce Adverse Effects
 , , and
, , and 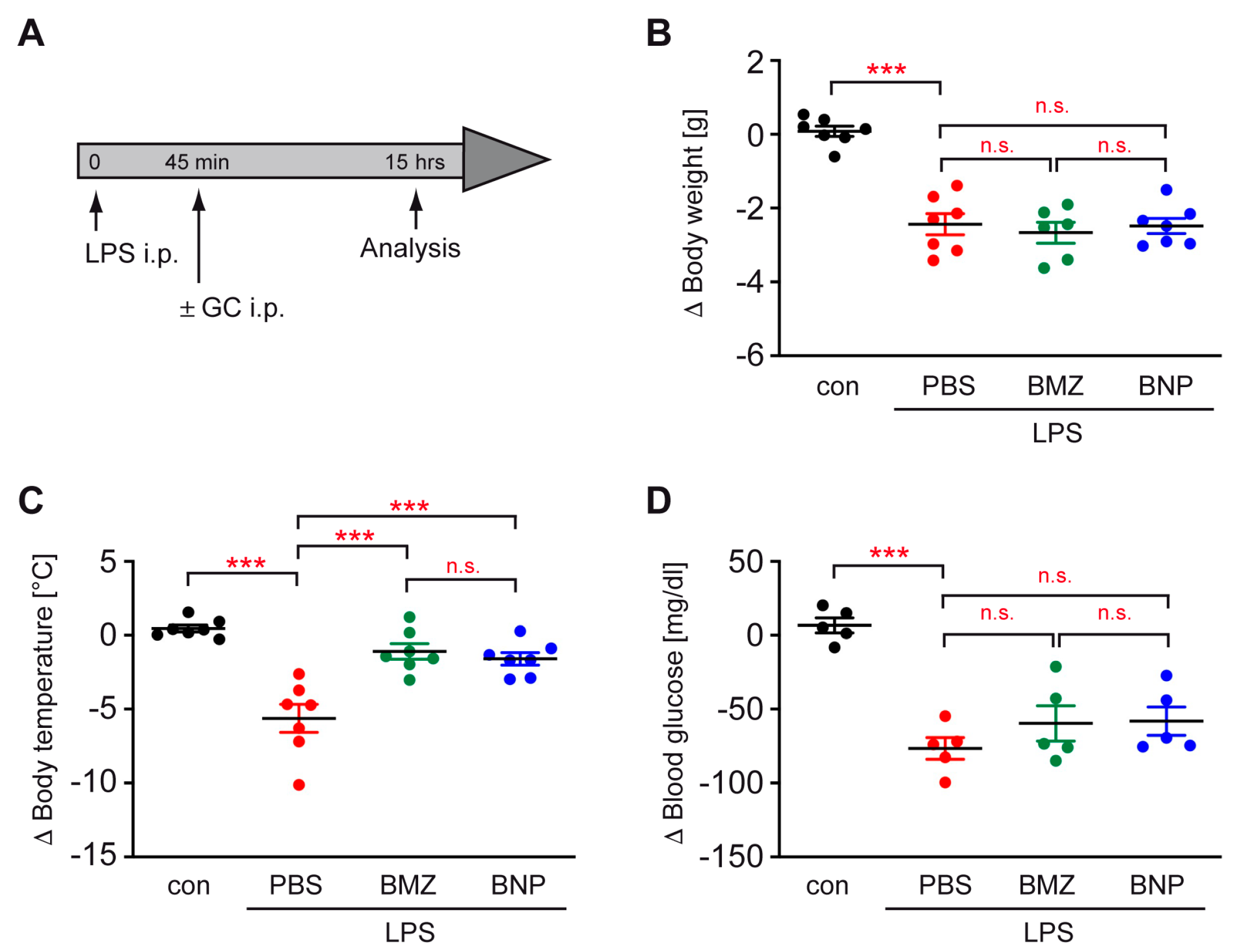{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Synthesis, Characterization and Tissue Distribution of BNPs
2.2. GC Treatment Prevents Hypothermia in a Mouse Model of Sepsis
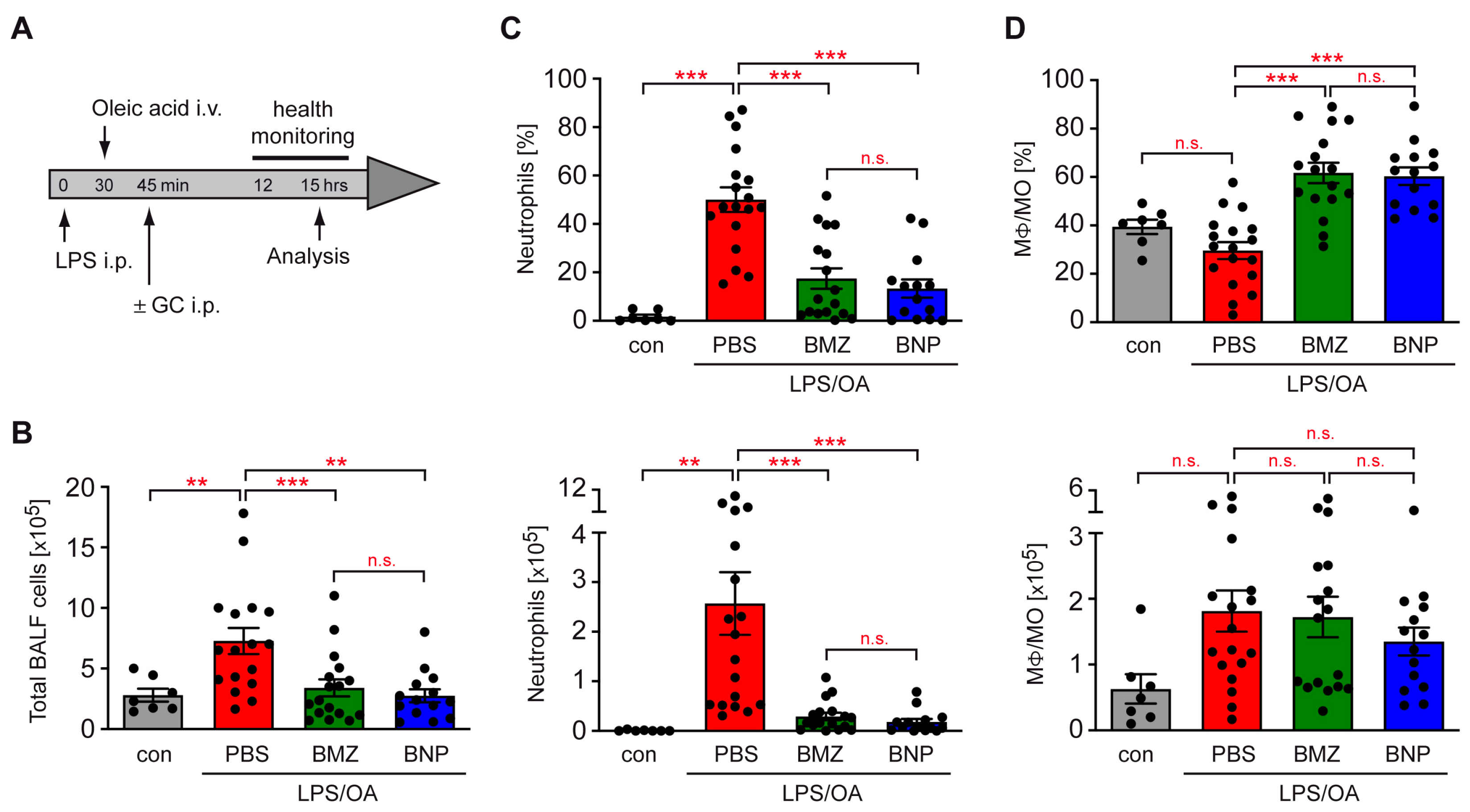
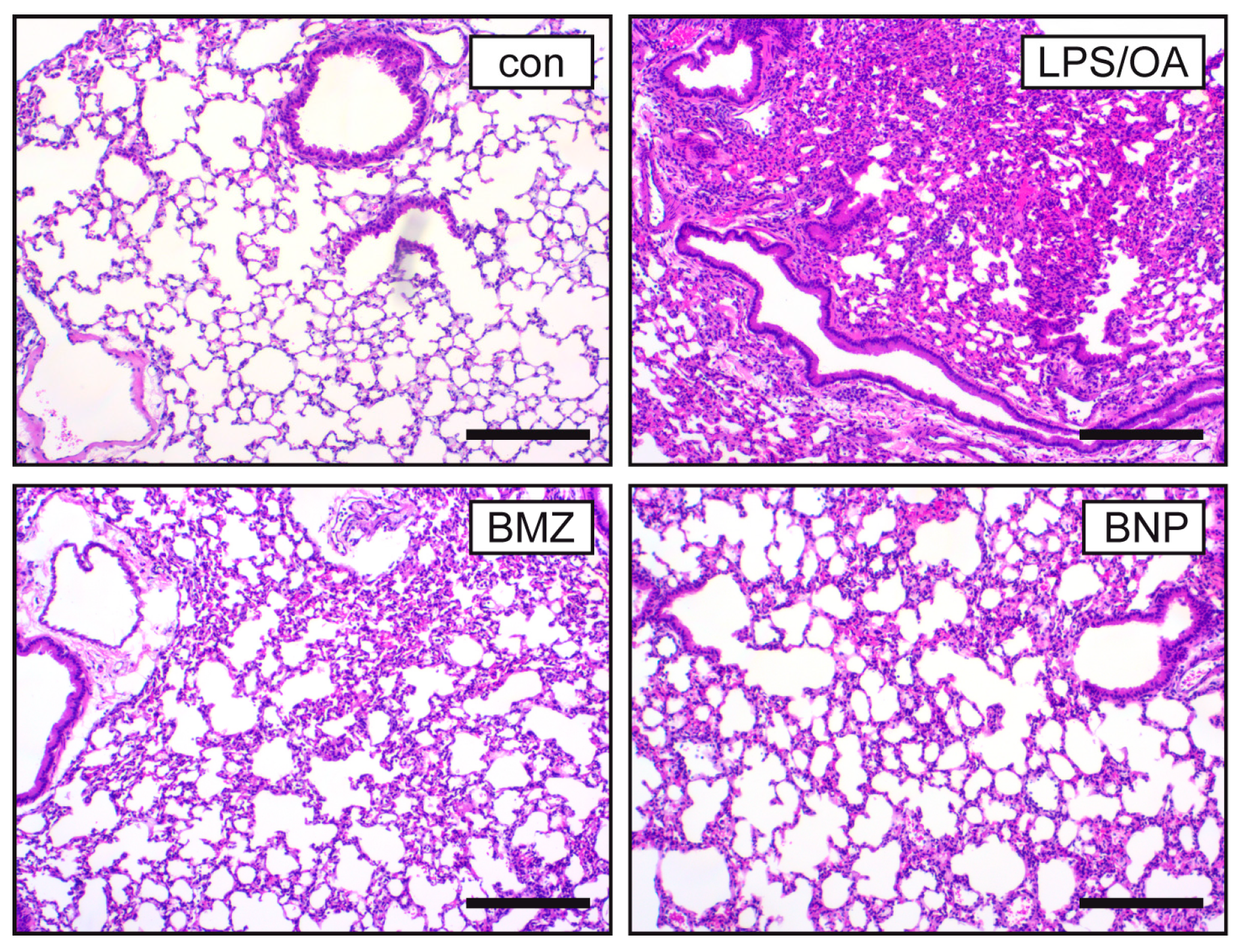
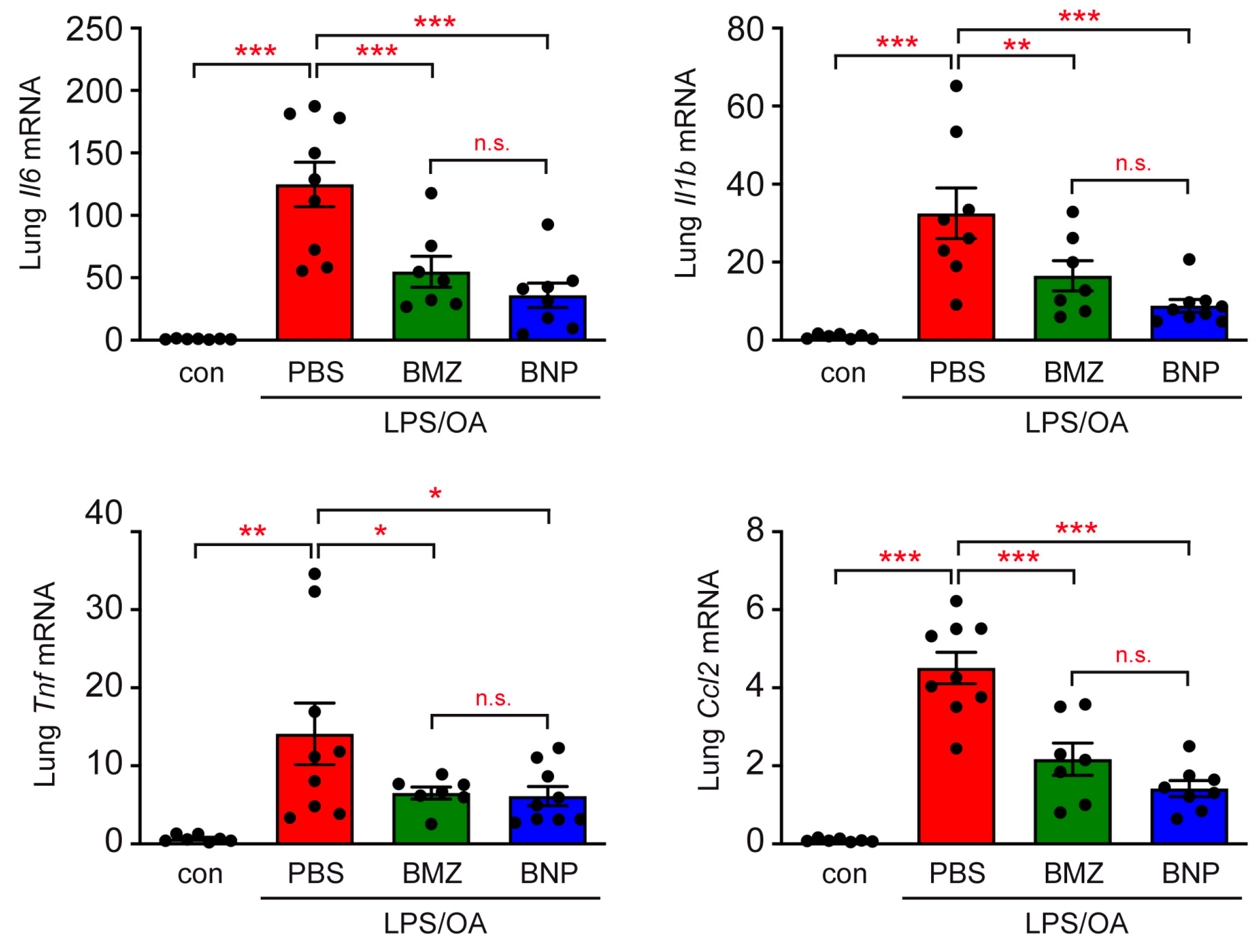
2.3. Airway Inflammation in a Mouse Model of ALI Is Repressed by GCs
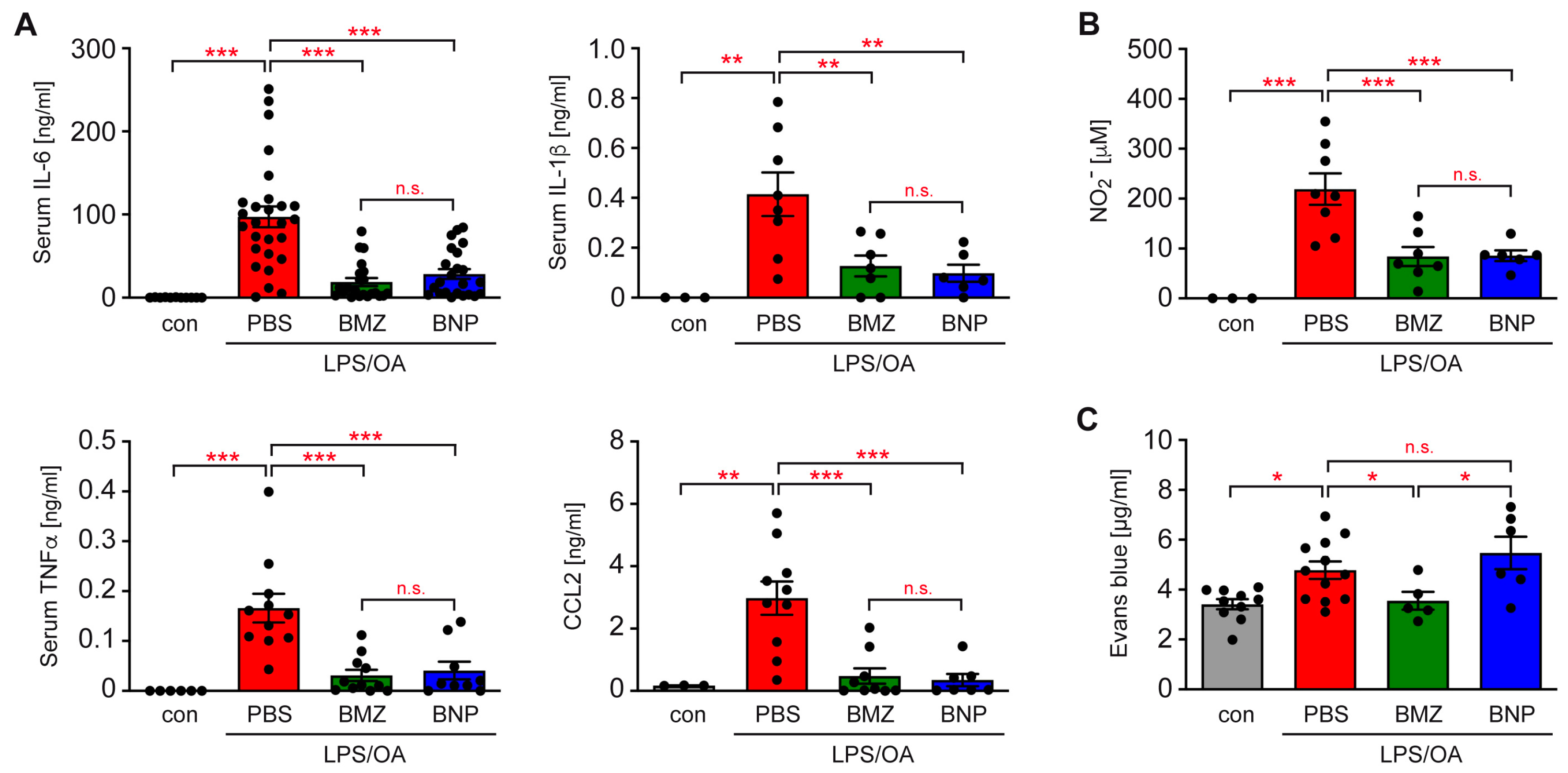
2.4. GCs Improve Systemic Inflammation in a Mouse Model of ALI
2.5. Differential Impact of GCs on the Integrity of the Alveolar–Capillary Barrier
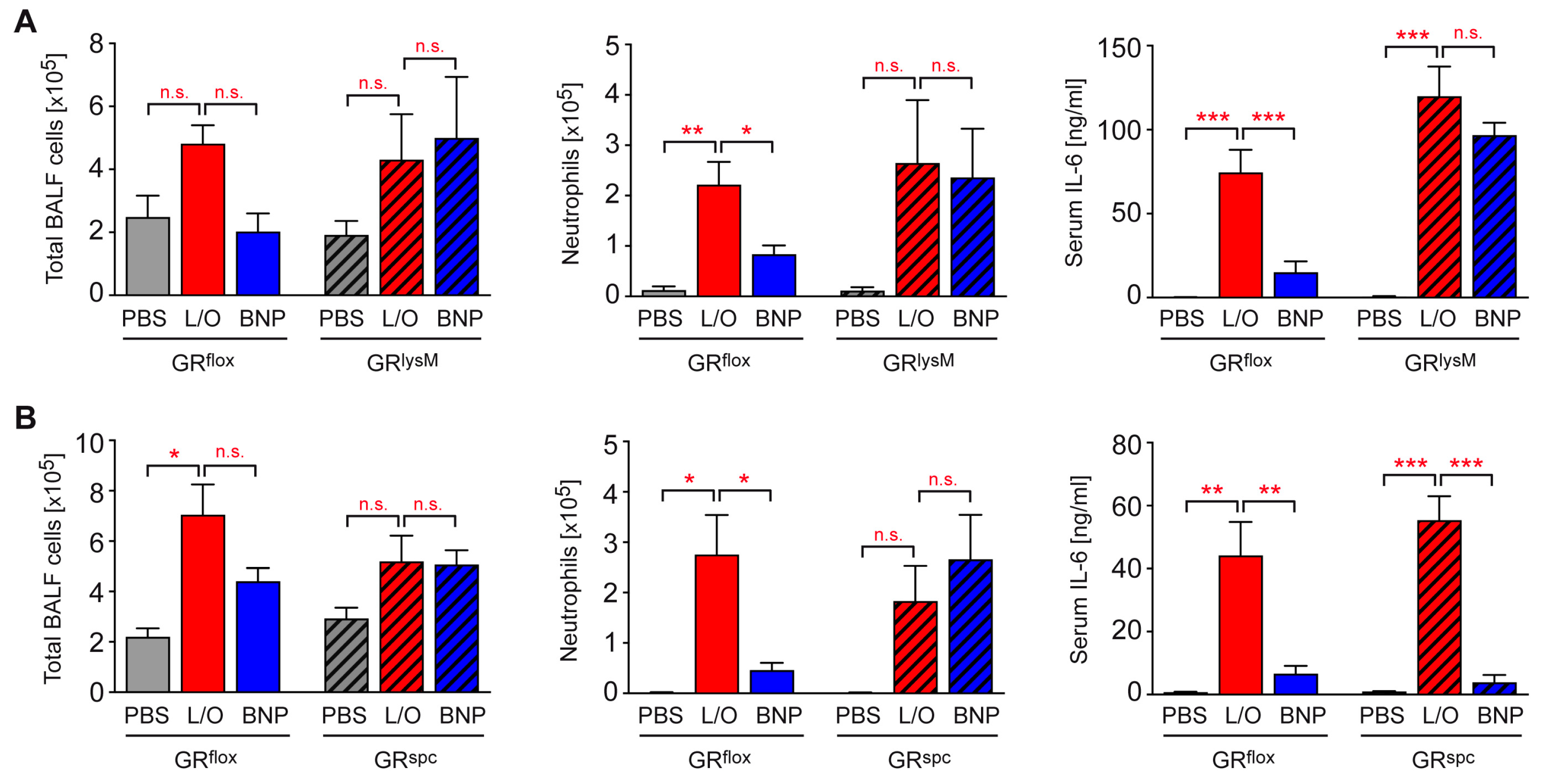
2.6. Identification of BNP Target Cells in ALI Therapy
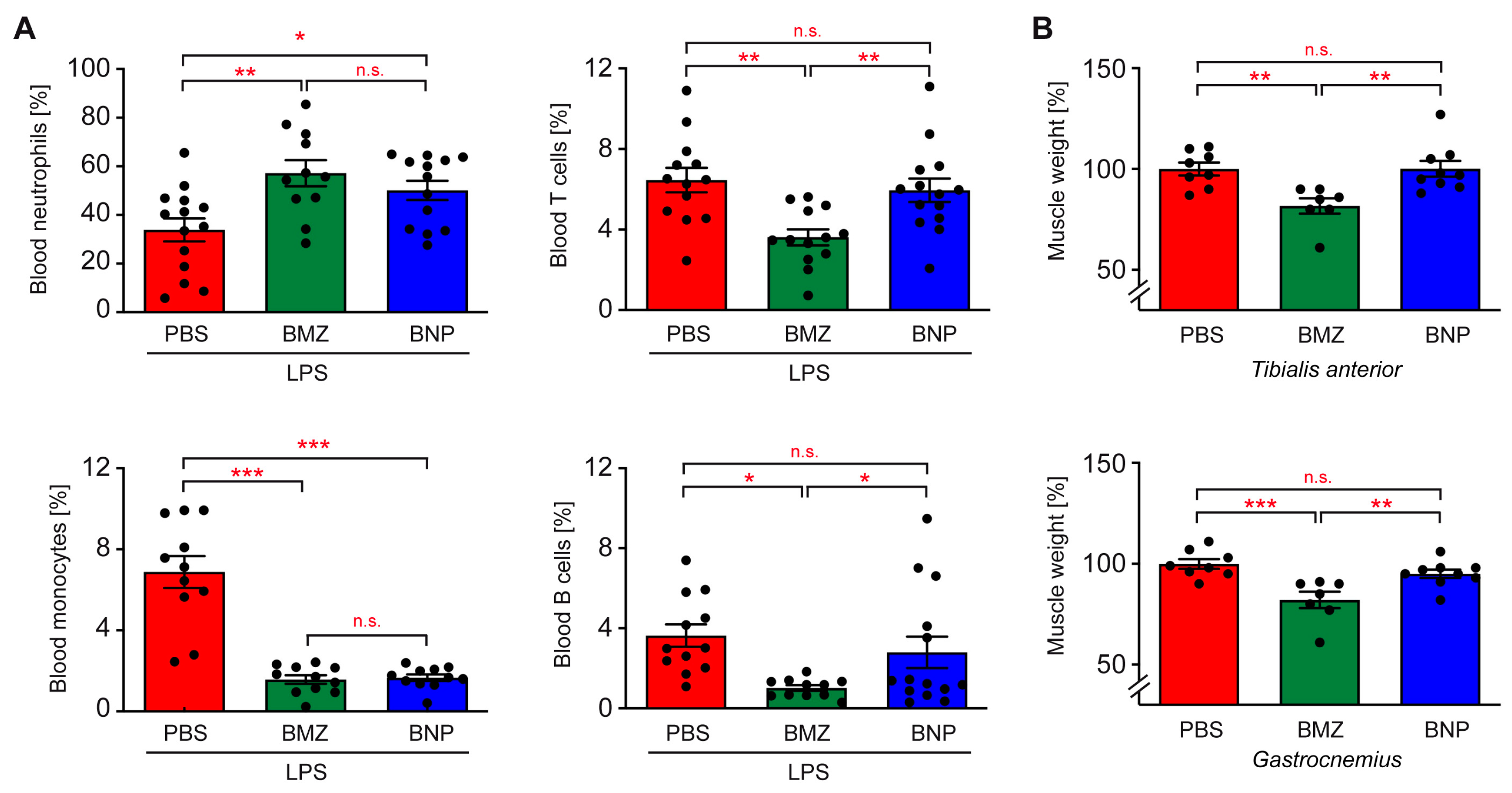
2.7. Cell-Type-Specific Effects of BNPs on Blood Leukocytes
2.8. Impact of the GC Nanoformulation on the Induction of Muscle Atrophy
3. Discussion
4. Materials and Methods
4.1. Animal Experimentation
4.2. Flow Cytometric Analyses
4.3. Histology
4.4. Quantitative RT-PCR Analysis
4.5. ELISA and Griess Test
4.6. Alveolar–Capillary Barrier Permeability Test
4.7. Nanoparticle Synthesis and Characterization
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, E.R.; Matthay, M.A. Acute lung injury: Epidemiology, pathogenesis, and treatment. J. Aerosol Med. Pulm. Drug Deliv. 2010, 23, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Chuquimia, O.D.; Petursdottir, D.H.; Rahman, M.J.; Hartl, K.; Singh, M.; Fernandez, C. The role of alveolar epithelial cells in initiating and shaping pulmonary immune responses: Communication between innate and adaptive immune systems. PLoS ONE 2012, 7, e32125. [Google Scholar] [CrossRef] [PubMed]
- Fehrenbach, H. Alveolar epithelial type II cell: Defender of the alveolus revisited. Respir. Res. 2001, 2, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Scozzi, D.; Liao, F.; Krupnick, A.S.; Kreisel, D.; Gelman, A.E. The role of neutrophil extracellular traps in acute lung injury. Front. Immunol. 2022, 13, 953195. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.C.; Fessler, M.B. Regulatory mechanisms of neutrophil migration from the circulation to the airspace. Cell Mol. Life Sci. 2021, 78, 4095–4124. [Google Scholar] [CrossRef] [PubMed]
- Matthay, M.A.; Zemans, R.L.; Zimmerman, G.A.; Arabi, Y.M.; Beitler, J.R.; Mercat, A.; Herridge, M.; Randolph, A.G.; Calfee, C.S. Acute respiratory distress syndrome. Nat. Rev. Dis. Primers 2019, 5, 18. [Google Scholar] [CrossRef] [PubMed]
- Diaz, J.V.; Brower, R.; Calfee, C.S.; Matthay, M.A. Therapeutic strategies for severe acute lung injury. Crit. Care Med. 2010, 38, 1644–1650. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ge, P.; Liu, J.; Luo, Y.; Guo, H.; Zhang, G.; Xu, C.; Chen, H. Glucocorticoid Treatment in Acute Respiratory Distress Syndrome: An Overview on Mechanistic Insights and Clinical Benefit. Int. J. Mol. Sci. 2023, 24, 12138. [Google Scholar] [CrossRef]
- Meduri, G.U.; Bridges, L.; Shih, M.C.; Marik, P.E.; Siemieniuk, R.A.C.; Kocak, M. Prolonged glucocorticoid treatment is associated with improved ARDS outcomes: Analysis of individual patients’ data from four randomized trials and trial-level meta-analysis of the updated literature. Intensive Care Med. 2016, 42, 829–840. [Google Scholar] [CrossRef]
- Steinberg, K.P.; Hudson, L.D.; Goodman, R.B.; Hough, C.L.; Lanken, P.N.; Hyzy, R.; Thompson, B.T.; Ancukiewicz, M.; National Heart, L.; Blood Institute Acute Respiratory Distress Syndrome Clinical Trials, N. Efficacy and safety of corticosteroids for persistent acute respiratory distress syndrome. N. Engl. J. Med. 2006, 354, 1671–1684. [Google Scholar] [CrossRef]
- Li, L.; Huang, Q.; Wang, D.C.; Ingbar, D.H.; Wang, X. Acute lung injury in patients with COVID-19 infection. Clin. Transl. Med. 2020, 10, 20–27. [Google Scholar] [CrossRef]
- Group, R.C.; Horby, P.; Lim, W.S.; Emberson, J.R.; Mafham, M.; Bell, J.L.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; et al. Dexamethasone in Hospitalized Patients with COVID-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar] [CrossRef]
- The WHO Rapid Evidence Appraisal for COVID-19 Therapies (REACT) Working Group; Sterne, J.A.C.; Murthy, S.; Diaz, J.V.; Slutsky, A.S.; Villar, J.; Angus, D.C.; Annane, D.; Azevedo, L.C.P.; Berwanger, O.; et al. Association between Administration of Systemic Corticosteroids and Mortality among Critically Ill Patients with COVID-19: A Meta-analysis. JAMA 2020, 324, 1330–1341. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, S.D.; Amouret, A.; Muzzi, C.; Vettorazzi, S.; Tuckermann, J.P.; Lühder, F.; Reichardt, H.M. The Role of Glucocorticoids in Inflammatory Diseases. Cells 2021, 10, 2921. [Google Scholar] [CrossRef] [PubMed]
- Escoter-Torres, L.; Caratti, G.; Mechtidou, A.; Tuckermann, J.; Uhlenhaut, N.H.; Vettorazzi, S. Fighting the Fire: Mechanisms of Inflammatory Gene Regulation by the Glucocorticoid Receptor. Front. Immunol. 2019, 10, 1859. [Google Scholar] [CrossRef] [PubMed]
- Erlacher, M.; Michalak, E.M.; Kelly, P.N.; Labi, V.; Niederegger, H.; Coultas, L.; Adams, J.M.; Strasser, A.; Villunger, A. BH3-only proteins Puma and Bim are rate-limiting for gamma-radiation- and glucocorticoid-induced apoptosis of lymphoid cells in vivo. Blood 2005, 106, 4131–4138. [Google Scholar] [CrossRef]
- Zacharchuk, C.M.; Mercep, M.; Chakraborti, P.K.; Simons, S.S., Jr.; Ashwell, J.D. Programmed T lymphocyte death. Cell activation- and steroid-induced pathways are mutually antagonistic. J. Immunol. 1990, 145, 4037–4045. [Google Scholar] [CrossRef]
- Almawi, W.Y.; Beyhum, H.N.; Rahme, A.A.; Rieder, M.J. Regulation of cytokine and cytokine receptor expression by glucocorticoids. J. Leukoc. Biol. 1996, 60, 563–572. [Google Scholar] [CrossRef]
- Schweingruber, N.; Fischer, H.J.; Fischer, L.; van den Brandt, J.; Karabinskaya, A.; Labi, V.; Villunger, A.; Kretzschmar, B.; Huppke, P.; Simons, M.; et al. Chemokine-mediated redirection of T cells constitutes a critical mechanism of glucocorticoid therapy in autoimmune CNS responses. Acta Neuropathol. 2014, 127, 713–729. [Google Scholar] [CrossRef]
- Baschant, U.; Tuckermann, J. The role of the glucocorticoid receptor in inflammation and immunity. J. Steroid Biochem. Mol. Biol. 2010, 120, 69–75. [Google Scholar] [CrossRef]
- Klassen, C.; Karabinskaya, A.; Dejager, L.; Vettorazzi, S.; Van Moorleghem, J.; Lühder, F.; Meijsing, S.H.; Tuckermann, J.P.; Bohnenberger, H.; Libert, C.; et al. Airway Epithelial Cells Are Crucial Targets of Glucocorticoids in a Mouse Model of Allergic Asthma. J. Immunol. 2017, 199, 48–61. [Google Scholar] [CrossRef]
- Stratakis, C.A. An update on Cushing syndrome in pediatrics. Ann. Endocrinol. 2018, 79, 125–131. [Google Scholar] [CrossRef]
- Schäcke, H.; Docke, W.D.; Asadullah, K. Mechanisms involved in the side effects of glucocorticoids. Pharmacol. Ther. 2002, 96, 23–43. [Google Scholar] [CrossRef]
- Caplan, A.; Fett, N.; Rosenbach, M.; Werth, V.P.; Micheletti, R.G. Prevention and management of glucocorticoid-induced side effects: A comprehensive review: Ocular, cardiovascular, muscular, and psychiatric side effects and issues unique to pediatric patients. J. Am. Acad. Dermatol. 2017, 76, 201–207. [Google Scholar] [CrossRef]
- Matute-Bello, G.; Frevert, C.W.; Martin, T.R. Animal models of acute lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 295, L379–L399. [Google Scholar] [CrossRef]
- Tu, G.W.; Shi, Y.; Zheng, Y.J.; Ju, M.J.; He, H.Y.; Ma, G.G.; Hao, G.W.; Luo, Z. Glucocorticoid attenuates acute lung injury through induction of type 2 macrophage. J. Transl. Med. 2017, 15, 181. [Google Scholar] [CrossRef]
- Vettorazzi, S.; Bode, C.; Dejager, L.; Frappart, L.; Shelest, E.; Klassen, C.; Tasdogan, A.; Reichardt, H.M.; Libert, C.; Schneider, M.; et al. Glucocorticoids limit acute lung inflammation in concert with inflammatory stimuli by induction of SphK1. Nat. Commun. 2015, 6, 7796. [Google Scholar] [CrossRef]
- Wepler, M.; Preuss, J.M.; Merz, T.; Hartmann, C.; Wachter, U.; McCook, O.; Vogt, J.; Kress, S.; Groger, M.; Fink, M.; et al. Impaired Glucocorticoid Receptor Dimerization Aggravates LPS-Induced Circulatory and Pulmonary Dysfunction. Front. Immunol. 2019, 10, 3152. [Google Scholar] [CrossRef] [PubMed]
- Lühder, F.; Reichardt, H.M. Novel Drug Delivery Systems Tailored for Improved Administration of Glucocorticoids. Int. J. Mol. Sci. 2017, 18, 1836. [Google Scholar] [CrossRef] [PubMed]
- Linker, R.A.; Weller, C.; Luhder, F.; Mohr, A.; Schmidt, J.; Knauth, M.; Metselaar, J.M.; Gold, R. Liposomal glucocorticosteroids in treatment of chronic autoimmune demyelination: Long-term protective effects and enhanced efficacy of methylprednisolone formulations. Exp. Neurol. 2008, 211, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Jiang, J.; Chen, W.; Jiang, H.; Zhang, Z.; Sun, X. Targeted delivery of low-dose dexamethasone using PCL-PEG micelles for effective treatment of rheumatoid arthritis. J. Control. Release 2016, 230, 64–72. [Google Scholar] [CrossRef]
- Ren, K.; Dusad, A.; Yuan, F.; Yuan, H.; Purdue, P.E.; Fehringer, E.V.; Garvin, K.L.; Goldring, S.R.; Wang, D. Macromolecular prodrug of dexamethasone prevents particle-induced peri-implant osteolysis with reduced systemic side effects. J. Control. Release 2014, 175, 1–9. [Google Scholar] [CrossRef]
- Heck, J.G.; Napp, J.; Simonato, S.; Mollmer, J.; Lange, M.; Reichardt, H.M.; Staudt, R.; Alves, F.; Feldmann, C. Multifunctional phosphate-based inorganic-organic hybrid nanoparticles. J. Am. Chem. Soc. 2015, 137, 7329–7336. [Google Scholar] [CrossRef]
- Kaiser, T.K.; Khorenko, M.; Moussavi, A.; Engelke, M.; Boretius, S.; Feldmann, C.; Reichardt, H.M. Highly selective organ distribution and cellular uptake of inorganic-organic hybrid nanoparticles customized for the targeted delivery of glucocorticoids. J. Control. Release 2020, 319, 360–370. [Google Scholar] [CrossRef]
- Kaiser, T.K.; Li, H.; Rossmann, L.; Reichardt, S.D.; Bohnenberger, H.; Feldmann, C.; Reichardt, H.M. Glucocorticoids delivered by inorganic-organic hybrid nanoparticles mitigate acute graft-versus-host disease and sustain graft-versus-leukemia activity. Eur. J. Immunol. 2020, 50, 1220–1233. [Google Scholar] [CrossRef]
- Montes-Cobos, E.; Ring, S.; Fischer, H.J.; Heck, J.; Strauss, J.; Schwaninger, M.; Reichardt, S.D.; Feldmann, C.; Lühder, F.; Reichardt, H.M. Targeted delivery of glucocorticoids to macrophages in a mouse model of multiple sclerosis using inorganic-organic hybrid nanoparticles. J. Control. Release 2017, 245, 157–169. [Google Scholar] [CrossRef]
- Kunze, J.; Koelling, S.; Reich, M.; Wimmer, M.A. Use of ultrasonic nebulizer with desolvator membrane for the determination of titanium and zirconium in human serum by means of inductively coupled plasma--mass spectroscopy. Fresenius J. Anal. Chem. 2000, 366, 165–166. [Google Scholar] [CrossRef]
- Vandewalle, J.; Libert, C. Glucocorticoids in Sepsis: To Be or Not to Be. Front. Immunol. 2020, 11, 1318. [Google Scholar] [CrossRef]
- Wu, Y.; Ma, J.; Woods, P.S.; Chesarino, N.M.; Liu, C.; Lee, L.J.; Nana-Sinkam, S.P.; Davis, I.C. Selective targeting of alveolar type II respiratory epithelial cells by anti-surfactant protein-C antibody-conjugated lipoplexes. J. Control. Release 2015, 203, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Rubenfeld, G.D.; Caldwell, E.; Peabody, E.; Weaver, J.; Martin, D.P.; Neff, M.; Stern, E.J.; Hudson, L.D. Incidence and outcomes of acute lung injury. N. Engl. J. Med. 2005, 353, 1685–1693. [Google Scholar] [CrossRef] [PubMed]
- Festic, E.; Carr, G.E.; Cartin-Ceba, R.; Hinds, R.F.; Banner-Goodspeed, V.; Bansal, V.; Asuni, A.T.; Talmor, D.; Rajagopalan, G.; Frank, R.D.; et al. Randomized Clinical Trial of a Combination of an Inhaled Corticosteroid and Beta Agonist in Patients at Risk of Developing the Acute Respiratory Distress Syndrome. Crit. Care Med. 2017, 45, 798–805. [Google Scholar] [CrossRef]
- Munch, M.W.; Granholm, A.; Malaska, J.; Stasek, J.; Rodriguez, P.O.; Pitre, T.; Wilson, R.; Savovic, J.; Rochwerg, B.; Svobodnik, A.; et al. Dexamethasone doses in patients with COVID-19 and hypoxia: A systematic review and meta-analysis. Acta Anaesthesiol. Scand. 2023; online ahead of print. [Google Scholar] [CrossRef]
- Dahlberg, E.; Thalen, A.; Brattsand, R.; Gustafsson, J.A.; Johansson, U.; Roempke, K.; Saartok, T. Correlation between chemical structure, receptor binding, and biological activity of some novel, highly active, 16 alpha, 17 alpha-acetal-substituted glucocorticoids. Mol. Pharmacol. 1984, 25, 70–78. [Google Scholar]
- Ozbakir, B.; Crielaard, B.J.; Metselaar, J.M.; Storm, G.; Lammers, T. Liposomal corticosteroids for the treatment of inflammatory disorders and cancer. J. Control. Release 2014, 190, 624–636. [Google Scholar] [CrossRef]
- Noetzlin, S.; Breville, G.; Seebach, J.D.; Gastaldi, G. Short-term glucocorticoid-related side effects and adverse reactions: A narrative review and practical approach. Swiss Med. Wkly. 2022, 152, w30088. [Google Scholar] [CrossRef]
- Liu, D.; Ahmet, A.; Ward, L.; Krishnamoorthy, P.; Mandelcorn, E.D.; Leigh, R.; Brown, J.P.; Cohen, A.; Kim, H. A practical guide to the monitoring and management of the complications of systemic corticosteroid therapy. Allergy Asthma Clin. Immunol. 2013, 9, 30. [Google Scholar] [CrossRef]
- Levin, O.S.; Polunina, A.G.; Demyanova, M.A.; Isaev, F.V. Steroid myopathy in patients with chronic respiratory diseases. J. Neurol. Sci. 2014, 338, 96–101. [Google Scholar] [CrossRef]
- Bodine, S.C.; Furlow, J.D. Glucocorticoids and Skeletal Muscle. Adv. Exp. Med. Biol. 2015, 872, 145–176. [Google Scholar] [CrossRef]
- Rock, J.R.; Barkauskas, C.E.; Cronce, M.J.; Xue, Y.; Harris, J.R.; Liang, J.; Noble, P.W.; Hogan, B.L. Multiple stromal populations contribute to pulmonary fibrosis without evidence for epithelial to mesenchymal transition. Proc. Natl. Acad. Sci. USA 2011, 108, E1475–E1483. [Google Scholar] [CrossRef] [PubMed]
- Clausen, B.E.; Burkhardt, C.; Reith, W.; Renkawitz, R.; Forster, I. Conditional gene targeting in macrophages and granulocytes using LysMcre mice. Transgenic Res. 1999, 8, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, S.D.; Lühder, F.; Wiegers, G.J.; Reichardt, H.M. A flow cytometric approach to study glucocorticoid receptor expression in immune cell subpopulations of genetically engineered mice. Immunol. Lett. 2021, 233, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Kleiman, A.; Hübner, S.; Rodriguez Parkitna, J.M.; Neumann, A.; Hofer, S.; Weigand, M.A.; Bauer, M.; Schmid, W.; Schütz, G.; Libert, C.; et al. Glucocorticoid receptor dimerization is required for survival in septic shock via suppression of interleukin-1 in macrophages. FASEB J. 2012, 26, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Cossarizza, A.; Chang, H.D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef]
- Moitra, J.; Sammani, S.; Garcia, J.G. Re-evaluation of Evans Blue dye as a marker of albumin clearance in murine models of acute lung injury. Transl. Res. 2007, 150, 253–265. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albers, G.J.; Amouret, A.; Ciupka, K.; Montes-Cobos, E.; Feldmann, C.; Reichardt, H.M. Glucocorticoid Nanoparticles Show Full Therapeutic Efficacy in a Mouse Model of Acute Lung Injury and Concomitantly Reduce Adverse Effects. Int. J. Mol. Sci. 2023, 24, 16843. https://doi.org/10.3390/ijms242316843
Albers GJ, Amouret A, Ciupka K, Montes-Cobos E, Feldmann C, Reichardt HM. Glucocorticoid Nanoparticles Show Full Therapeutic Efficacy in a Mouse Model of Acute Lung Injury and Concomitantly Reduce Adverse Effects. International Journal of Molecular Sciences. 2023; 24(23):16843. https://doi.org/10.3390/ijms242316843
Chicago/Turabian StyleAlbers, Gesa J., Agathe Amouret, Katrin Ciupka, Elena Montes-Cobos, Claus Feldmann, and Holger M. Reichardt. 2023. "Glucocorticoid Nanoparticles Show Full Therapeutic Efficacy in a Mouse Model of Acute Lung Injury and Concomitantly Reduce Adverse Effects" International Journal of Molecular Sciences 24, no. 23: 16843. https://doi.org/10.3390/ijms242316843