
Sodium-Doped 3-Amino-4-hydroxybenzoic Acid: Rediscovered Matrix for Direct MALDI Glycotyping of O-Linked Glycopeptides and Intact Mucins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. MALDI-ISD Analysis of O-Linked Cyclic Glycopeptides
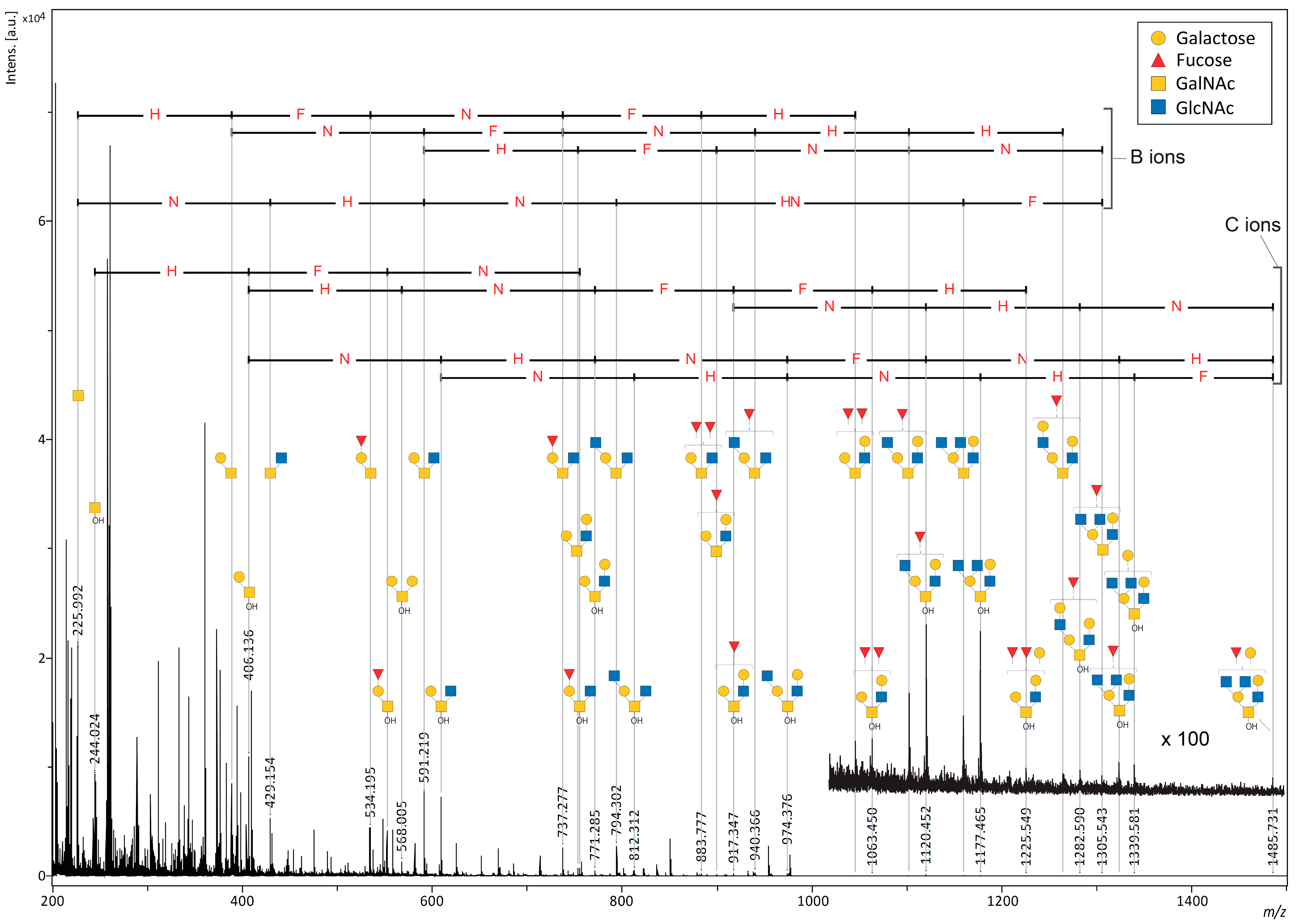
2.2. MALDI-ISD O-Glycan Analysis of Intact Mucin
2.3. Pseudo MS3 O-Glycan Analysis of cAFGPs and Intact Mucin with the AHB/Na Matrix
3. Materials and Methods
3.1. Materials and Reagents
3.2. Matrices
3.3. Sample Preparation for MALDI TOF MS Analysis
3.4. MALDI-TOF and TOF/TOF MS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Varki, A. Biological roles of glycans. Glycobiology 2017, 27, 3–49. [Google Scholar] [CrossRef] [PubMed]
- Cazet, A.; Julien, S.; Bobowski, M.; Burchell, J.; Delannoy, P. Tumour-associated carbohydrate antigens in breast cancer. Breast Cancer Res. 2010, 12, 204. [Google Scholar] [CrossRef] [PubMed]
- Kudelka, M.R.; Ju, T.; Heimburg-Molinaro, J.; Cummings, R.D. Simple sugars to complex disease—Mucin-type O-glycans in cancer. Adv. Cancer Res. 2015, 126, 53–135. [Google Scholar] [PubMed]
- Gizaw, S.T.; Gaunitz, S.; Novotny, M.V. Highly Sensitive O-Glycan Profiling for Human Serum Proteins Reveals Gender-Dependent Changes in Colorectal Cancer Patients. Anal. Chem. 2019, 91, 6180–6189. [Google Scholar] [CrossRef]
- Borgert, A.; Foley, B.L.; Live, D. Contrasting the conformational effects of alpha-O-GalNAc and alpha-O-Man glycan protein modifications and their impact on the mucin-like region of alpha-dystroglycan. Glycobiology 2021, 31, 649–661. [Google Scholar] [CrossRef]
- Devries, A.L.; Wohlschlag, D.E. Freezing Resistance in Some Antarctic Fishes. Science 1969, 163, 1073–1075. [Google Scholar] [CrossRef]
- Tachibana, Y.; Fletcher, G.L.; Fujitani, N.; Tsuda, S.; Monde, K.; Nishimura, S.I. Antifreeze glycoproteins: Elucidation of the structural motifs that are essential for antifreeze activity. Angew. Chem. Int. Ed. 2004, 43, 856–862. [Google Scholar] [CrossRef]
- Tanaka, K.; Waki, H.; Ido, Y.; Akita, S.; Yoshida, Y.; Yoshida, T.; Matsuo, T. Protein and polymer analyses up to m/z 100 000 by laser ionization time-of-flight mass spectrometry. Rapid Commun. Mass Spectrum. 1988, 2, 151–153. [Google Scholar] [CrossRef]
- Karas, M.; Hillenkamp, F. Laser Desorption Ionization of Proteins with Molecular Masses Exceeding 10000 Daltons. Anal. Chem. 1988, 60, 2299–2301. [Google Scholar] [CrossRef]
- Karas, M.; Bahr, U.; Ingendoh, A.; Nordhoff, E.; Stahl, B.; Strupat, K.; Hillenkamp, F. Principles and Applications of Matrix-Assisted Uv Laser Desorption Ionization Mass-Spectrometry. Anal. Chim. Acta 1990, 241, 175–185. [Google Scholar] [CrossRef]
- Karas, M.; Krüger, R. Ion formation in MALDI: The cluster ionization mechanism. Chem. Rev. 2003, 103, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Mock, K.K.; Davey, M.; Cottrell, J.S. The Analysis of Underivatised Oligosaccharides by Matrix-Assisted Laser Desorption Mass-Spectrometry. Biochem. Bioph. Res. Commun. 1991, 177, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Stahl, B.; Steup, M.; Karas, M.; Hillenkamp, F. Analysis of Neutral Oligosaccharides by Matrix-Assisted Laser Desorption—Ionization Mass-Spectrometry. Anal. Chem. 1991, 63, 1463–1466. [Google Scholar] [CrossRef]
- Strupat, K.; Karas, M.; Hillenkamp, F. 2,5-Dihydroxybenzoic Acid—A New Matrix for Laser Desorption Ionization Mass-Spectrometry. Int. J. Mass Spectrom. 1991, 111, 89–102. [Google Scholar] [CrossRef]
- Harvey, D.J. Analysis of carbohydrates and glycoconjugates by matrix-assisted laser desorption/ionization mass spectrometry: An update for 2019–2020. Mass Spectrom. Rev. 2023, 42, 1984–2206. [Google Scholar] [CrossRef]
- Wang, J.Q.; Zhao, J.; Nie, S.P.; Xie, M.Y.; Li, S.P. MALDI mass spectrometry in food carbohydrates analysis: A review of recent researches. Food. Chem. 2023, 399, 133968. [Google Scholar] [CrossRef] [PubMed]
- Snovida, S.I.; Chen, V.C.; Perreault, H. Use of a 2,5-dihydroxybenzoic acid/aniline MALDI matrix for improved detection and on-target derivatization of glycans: A preliminary report. Anal. Chem. 2006, 78, 8561–8568. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Shen, S.S.; Wu, D.T.; Cai, P.F.; Pan, Y.J. Novel ionic liquid matrices for qualitative and quantitative detection of carbohydrates by matrix assisted laser desorption/ionization mass spectrometry. Anal. Chim. Acta 2017, 985, 114–120. [Google Scholar] [CrossRef]
- Urakami, S.; Hinou, H. Glycan-Selective MALDI In-Source Decay Analysis of Intact Glycoproteins. Anal. Sense 2022, 2, e202100040. [Google Scholar] [CrossRef]
- Urakami, S.; Hinou, H. Direct MALDI Glycotyping of Glycoproteins Toward Practical Subtyping of Biological Samples. ACS Omega 2022, 7, 39280–39286. [Google Scholar] [CrossRef]
- Suckau, D.; Resemann, A. T3-Sequencing: Targeted Characterization of the N- and C-Terminal of Undigested Proteins by Mass Spectrometry. Anal. Chem. 2003, 75, 5817–5824. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, D.; Smargiasso, N.; De Pauw, E. New Approach for Pseudo-MS3 Analysis of Peptides and Proteins via MALDI In-Source Decay Using Radical Recombination with 1,5-Diaminonaphthalene. Anal. Chem. 2014, 86, 2451–2457. [Google Scholar] [CrossRef] [PubMed]
- Hachisu, M.; Hinou, H.; Takamichi, M.; Tsuda, S.; Koshidaa, S.; Nishimura, S.I. One-pot synthesis of cyclic antifreeze glycopeptides. Chem. Commun. 2009, 1641–1643. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, Y.; Iwamoto, S.; Tanaka, K. Rapid sequencing and disulfide mapping of peptides containing disulfide bonds by using 1,5-diaminonaphthalene as a reductive matrix. J. Mass Spectrom. 2006, 41, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, D.; Smargiasso, N.; Quinton, L.; De Pauw, E. Peptide backbone fragmentation initiated by side-chain loss at cysteine residue in matrix-assisted laser desorption/ionization in-source decay mass spectrometry. J. Mass Spectrom. 2013, 48, 352–360. [Google Scholar] [CrossRef]
- Corfield, A.P. Mucins: A biologically relevant glycan barrier in mucosal protection. BBA-Gen. Subjects 2015, 1850, 236–252. [Google Scholar] [CrossRef]
- Johansson, M.E.; Sjovall, H.; Hansson, G.C. The gastrointestinal mucus system in health and disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 352–361. [Google Scholar] [CrossRef]
- Barada, E.; Hinou, H. BOA/DHB/Na: An Efficient UV-MALDI Matrix for High-Sensitivity and Auto-Tagging Glycomics. Int. J. Mol. Sci. 2022, 23, 12510. [Google Scholar] [CrossRef]
- Yamada, K.; Hirabayashi, J.; Kakehi, K. Analysis of O-Glycans as 9-Fluorenylmethyl Derivatives and Its Application to the Studies on Glycan Array. Anal. Chem. 2013, 85, 3325–3333. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urakami, S.; Hinou, H. Sodium-Doped 3-Amino-4-hydroxybenzoic Acid: Rediscovered Matrix for Direct MALDI Glycotyping of O-Linked Glycopeptides and Intact Mucins. Int. J. Mol. Sci. 2023, 24, 16836. https://doi.org/10.3390/ijms242316836
Urakami S, Hinou H. Sodium-Doped 3-Amino-4-hydroxybenzoic Acid: Rediscovered Matrix for Direct MALDI Glycotyping of O-Linked Glycopeptides and Intact Mucins. International Journal of Molecular Sciences. 2023; 24(23):16836. https://doi.org/10.3390/ijms242316836
Chicago/Turabian StyleUrakami, Shogo, and Hiroshi Hinou. 2023. "Sodium-Doped 3-Amino-4-hydroxybenzoic Acid: Rediscovered Matrix for Direct MALDI Glycotyping of O-Linked Glycopeptides and Intact Mucins" International Journal of Molecular Sciences 24, no. 23: 16836. https://doi.org/10.3390/ijms242316836