
Noncoupled Mitochondrial Respiration as Therapeutic Approach for the Treatment of Metabolic Diseases: Focus on Transgenic Animal Models

Abstract
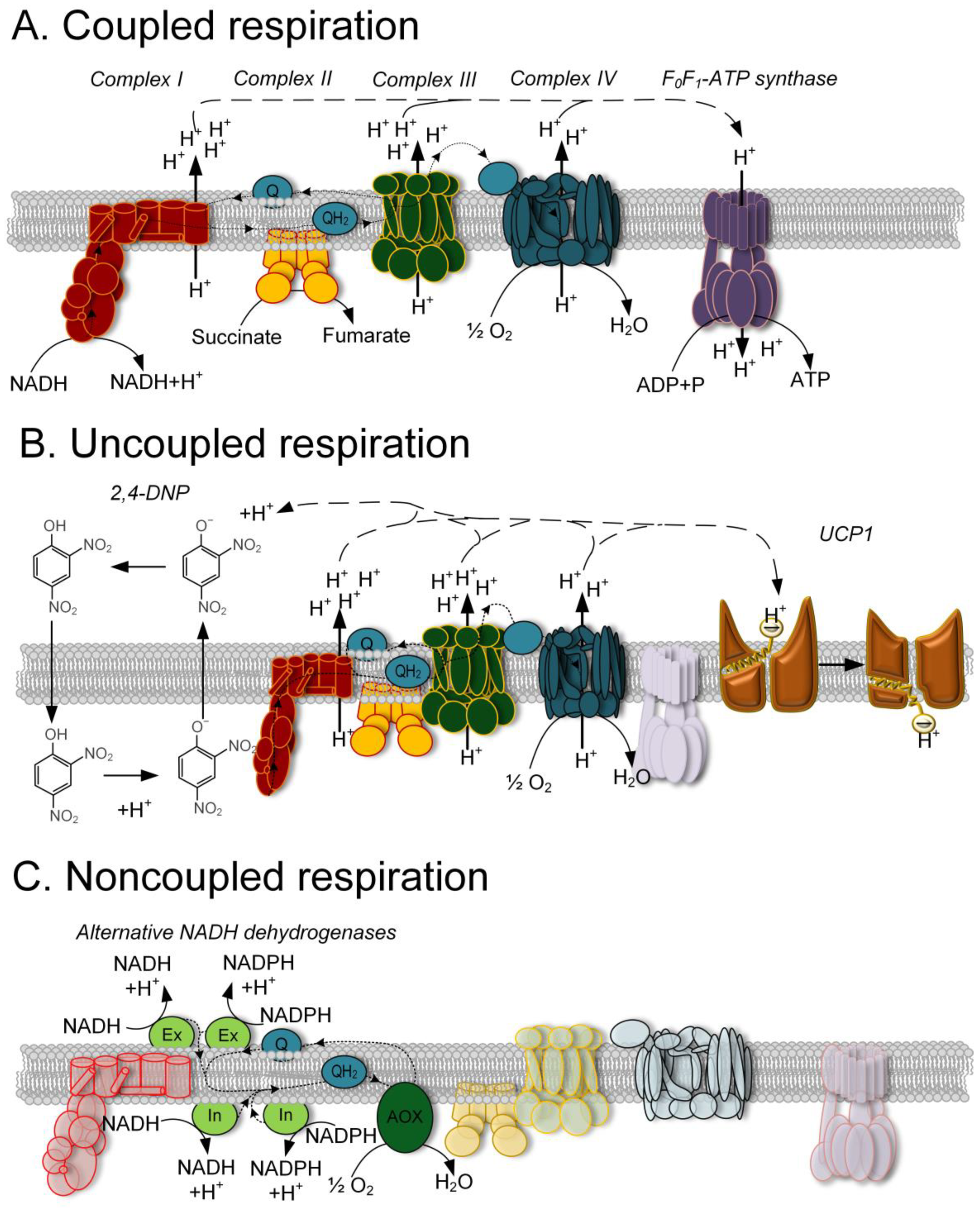
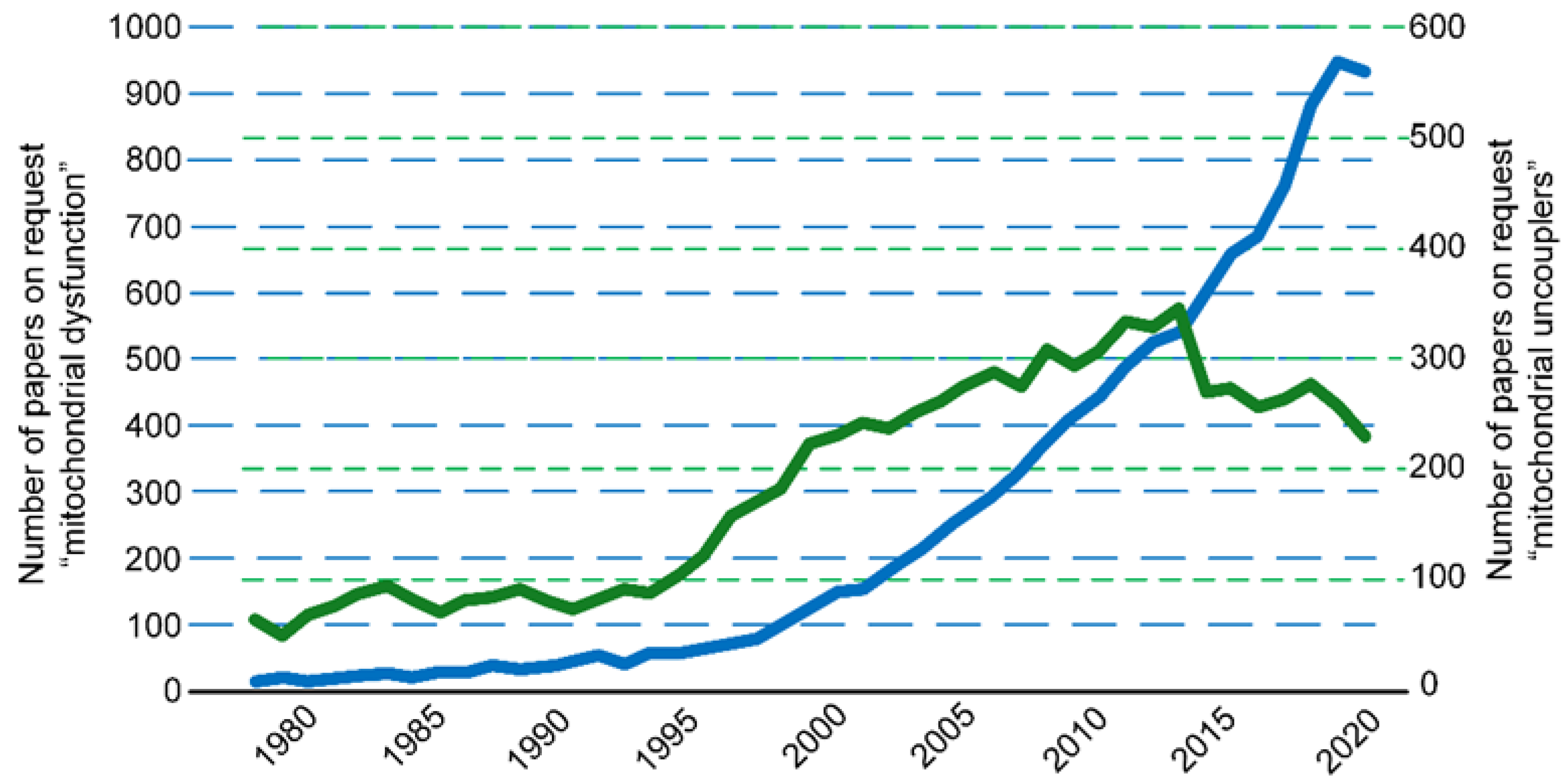
:1. Introduction
2. Chemical Uncouplers
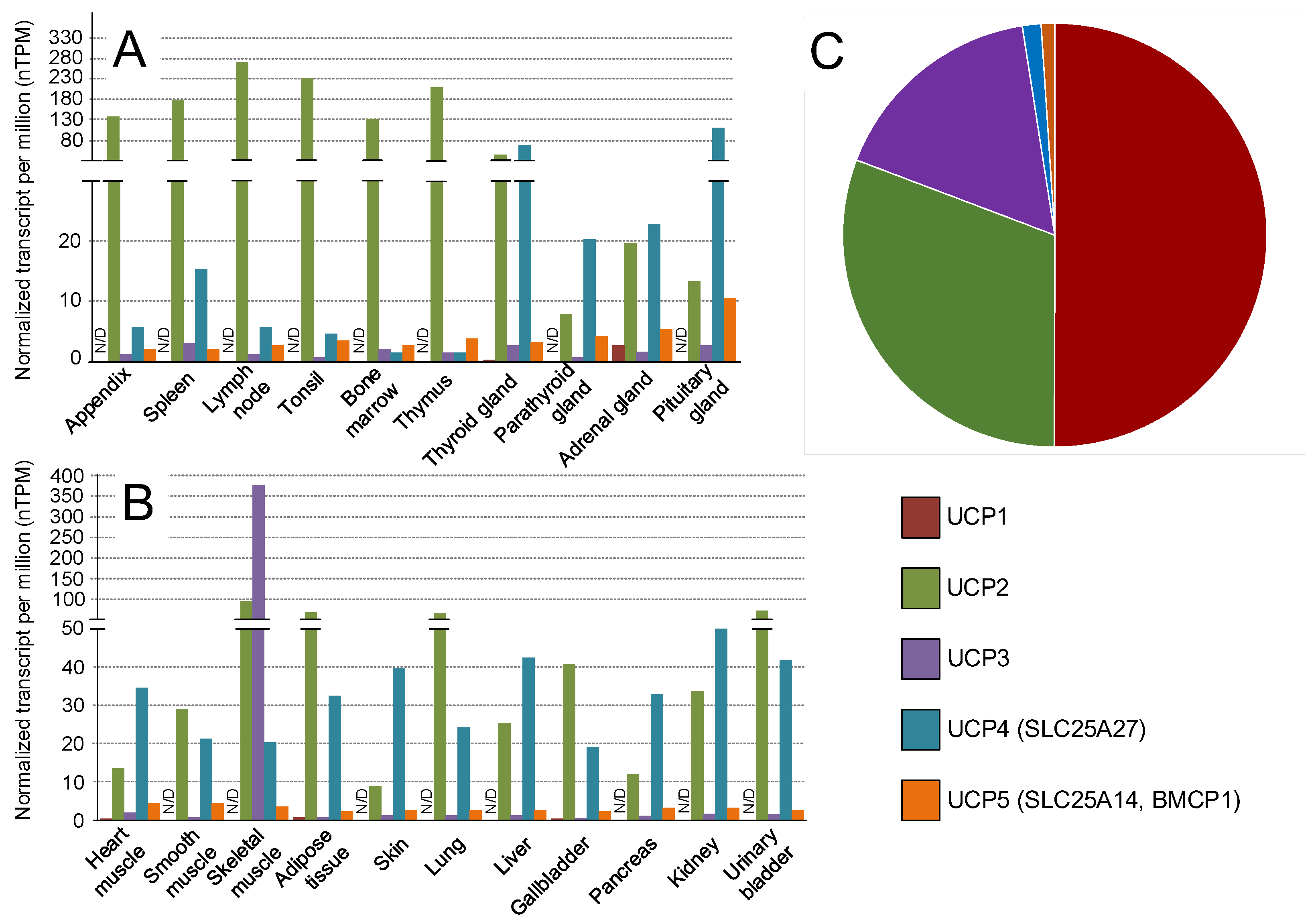
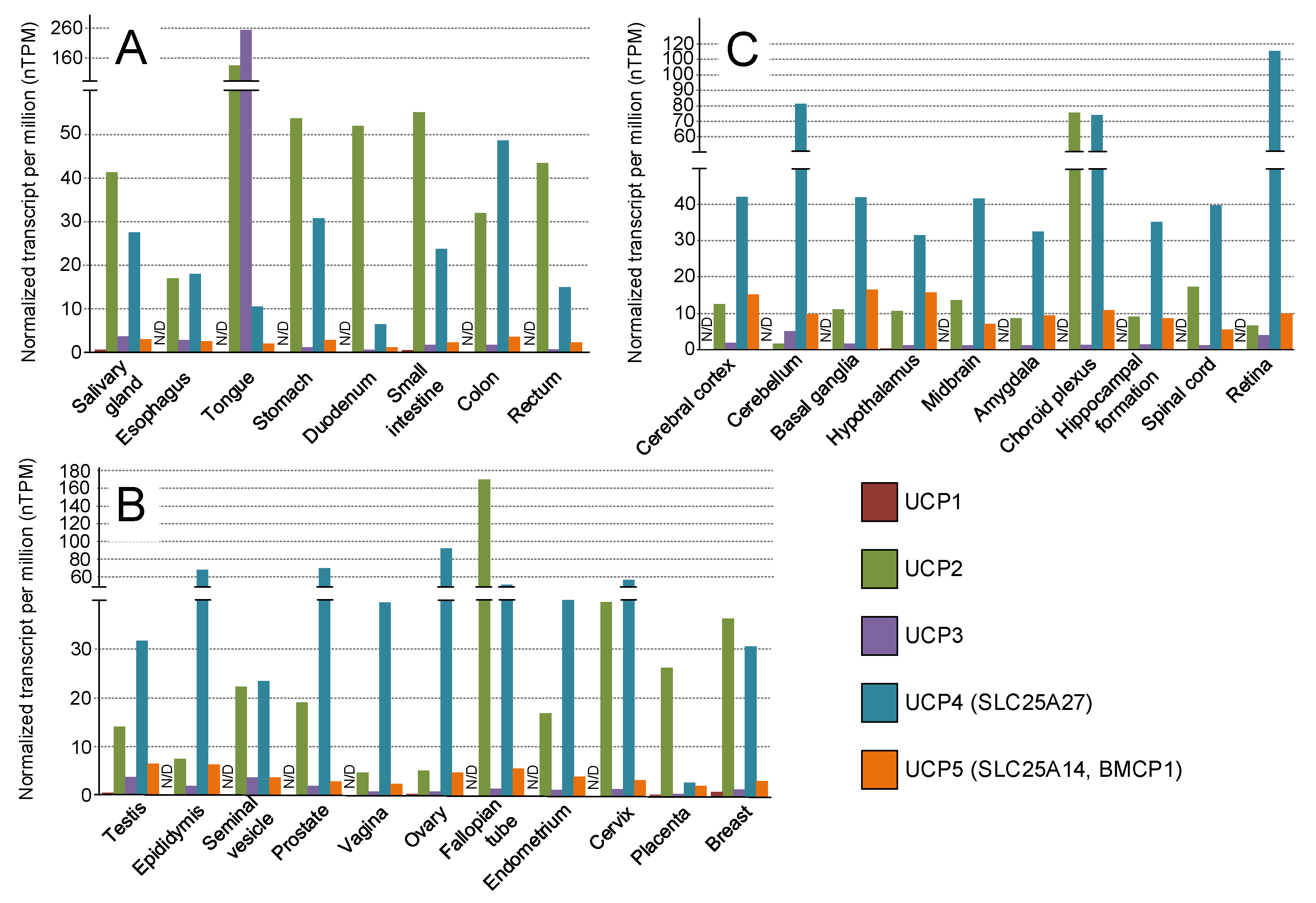
3. Transgenic Models Which Overexpress UCPs
4. Transgenic Animal Models in Which Components of Alternative Respiratory Pathways Are Expressed
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Skulachev, V.P. Uncoupling of Respiration and Phosphorylation. In Frontiers of Cellular Bioenergetics; Papa, S., Guerrieri, F., Eds.; Springer: Boston, MA, USA, 1999; pp. 89–118. [Google Scholar] [CrossRef]
- Jonckheere, A.I.; Smeitink, J.A.M.; Rodenburg, R.J.T. Mitochondrial ATP synthase: Architecture, function and pathology. J. Inherit. Metab. Dis. 2012, 35, 211–225. [Google Scholar] [CrossRef]
- Orrenius, S.; Gogvadze, V.; Zhivotovsky, B. Mitochondrial oxidative stress: Implications for cell death. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 143–183. [Google Scholar] [CrossRef]
- Rajman, L.; Chwalek, K.; Sinclair, D.A. Therapeutic Potential of NAD-Boosting Molecules: The in vivo Evidence. Cell Metab. 2018, 27, 529–547. [Google Scholar] [CrossRef]
- Fedorenko, A.; Lishko, P.V.; Kirichok, Y. Mechanism of fatty-acid-dependent UCP1 uncoupling in brown fat mitochondria. Cell 2012, 151, 400–413. [Google Scholar] [CrossRef] [PubMed]
- Kotova, E.A.; Antonenko, Y.N. Fifty Years of Research on Protonophores: Mitochondrial Uncoupling as a Basis for Therapeutic Action. Acta Nat. 2022, 14, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, R.; Johnson, E.; Byrne, F.L. Exploring the therapeutic potential of mitochondrial uncouplers in cancer. Mol. Metab. 2021, 51, 101222. [Google Scholar] [CrossRef]
- Eubel, H.; Heinemeyer, J.; Sunderhaus, S.; Braun, H.P. Respiratory chain supercomplexes in plant mitochondria. Plant Physiol. Biochem. 2004, 42, 937–942. [Google Scholar] [CrossRef]
- Rich, P.R.; Maréchal, A. The mitochondrial respiratory chain. Essays Biochem. 2010, 47, 1–23. [Google Scholar] [CrossRef]
- Sousa, J.S.; D′Imprima, E.; Vonck, J. Mitochondrial Respiratory Chain Complexes. Subcell. Biochem. 2018, 87, 167–227. [Google Scholar] [CrossRef]
- McDonald, A.; Vanlerberghe, G. Branched mitochondrial electron transport in the Animalia: Presence of alternative oxidase in several animal phyla. IUBMB Life 2004, 56, 333–341. [Google Scholar] [CrossRef]
- Vicente, J.A.; Peixoto, F.; Lopes, M.L.; Madeira, V.M. Differential sensitivities of plant and animal mitochondria to the herbicide paraquat. J. Biochem. Mol. Toxicol. 2001, 15, 322–330. [Google Scholar] [CrossRef]
- Fernandez-Vizarra, E.; Zeviani, M. Mitochondrial disorders of the OXPHOS system. FEBS Lett. 2021, 595, 1062–1106. [Google Scholar] [CrossRef] [PubMed]
- Brookes, P.S. Mitochondrial H(+) leak and ROS generation: An odd couple. Free Radic. Biol. Med. 2005, 38, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Childress, E.S.; Alexopoulos, S.J.; Hoehn, K.L.; Santos, W.L. Small Molecule Mitochondrial Uncouplers and Their Therapeutic Potential. J. Med. Chem. 2018, 61, 4641–4655. [Google Scholar] [CrossRef] [PubMed]
- Grundlingh, J.; Dargan, P.I.; El-Zanfaly, M.; Wood, D.M. 2,4-Dinitrophenol (DNP): A Weight Loss Agent with Significant Acute Toxicity and Risk of Death. J. Med. Toxicol. 2011, 7, 205–212. [Google Scholar] [CrossRef]
- Samartsev, V.N.; Semenova, A.A.; Dubinin, M.V. A Comparative Study of the Action of Protonophore Uncouplers and Decoupling Agents as Inducers of Free Respiration in Mitochondria in States 3 and 4: Theoretical and Experimental Approaches. Cell Biochem. Biophys. 2020, 78, 203–216. [Google Scholar] [CrossRef]
- Nicholls, D.G.; Ferguson, S.J. 2—Ion Transport Across Energy-Conserving Membranes. In Bioenergetics, 4th ed.; Nicholls, D.G., Ferguson, S.J., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 13–25. [Google Scholar] [CrossRef]
- Tainter, M.L.; Stockton, A.B.; Cutting, W.C. Use of dinitrophenol in obesity and related conditions: A progress report. JAMA 1933, 101, 1472–1475. [Google Scholar] [CrossRef]
- Tainter, M.L.; Cutting, W.C.; Hines, E. Effects of moderated doses of dinitrophenol on the energy exchange and nitrogen metabolism of patient under condition of restricted dietary. J. Pharmacol. Exp. Ther. 1935, 55, 326–353. [Google Scholar]
- Colman, E. Dinitrophenol and obesity: An early twentieth-century regulatory dilemma. Regul. Toxicol. Pharmacol. 2007, 48, 115–117. [Google Scholar] [CrossRef]
- Harper, J.A.; Dickinson, K.; Brand, M.D. Mitochondrial uncoupling as a target for drug development for the treatment of obesity. Obes. Rev. 2001, 2, 255–265. [Google Scholar] [CrossRef]
- Rognstad, R.; Katz, J. The effect of 2,4-dinitrophenol on adipose-tissue metabolism. Biochem. J. 1969, 111, 431–444. [Google Scholar] [CrossRef] [PubMed]
- El-Guindy, M.M.; Neder, A.C.; Gomes, C.B. 2,4-Dinitrophenol--mechanism of action. Cell Mol. Biol. Incl. Cyto Enzymol. 1981, 27, 399–402. [Google Scholar] [PubMed]
- Moffatt, E.J.; Miyamoto, M.D. Effect of sodium and calcium channel blockade on the increase in spontaneous transmitter release produced by the mitochondrial inhibitor, dinitrophenol. J. Pharmacol. Exp. Ther. 1988, 244, 613–618. [Google Scholar] [PubMed]
- Mudge, G.H. Electrolyte and water metabolism of rabbit kidney slices; effect of metabolic inhibitors. Am. J. Physiol. 1951, 167, 206–223. [Google Scholar] [CrossRef] [PubMed]
- Skulachev, V.P. Electric fields in coupling membranes. FEBS Lett. 1970, 11, 301–308. [Google Scholar] [CrossRef]
- Heytler, P.G.; Prichard, W.W. A new class of uncoupling agents—Carbonyl cyanide phenylhydrazones. Biochem. Biophys. Res. Commun. 1962, 7, 272–275. [Google Scholar] [CrossRef]
- Demine, S.; Renard, P.; Arnould, T. Mitochondrial Uncoupling: A Key Controller of Biological Processes in Physiology and Diseases. Cells 2019, 8, 795. [Google Scholar] [CrossRef]
- Duchen, M.R. Effects of metabolic inhibition on the membrane properties of isolated mouse primary sensory neurones. J. Physiol. 1990, 424, 387–409. [Google Scholar] [CrossRef]
- Park, K.S.; Jo, I.; Pak, K.; Bae, S.W.; Rhim, H.; Suh, S.H.; Park, S.; Zhu, M.; So, I.; Kim, K. FCCP depolarizes plasma membrane potential by activating proton and Na+ currents in bovine aortic endothelial cells. Pflugers Arch. 2002, 443, 344–352. [Google Scholar] [CrossRef]
- Urra, F.A.; Muñoz, F.; Córdova-Delgado, M.; Ramírez, M.P.; Peña-Ahumada, B.; Rios, M.; Cruz, P.; Ahumada-Castro, U.; Bustos, G.; Silva-Pavez, E.; et al. FR58P1a; a new uncoupler of OXPHOS that inhibits migration in triple-negative breast cancer cells via Sirt1/AMPK/β1-integrin pathway. Sci. Rep. 2018, 8, 13190. [Google Scholar] [CrossRef]
- Kenwood, B.M.; Weaver, J.L.; Bajwa, A.; Poon, I.K.; Byrne, F.L.; Murrow, B.A.; Calderone, J.A.; Huang, L.; Divakaruni, A.S.; Tomsig, J.L.; et al. Identification of a novel mitochondrial uncoupler that does not depolarize the plasma membrane. Mol. Metab. 2013, 3, 114–123. [Google Scholar] [CrossRef]
- Plotnikov, E.Y.; Silachev, D.N.; Jankauskas, S.S.; Rokitskaya, T.I.; Chupyrkina, A.A.; Pevzner, I.B.; Zorova, L.D.; Isaev, N.K.; Antonenko, Y.N.; Skulachev, V.P.; et al. Mild uncoupling of respiration and phosphorylation as a mechanism providing nephro- and neuroprotective effects of penetrating cations of the SkQ family. Biochemistry 2012, 77, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Dalla Via, L.; García-Argáez, A.N.; Braga, A.; Martinez-Vazquez, M.; Grancara, S.; Martinis, P.; Agostinelli, E.; Toninello, A. An eudesman derivative from Verbesina persicifolia D.C. as a natural mild uncoupler in liver mitochondria. A new potential anti-obesity agent? Curr. Pharm. Des. 2014, 20, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Khailova, L.S.; Silachev, D.N.; Rokitskaya, T.I.; Armine, V.; Avetisyan, A.V.; Lyamsaev, K.G.; Severina, I.I.; Il′yasova, T.M.; Gulyaev, M.V.; Dedukhova, V.I.; et al. A short-chain alkyl derivative of Rhodamine 19 acts as a mild uncoupler of mitochondria and a neuroprotector. Biochim. Biophys. Acta 2014, 1837, 1739–1747. [Google Scholar] [CrossRef] [PubMed]
- Sztark, F.; Ouhabi, R.; Dabadie, P.; Mazat, J.P. Effects of the local anesthetic bupivacaine on mitochondrial energy metabolism: Change from uncoupling to decoupling depending on the respiration state. Biochem. Mol. Biol. Int. 1997, 43, 997–1003. [Google Scholar] [CrossRef]
- Cho, I.; Song, H.O.; Cho, J.H. Flavonoids mitigate neurodegeneration in aged Caenorhabditis elegans by mitochondrial uncoupling. Food Sci. Nutr. 2020, 8, 6633–6642. [Google Scholar] [CrossRef]
- Han, D.; Matsumaru, K.; Rettori, D.; Kaplowitz, N. Usnic acid-induced necrosis of cultured mouse hepatocytes: Inhibition of mitochondrial function and oxidative stress. Biochem. Pharmacol. 2004, 67, 439–451. [Google Scholar] [CrossRef]
- Kadri, H.; Lambourne, O.A.; Mehellou, Y. Niclosamide, a Drug with Many (Re)purposes. ChemMedChem 2018, 13, 1088–1091. [Google Scholar] [CrossRef]
- Tjahjono, E.; Pei, J.; Revtovich, A.V.; Liu, T.J.E.; Swadi, A.; Hancu, M.C.; Tolar, J.G.; Kirienko, N.V. Mitochondria-affecting small molecules ameliorate proteostasis defects associated with neurodegenerative diseases. Sci. Rep. 2021, 11, 17733. [Google Scholar] [CrossRef]
- Lim, H.W.; Lim, H.Y.; Wong, K.P. Uncoupling of oxidative phosphorylation by curcumin: Implication of its cellular mechanism of action. Biochem. Biophys. Res. Commun. 2009, 389, 187–192. [Google Scholar] [CrossRef]
- Jian, C.; Fu, J.; Cheng, X.; Shen, L.J.; Ji, Y.X.; Wang, X.; Pan, S.; Tian, H.; Tian, S.; Liao, R.; et al. Low-Dose Sorafenib Acts as a Mitochondrial Uncoupler and Ameliorates Nonalcoholic Steatohepatitis. Cell Metab. 2020, 31, 892–908.e11. [Google Scholar] [CrossRef] [PubMed]
- Figarola, J.L.; Singhal, J.; Tompkins, J.D.; Rogers, G.W.; Warden, C.; Horne, D.; Riggs, A.D.; Awasthi, S.; Singhal, S.S. SR4 Uncouples Mitochondrial Oxidative Phosphorylation, Modulates AMP-dependent Kinase (AMPK)-Mammalian Target of Rapamycin (mTOR) Signaling, and Inhibits Proliferation of HepG2 Hepatocarcinoma Cells. J. Biol. Chem. 2015, 290, 30321–30341. [Google Scholar] [CrossRef]
- Zhang, W.; Sviripa, V.M.; Kril, L.M.; Yu, T.; Xie, Y.; Hubbard, W.B.; Sullivan, P.G.; Chen, X.; Zhan, C.G.; Yang-Hartwich, Y.; et al. An Underlying Mechanism of Dual Wnt Inhibition and AMPK Activation: Mitochondrial Uncouplers Masquerading as Wnt Inhibitors. J. Med. Chem. 2019, 62, 11348–11358. [Google Scholar] [CrossRef] [PubMed]
- Yau, W.W.; Singh, B.K.; Lesmana, R.; Zhou, J.; Sinha, R.A.; Wong, K.A.; Wu, Y.; Bay, B.H.; Sugii, S.; Sunet, L.; et al. Thyroid hormone (T3) stimulates brown adipose tissue activation via mitochondrial biogenesis and MTOR-mediated mitophagy. Autophagy 2019, 15, 131–150. [Google Scholar] [CrossRef] [PubMed]
- Anisimov, V.N.; Bakeeva, L.E.; Egormin, P.A.; Filenko, O.F.; Isakova, E.F.; Manskikh, V.N.; Mikhelson, V.M.; Panteleeva, A.A.; Pasyukova, E.G.; Pilipenko, D.I.; et al. Mitochondria-targeted plastoquinone derivatives as tools to interrupt execution of the aging program. 5. SkQ1 prolongs lifespan and prevents development of traits of senescence. Biochemistry 2008, 73, 1329–1342. [Google Scholar] [CrossRef]
- Guo, J.; Tao, H.; Alasadi, A.; Huang, Q.; Jin, S. Niclosamide piperazine prevents high-fat diet-induced obesity and diabetic symptoms in mice. Eat. Weight. Disord. 2019, 24, 91–96. [Google Scholar] [CrossRef]
- Perry, R.J.; Kim, T.; Zhang, X.M.; Lee, H.Y.; Pesta, D.; Popov, V.B.; Zhang, D.; Rahimi, Y.; Jurczak, M.J.; Cline, G.W.; et al. Reversal of hypertriglyceridemia, fatty liver disease, and insulin resistance by a liver-targeted mitochondrial uncoupler. Cell Metab. 2013, 18, 740–748. [Google Scholar] [CrossRef]
- Geisler, J.G.; Marosi, K.; Halpern, J.; Mattson, M.P. DNP, mitochondrial uncoupling, and neuroprotection: A little dab′ll do ya. Alzheimers Dement. 2017, 13, 582–591. [Google Scholar] [CrossRef]
- Silachev, D.N.; Khailova, L.S.; Babenko, V.A.; Gulyaev, M.V.; Kovalchuk, S.I.; Zorova, L.D.; Plotnikov, E.Y.; Antonenko, Y.N.; Zorov, D.B. Neuroprotective effect of glutamate-substituted analog of gramicidin A is mediated by the uncoupling of mitochondria. Biochim. Biophys. Acta 2014, 1840, 3434–3442. [Google Scholar] [CrossRef]
- Gao, J.L.; Zhao, J.; Zhu, H.B.; Peng, X.; Zhu, J.X.; Ma, M.H.; Fu, Y.; Hu, N.; Tai, Y.; Xuan, X.C.; et al. Characterizations of mitochondrial uncoupling induced by chemical mitochondrial uncouplers in cardiomyocytes. Free Radic. Biol. Med. 2018, 124, 288–298. [Google Scholar] [CrossRef]
- Minners, J.; van den Bos, E.J.; Yellon, D.M.; Schwalb, H.; Opie, L.H.; Sack, M.N. Dinitrophenol, cyclosporin A, and trimetazidine modulate preconditioning in the isolated rat heart: Support for a mitochondrial role in cardioprotection. Cardiovasc. Res. 2000, 47, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Zhang, Y.; Zeng, X.; Shulman, G.I.; Jin, S. Niclosamide ethanolamine-induced mild mitochondrial uncoupling improves diabetic symptoms in mice. Nat. Med. 2014, 20, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Geisler, J.G. 2,4 Dinitrophenol as Medicine. Cells 2019, 8, 280. [Google Scholar] [CrossRef]
- Kalinovich, A.V.; Shabalina, I.G. Novel Mitochondrial Cationic Uncoupler C4R1 Is an Effective Treatment for Combating Obesity in Mice. Biochemistry 2015, 80, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Singhal, S.S.; Figarola, J.; Singhal, J.; Leake, K.; Nagaprashantha, L.; Lincoln, C.; Gugiu, B.G.; Horne, D.; Jove, R.; Awasthi, S.; et al. 1,3-Bis(3,5-dichlorophenyl) urea compound ′COH-SR4′ inhibits proliferation and activates apoptosis in melanoma. Biochem. Pharmacol. 2012, 84, 1419–1427. [Google Scholar] [CrossRef]
- Pearson, R.D.; Hewlett, E.L. Niclosamide therapy for tapeworm infections. Ann. Intern. Med. 1985, 102, 550–551. [Google Scholar] [CrossRef]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Conti, B.; Sugama, S.; Lucero, J.; Winsky-Sommerer, R.; Wirz, S.A.; Maher, P.; Andrews, Z.; Barr, A.M.; Morale, M.C.; Paneda, C.; et al. Uncoupling protein 2 protects dopaminergic neurons from acute 1,2,3,6-methyl-phenyl-tetrahydropyridine toxicity. J. Neurochem. 2005, 93, 493–501. [Google Scholar] [CrossRef]
- Bergman, J.; Botling, J.; Fagerberg, L.; Hallström, B.M.; Djureinovic, D.; Uhlén, M.; Pontén, F. The Human Adrenal Gland Proteome Defined by Transcriptomics and Antibody-Based Profiling. Endocrinology 2017, 158, 239–251. [Google Scholar] [CrossRef]
- Lim, J.; Park, H.S.; Kim, J.; Jang, Y.J.; Kim, J.-H.; Lee, Y.; Heo, Y. Depot-specific UCP1 expression in human white adipose tissue and its association with obesity-related markers. Int. J. Obes. 2020, 44, 697–706. [Google Scholar] [CrossRef]
- Poher, A.L.; Veyrat-Durebex, C.; Altirriba, J.; Montet, X.; Colin, D.J.; Caillon, A.; Lyautey, J.; Rohner-Jeanrenaud, F. Ectopic UCP1 Overexpression in White Adipose Tissue Improves Insulin Sensitivity in Lou/C Rats, a Model of Obesity Resistance. Diabetes 2015, 64, 3700–3712. [Google Scholar] [CrossRef]
- Couplan, E.; Gelly, C.; Goubern, M.; Fleury, C.; Quesson, B.; Silberberg, M.; Thiaudière, E.; Mateo, P.; Lonchampt, M.; Levens, N.; et al. High level of uncoupling protein 1 expression in muscle of transgenic mice selectively affects muscles at rest and decreases their IIb fiber content. J. Biol. Chem. 2002, 277, 43079–43088. [Google Scholar] [CrossRef] [PubMed]
- Hoerter, J.; Gonzalez-Barroso, M.D.; Couplan, E.; Mateo, P.; Gelly, C.; Cassard-Doulcier, A.-M.; Diolez, P.; Bouillaud, F. Mitochondrial uncoupling protein 1 expressed in the heart of transgenic mice protects against ischemic-reperfusion damage. Circulation 2004, 110, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Klaus, S.; Rudolph, B.; Dohrmann, C.; Wehr, R. Expression of uncoupling protein 1 in skeletal muscle decreases muscle energy efficiency and affects thermoregulation and substrate oxidation. Physiol. Genomics 2005, 21, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Keipert, S.; Klaus, S.; Heldmaier, G.; Jastroch, M. UCP1 ectopically expressed in murine muscle displays native function and mitigates mitochondrial superoxide production. Biochim. Biophys. Acta 2010, 1797, 324–330. [Google Scholar] [CrossRef]
- Andreyev, A.Y.; Kushnareva, Y.E.; Starkov, A.A. Mitochondrial metabolism of reactive oxygen species. Biochemistry 2005, 70, 200–214. [Google Scholar] [CrossRef] [PubMed]
- Jia, P.; Wu, X.; Pan, T.; Xu, S.; Hu, J.; Ding, X. Uncoupling protein 1 inhibits mitochondrial reactive oxygen species generation and alleviates acute kidney injury. EBioMedicine 2019, 49, 331–340. [Google Scholar] [CrossRef]
- Fridell, Y.W.; Sánchez-Blanco, A.; Silvia, B.A.; Helfand, S.L. Targeted expression of the human uncoupling protein 2 (hUCP2) to adult neurons extends life span in the fly. Cell Metab. 2005, 1, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Islam, R.; Yang, L.; Sah, M.; Kannan, K.; Anamani, D.; Vijayan, C.; Kwok, J.; Cantino, M.E.; Beal, M.F.; Fridell, Y.-W.C. A neuroprotective role of the human uncoupling protein 2 (hUCP2) in a Drosophila Parkinson′s disease model. Neurobiol. Dis. 2012, 46, 137–146. [Google Scholar] [CrossRef]
- Andrews, Z.B.; Horvath, T.L. Uncoupling protein-2 regulates lifespan in mice. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E621–E627. [Google Scholar] [CrossRef]
- Peixoto, P.M.; Kim, H.J.; Sider, B.; Starkov, A.; Horvath, T.L.; Manfredi, G. UCP2 overexpression worsens mitochondrial dysfunction and accelerates disease progression in a mouse model of amyotrophic lateral sclerosis. Mol. Cell Neurosci. 2013, 57, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Deierborg, T.; Wieloch, T.; Diano, S.; Warden, C.H.; Horvath, T.L.; Mattiasson, G. Overexpression of UCP2 protects thalamic neurons following global ischemia in the mouse. J. Cereb. Blood Flow Metab. 2008, 28, 1186–1195. [Google Scholar] [CrossRef] [PubMed]
- Haines, B.; Li, P.A. Overexpression of mitochondrial uncoupling protein 2 inhibits inflammatory cytokines and activates cell survival factors after cerebral ischemia. PLoS ONE 2012, 7, e31739. [Google Scholar] [CrossRef]
- Mattiasson, G.; Shamloo, M.; Gido, G.; Mathi, K.; Tomasevic, G.; Yi, S.; Warden, C.H.; Castilho, R.F.; Melcher, T.; Gonzalez-Zulueta, M.; et al. Uncoupling protein-2 prevents neuronal death and diminishes brain dysfunction after stroke and brain trauma. Nat. Med. 2003, 9, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Hass, D.T.; Barnstable, C.J. Cell Autonomous Neuroprotection by the Mitochondrial Uncoupling Protein 2 in a Mouse Model of Glaucoma. Front. Neurosci. 2019, 13, 201. [Google Scholar] [CrossRef]
- Diano, S.; Matthews, R.T.; Patrylo, P.; Yang, L.; Beal, M.F.; Barnstable, C.J.; Horvath, T.L. Uncoupling protein 2 prevents neuronal death including that occurring during seizures: A mechanism for preconditioning. Endocrinology 2003, 144, 5014–5021. [Google Scholar] [CrossRef]
- Shang, Y.; Liu, Y.; Du, L.; Wang, Y.; Cheng, X.; Xiao, W.; Wang, X.; Jin, H.; Yang, X.; Liu, S.; et al. Targeted expression of uncoupling protein 2 to mouse liver increases the susceptibility to lipopolysaccharide/galactosamine-induced acute liver injury. Hepatology 2009, 50, 1204–1216. [Google Scholar] [CrossRef]
- Li, H.; Wang, C.; Li, L.; Li, L. Skeletal muscle non-shivering thermogenesis as an attractive strategy to combat obesity. Life Sci. 2021, 269, 119024. [Google Scholar] [CrossRef]
- Moore, G.B.; Himms-Hagen, J.; Harper, M.E.; Clapham, J.C. Overexpression of UCP-3 in skeletal muscle of mice results in increased expression of mitochondrial thioesterase mRNA. Biochem. Biophys. Res. Commun. 2001, 283, 785–790. [Google Scholar] [CrossRef]
- Codella, R.; Alves, T.C.; Befroy, D.E.; Choi, C.S.; Luzi, L.; Rothman, D.L.; Kibbey, R.G.; Shulman, G.I. Overexpression of UCP3 decreases mitochondrial efficiency in mouse skeletal muscle in vivo. FEBS Lett. 2023, 597, 309–319. [Google Scholar] [CrossRef]
- Kim-Han, J.S.; Reichert, S.A.; Quick, K.L.; Dugan, L.L. BMCP1: A mitochondrial uncoupling protein in neurons which regulates mitochondrial function and oxidant production. J. Neurochem. 2001, 79, 658–668. [Google Scholar] [CrossRef]
- Kwok, K.H.; Ho, P.W.; Chu, A.C.; Ho, J.W.-M.; Liu, H.-F.; Yiu, D.C.-W.; Chan, K.-H.; Kung, M.H.-W.; Ramsden, D.B.; Ho, S.-L. Mitochondrial UCP5 is neuroprotective by preserving mitochondrial membrane potential, ATP levels, and reducing oxidative stress in MPP+ and dopamine toxicity. Free Radic. Biol. Med. 2010, 49, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Rupprecht, A.; Bräuer, A.U.; Smorodchenko, A.; Goyn, J.; Hilse, K.E.; Shabalina, I.G.; Infante-Duarte, C.; Pohl, E.E. Quantification of uncoupling protein 2 reveals its main expression in immune cells and selective up-regulation during T-cell proliferation. PLoS ONE 2012, 7, e41406. [Google Scholar] [CrossRef] [PubMed]
- Horvath, T.L.; Warden, C.H.; Hajos, M.; Lombardi, A.; Goglia, F.; Diano, S. Brain uncoupling protein 2: Uncoupled neuronal mitochondria predict thermal synapses in homeostatic centers. J. Neurosci. 1999, 19, 10417–10427. [Google Scholar] [CrossRef] [PubMed]
- Busceti, C.L.; Cotugno, M.; Bianchi, F.; Forte, M.; Stanzione, R.; Marchitti, S.; Battaglia, G.; Nicoletti, F.; Fornai, F.; Rubattu, S. Brain Overexpression of Uncoupling Protein-2 (UCP2) Delays Renal Damage and Stroke Occurrence in Stroke-Prone Spontaneously Hypertensive Rats. Int. J. Mol. Sci. 2020, 21, 4289. [Google Scholar] [CrossRef] [PubMed]
- Clapham, J.C.; Arch, J.R.; Chapman, H.; Haynes, A.; Lister, C.; Moore, G.B.T.; Piercy, V.; Carter, S.A.; Lehner, I.; Smith, S.A.; et al. Mice overexpressing human uncoupling protein-3 in skeletal muscle are hyperphagic and lean. Nature 2000, 406, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Klotzsch, E.; Smorodchenko, A.; Löfler, L.; Moldzio, R.; Parkinson, E.; Schütz, G.J.; Pohl, E.E. Superresolution microscopy reveals spatial separation of UCP4 and F0F1-ATP synthase in neuronal mitochondria. Proc. Natl. Acad. Sci. USA 2015, 112, 130–135. [Google Scholar] [CrossRef]
- Rosenberg, N.; Reva, M.; Binda, F.; Restivo, L.; Depierre, P.; Puyal, J.; Briquet, M.; Bernardinelli, Y.; Rocher, A.; Markram, H.; et al. Overexpression of UCP4 in astrocytic mitochondria prevents multilevel dysfunctions in a mouse model of Alzheimer’s disease. Glia 2023, 71, 957–973. [Google Scholar] [CrossRef]
- Perreten Lambert, H.; Zenger, M.; Azarias, G.; Chatton, J.Y.; Magistretti, P.J.; Lengacher, S. Control of mitochondrial pH by uncoupling protein 4 in astrocytes promotes neuronal survival. J. Biol. Chem. 2014, 289, 31014–31028. [Google Scholar] [CrossRef]
- Dassa, E.P.; Dufour, E.; Gonçalves, S.; Paupe, V.; Hakkaart, G.A.; Jacobs, H.T.; Rustin, P. Expression of the alternative oxidase complements cytochrome c oxidase deficiency in human cells. EMBO Mol. Med. 2009, 1, 30–36. [Google Scholar] [CrossRef]
- El-Khoury, R.; Dufour, E.; Rak, M.; Ramanantsoa, N.; Grandchamp, N.; Csaba, Z.; Duvillié, B.; Bénit, P.; Gallego, J.; Gressens, P.; et al. Alternative oxidase expression in the mouse enables bypassing cytochrome c oxidase blockade and limits mitochondrial ROS overproduction. PLoS Genet. 2013, 9, e1003182. [Google Scholar] [CrossRef]
- El-Khoury, R.; Kaulio, E.; Lassila, K.A.; Crowther, D.C.; Jacobs, H.T.; Rustin, P. Expression of the alternative oxidase mitigates beta-amyloid production and toxicity in model systems. Free Radic. Biol. Med. 2016, 96, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Ayala, D.J.; Sanz, A.; Vartiainen, S.; Kemppainen, K.K.; Babusiak, M.; Mustalahti, E.; Costa, R.; Tuomela, T.; Zeviani, M.; Chung, J.; et al. Expression of the Ciona intestinalis alternative oxidase (AOX) in Drosophila complements defects in mitochondrial oxidative phosphorylation. Cell Metab. 2009, 9, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Giordano, L.; Farnham, A.; Dhandapani, P.K.; Salminen, L.; Bhaskaran, J.; Voswinckel, R.; Rauschkolb, P.; Scheibe, S.; Sommer, N.; Beisswenger, C.; et al. Alternative Oxidase Attenuates Cigarette Smoke-induced Lung Dysfunction and Tissue Damage. Am. J. Respir. Cell Mol. Biol. 2019, 60, 515–522. [Google Scholar] [CrossRef]
- Hakkaart, G.A.; Dassa, E.P.; Jacobs, H.T.; Rustin, P. Allotopic expression of a mitochondrial alternative oxidase confers cyanide resistance to human cell respiration. EMBO Rep. 2006, 7, 341–345. [Google Scholar] [CrossRef]
- Rajendran, J.; Purhonen, J.; Tegelberg, S.; Smolander, O.P.; Mörgelin, M.; Rozman, J.; Gailus-Durner, V.; Fuchs, H.; de Angelis, M.H.; Auvinen, P.; et al. Alternative oxidase-mediated respiration prevents lethal mitochondrial cardiomyopathy. EMBO Mol. Med. 2019, 11, e9456. [Google Scholar] [CrossRef]
- Szibor, M.; Dhandapani, P.K.; Dufour, E.; Holmström, K.M.; Zhuang, Y.; Salwig, I.; Wittig, I.; Heidler, J.; Gizatullina, Z.; Gainutdinov, T.; et al. Broad AOX expression in a genetically tractable mouse model does not disturb normal physiology. Dis. Model. Mech. 2017, 10, 163–171. [Google Scholar] [CrossRef]
- Gospodaryov, D.V.; Lushchak, O.V.; Rovenko, B.M.; Perkhulyn, N.V.; Gerards, M.; Tuomela, T.; Jacobs, H.T. Ciona intestinalis NADH dehydrogenase NDX confers stress-resistance and extended lifespan on Drosophila. Biochim. Biophys. Acta 2014, 1837, 1861–1869. [Google Scholar] [CrossRef] [PubMed]
- Gospodaryov, D.V.; Strilbytska, O.M.; Semaniuk, U.V.; Perkhulyn, N.V.; Rovenko, B.M.; Yurkevych, I.S.; Barata, A.G.; Dick, T.P.; Lushchak, O.V.; Jacobs, H.T. Alternative NADH dehydrogenase extends lifespan and increases resistance to xenobiotics in Drosophila. Biogerontology 2020, 21, 155–171. [Google Scholar] [CrossRef]
- Bahadorani, S.; Cho, J.; Lo, T.; Contreras, H.; Lawal, H.O.; Krantz, D.E.; Bradley, T.J.; Walker, D.W. Neuronal expression of a single-subunit yeast NADH-ubiquinone oxidoreductase (Ndi1) extends Drosophila lifespan. Aging Cell 2010, 9, 191–202. [Google Scholar] [CrossRef]
- Cho, J.; Hur, J.H.; Graniel, J.; Benzer, S.; Walker, D.W. Expression of yeast NDI1 rescues a Drosophila complex I assembly defect. PLoS ONE 2012, 7, e50644. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.H.; Bahadorani, S.; Graniel, J.; Koehler, C.L.; Ulgherait, M.; Rera, M.; Jones, D.L.; Walker, D.W. Increased longevity mediated by yeast NDI1 expression in Drosophila intestinal stem and progenitor cells. Aging 2013, 5, 662–681. [Google Scholar] [CrossRef] [PubMed]
- Sanz, A.; Soikkeli, M.; Portero-Otín, M.; Wilson, A.; Kemppainen, E.; McIlroy, G.; Ellilä, S.; Kemppainen, K.K.; Tuomela, T.; Lakanmaa, M.; et al. Expression of the yeast NADH dehydrogenase Ndi1 in Drosophila confers increased lifespan independently of dietary restriction. Proc. Natl. Acad. Sci. USA 2010, 107, 9105–9110. [Google Scholar] [CrossRef]
- Catania, A.; Iuso, A.; Bouchereau, J.; Kremer, L.S.; Paviolo, M.; Terrile, C.; Bénit, P.; Rasmusson, A.G.; Schwarzmayr, T.; Tiranti, V.; et al. Arabidopsis thaliana alternative dehydrogenases: A potential therapy for mitochondrial complex I deficiency? Perspectives and pitfalls. Orphanet J. Rare Dis. 2019, 14, 236. [Google Scholar] [CrossRef] [PubMed]
- Kemppainen, K.K.; Rinne, J.; Sriram, A.; Lakanmaa, M.; Zeb, A.; Tuomela, T.; Popplestone, A.; Singh, S.; Sanz, A.; Rustin, P.; et al. Expression of alternative oxidase in Drosophila ameliorates diverse phenotypes due to cytochrome oxidase deficiency. Hum. Mol. Genet. 2014, 23, 2078–2093. [Google Scholar] [CrossRef]
- Toivonen, J.M.; O′Dell, K.M.; Petit, N.; Irvine, S.C.; Knight, G.K.; Lehtonen, M.; Longmuir, M.; Luoto, K.; Touraille, S.; Wang, Z.; et al. Technical knockout, a Drosophila model of mitochondrial deafness. Genetics 2001, 159, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Kemppainen, K.K.; Kemppainen, E.; Jacobs, H.T. The alternative oxidase AOX does not rescue the phenotype of tko25t mutant flies. G3 Genes Genomes Genet. 2014, 4, 2013–2021. [Google Scholar] [CrossRef]
- Dhandapani, P.K.; Begines-Moreno, I.M.; Brea-Calvo, G.; Gärtner, U.; Graeber, T.G.; Sanchez, G.J.; Morty, R.E.; Schönig, K.; Hoeve, J.T.; Wietelmann, A.; et al. Hyperoxia but not AOX expression mitigates pathological cardiac remodeling in a mouse model of inflammatory cardiomyopathy. Sci. Rep. 2019, 9, 12741. [Google Scholar] [CrossRef]
- Gureev, A.P.; Sadovnikova, I.S.; Popov, V.N. Molecular Mechanisms of the Neuroprotective Effect of Methylene Blue. Biochemistry 2022, 87, 940–956. [Google Scholar] [CrossRef]
- Tretter, L.; Horvath, G.; Hölgyesi, A.; Essek, F.; Adam-Vizi, V. Enhanced hydrogen peroxide generation accompanies the beneficial bioenergetic effects of methylene blue in isolated brain mitochondria. Free Radic. Biol. Med. 2014, 77, 317–330. [Google Scholar] [CrossRef]
- Gureev, A.P.; Shaforostova, E.A.; Popov, V.N.; Starkov, A.A. Methylene blue does not bypass Complex III antimycin block in mouse brain mitochondria. FEBS Lett. 2019, 593, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Bariotto-Dos-Santos, K.; Padovan-Neto, F.E.; Bortolanza, M.; Dos-Santos-Pereira, M.; Raisman-Vozari, R.; Tumas, V.; Del Bel, E. Repurposing an established drug: An emerging role for methylene blue in L-DOPA-induced dyskinesia. Eur. J. Neurosci. 2019, 49, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Heidari, R.; Monnier, V.; Martin, E.; Tricoire, H. Methylene Blue Partially Rescues Heart Defects in a Drosophila Model of Huntington′s Disease. J. Huntingtons Dis. 2015, 4, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Dibaj, P.; Zschüntzsch, J.; Steffens, H.; Scheffel, J.; Göricke, B.; Weishaupt, J.H.; Le Meur, K.; Kirchhoff, F.; Hanisch, U.-K.; Schomburg, E.D.; et al. Influence of methylene blue on microglia-induced inflammation and motor neuron degeneration in the SOD1(G93A) model for ALS. PLoS ONE 2012, 7, e43963. [Google Scholar] [CrossRef] [PubMed]
- Sadovnikova, I.S.; Gureev, A.P.; Ignatyeva, D.A.; Gryaznova, M.V.; Chernyshova, E.V.; Krutskikh, E.P.; Novikova, A.G.; Popov, V.N. Nrf2/ARE Activators Improve Memory in Aged Mice via Maintaining of Mitochondrial Quality Control of Brain and the Modulation of Gut Microbiome. Pharmaceuticals 2021, 14, 607. [Google Scholar] [CrossRef]
- Alasadi, A.; Chen, M.; Swapna, G.V.T.; Tao, H.; Guo, J.; Collantes, J.; Jin, S. Effect of mitochondrial uncouplers niclosamide ethanolamine (NEN) and oxyclozanide on hepatic metastasis of colon cancer. Cell Death Dis. 2018, 9, 215. [Google Scholar] [CrossRef]
- Alexopoulos, S.J.; Chen, S.Y.; Brandon, A.E.; Salamoun, J.M.; Byrne, F.L.; Garcia, C.J.; Hoehn, K.L. Mitochondrial uncoupler BAM15 reverses diet-induced obesity and insulin resistance in mice. Nat. Commun. 2020, 11, 2397. [Google Scholar] [CrossRef]
- Amireddy, N.; Puttapaka, S.N.; Vinnakota, R.L.; Ravuri, H.G.; Thonda, S.; Kalivendi, S.V. The unintended mitochondrial uncoupling effects of the FDA-approved anti-helminth drug nitazoxanide mitigates experimental parkinsonism in mice. J. Biol. Chem. 2021, 292, 15731–15743, Corrected in J. Biol. Chem. 2021, 297, 100864.. [Google Scholar] [CrossRef]
- Chen, Y.; Du, F.; Tang, L.; Xu, J.; Zhao, Y.; Wu, X.; Xiao, Z. Carboranes as unique pharmacophores in antitumor medicinal chemistry. Mol. Ther. Oncolytics. 2022, 24, 400–416. [Google Scholar] [CrossRef]
- Dabadie, P.; Bendriss, P.; Erny, P.; Mazat, J.P. Uncoupling effects of local anesthetics on rat liver mitochondria. FEBS Lett. 1987, 226, 77–82. [Google Scholar] [CrossRef]
- Fu, Y.Y.; Zhang, M.; Turner, N.; Zhang, L.N.; Dong, T.C.; Gu, M.; Li, J. A novel chemical uncoupler ameliorates obesity and related phenotypes in mice with diet-induced obesity by modulating energy expenditure and food intake. Diabetologia. 2013, 56, 2297–2307. [Google Scholar] [CrossRef]
- Guimarães, E.L.; Best, J.; Dollé, L.; Najimi, M.; Sokal, E.; van Grunsven, L.A. Mitochondrial uncouplers inhibit hepatic stellate cell activation. BMC Gastroenterol. 2012, 12, 68. [Google Scholar] [CrossRef] [PubMed]
- Hägg, M.; Berndtsson, M.; Mandic, A.; Zhou, R.; Shoshan, M.C.; Linder, S. Induction of endoplasmic reticulum stress by ellipticine plant alkaloids. Mol. Cancer Ther. 2004, 3, 489–497. [Google Scholar] [CrossRef]
- Holtrup, F.; Bauer, A.; Fellenberg, K.; Hilger, R.A.; Wink, M.; Hoheisel, J.D. Microarray analysis of nemorosone-induced cytotoxic effects on pancreatic cancer cells reveals activation of the unfolded protein response (UPR). Br. J. Pharmacol. 2011, 162, 1045–1059. [Google Scholar] [CrossRef] [PubMed]
- Ilivicky, J.; Casida, J.E. Uncoupling action of 2,4-dinitrophenols, 2-trifluoromethylbenzimidazoles and certain other pesticide chemicals upon mitochondria from different sources and its relation to toxicity. Biochem. Pharmacol. 1969, 18, 1389–1401. [Google Scholar] [CrossRef] [PubMed]
- Kanemoto, N.; Okamoto, T.; Tanabe, K.; Shimada, T.; Minoshima, H.; Hidoh, Y.; Sato, S. Antidiabetic and cardiovascular beneficial effects of a liver-localized mitochondrial uncoupler. Nat. Commun. 2019, 10, 2172. [Google Scholar] [CrossRef] [PubMed]
- Nie, L.; Yuan, X.L.; Jiang, K.T.; Jiang, Y.H.; Yuan, J.; Luo, L.; Sun, C. Salsalate Activates Skeletal Muscle Thermogenesis and Protects Mice from High-Fat Diet Induced Metabolic Dysfunction. EBioMedicine. 2017, 23, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Pardo-Andreu, G.L.; Nuñez-Figueredo, Y.; Tudella, V.G.; Cuesta-Rubio, O.; Rodrigues, F.P.; Pestana, C.R.; Curti, C. The anti-cancer agent guttiferone-A permeabilizes mitochondrial membrane: Ensuing energetic and oxidative stress implications. Toxicol. Appl. Pharmacol. 2011, 253, 282–289. [Google Scholar] [CrossRef]
- Rawling, T.; MacDermott-Opeskin, H.; Roseblade, A.; Pazderka, C.; Clarke, C.; Bourget, K.; Murray, M. Aryl urea substituted fatty acids: A new class of protonophoric mitochondrial uncoupler that utilises a synthetic anion transporter. Chem. Sci. 2020, 11, 12677–12685. [Google Scholar] [CrossRef]
- Rokitskaya, T.I.; Khailova, L.S.; Makarenkov, A.V.; Ol'shevskaya, V.A.; Kalinin, V.N.; Antonenko, Y.N. Weak C-H acids as protonophores can carry hydrogen ions through lipid membranes and mitochondria: A case of o-carborane. Phys Chem Chem Phys. 2016, 18, 16476–16482. [Google Scholar] [CrossRef]
- Sagara, Y.; Ishige, K.; Tsai, C.; Maher, P. Tyrphostins protect neuronal cells from oxidative stress. J. Biol. Chem. 2002, 277, 36204–36215. [Google Scholar] [CrossRef] [PubMed]
- Thomason, L.C.; Court, D.L. Evidence that bacteriophage λ lysogens may induce in response to the proton motive force uncoupler CCCP. FEMS Microbiol. Lett. 2016, 363, fnv244. [Google Scholar] [CrossRef] [PubMed]
- Williamson, R.L.; Metcalf, R.L. Salicylanilides: A new group of active uncouplers of oxidative phosphorylation. Science. 1967, 158, 1694–1695. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; He, H.; Xiang, C.; Fan, X.Y.; Yang, L.Y.; Yuan, L.; Jiang, F.L.; Liu, Y. Uncoupling Effect of F16 Is Responsible for Its Mitochondrial Toxicity and Anticancer Activity. Toxicol. Sci. 2018, 161, 431–442. [Google Scholar] [CrossRef]
- Marín-Prida, J.; Pardo Andreu, G.L.; Rossignoli, C.P.; Durruthy, M.G.; Rodríguez, E.O.; Reyes, Y.V.; Acosta, R.F.; Uyemura, S.A.; Alberici, L.C. The cytotoxic effects of VE-3N, a novel 1,4-dihydropyridine derivative, involve the mitochondrial bioenergetic disruption via uncoupling mechanisms. Toxicol. Vitr. 2017, 42, 21–30. [Google Scholar] [CrossRef]
- Dejonghe, W.; Kuenen, S.; Mylle, E.; Vasileva, M.; Keech, O.; Viotti, C.; Swerts, J.; Fendrych, M.; Ortiz-Morea, F.A.; Mishev, K.; et al. Mitochondrial uncouplers inhibit clathrin-mediated endocytosis largely through cytoplasmic acidification. Nat. Commun. 2016, 7, 11710. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overexpressed Gene | Transgenic Strains | Model of Disease | Effect | Reference |
|---|---|---|---|---|
| UCP1 | Transgenic strains expressing UCP1 in muscle and heart | Body weight was reduced with the same food intake. The decrease in weight mainly occurred in muscle tissue. No changes were observed in cardiac muscle | [64] | |
| UCP1 | Transgenic strains expressing UCP1 in muscle and heart | Heart ischemia/reperfusion | Functional recovery on reperfusion was improved | [65] |
| UCP1 | HSA-mUCP1 mice expressing UCP1 in the skeletal muscles | RQ level was increased, indicating an overall increase in glucose oxidation | [66] | |
| UCP1 | HSA-mUCP1 mice expressing UCP1 in the skeletal muscles | UCP1 expression in skeletal muscle reduced the risk of reverse electron transfer and the production of reactive oxygen species | [67] | |
| UCP1 | Lou/C rats with UCP1 overexpression in WAT | Obesity | Prevented body weight gain, decreased fat mass, and improved insulin sensitivity | [63] |
| UCP2 | Transgenic fly, UAS-hUCP2 | Increased hUCP2 expression in the adult nervous system, extended life span | [70] | |
| UCP2 | Transgenic fly, UAS-hUCP2 | PD model | Less ROS accumulation, heightened resistance to rotenone-induced lethality, and extended life span | [71] |
| UCP2 | Transgenic mice overexpressing UCP2 in catecholaminergic neurons (TH-UCP2) | PD model | Upon acute exposure to MPTP, TH-UCP2 mice showed neuroprotection and retention of locomotor functions | [60] |
| UCP2 | Transgenic mice overexpressing hUCP2 | ALS model | Increased survival of sod2−/− mice | [72] |
| UCP2 | Transgenic mice overexpressing hUCP2 | ALS model | Worsened mitochondrial dysfunction and accelerated disease progression of sod2−/− mice | [73] |
| UCP2 | UCP2/3 transgenic overexpressing mice | Global ischemia | Overexpression of UCP2 protects thalamic neurons following global ischemia | [74] |
| UCP2 | UCP2/3 transgenic overexpressing mice | Focal ischemia | Overexpression of UCP2 blunted the ischemia-induced increase in IL-6 and decrease in Bcl2. | [75] |
| UCP2 | UCP2/3 transgenic overexpressing mice | Stroke TBI | Overexpression of UCP2 enhased of neurological recovery | [76] |
| UCP2 | Ucp2KIfl/fl mice | Glaucoma | Decreased glaucomatous cell death | [77] |
| UCP2 | UCP2/3-overexpressing mice | Epileptic seizures | Increased mitochondrial number and ATP levels with a parallel decrease in free radical-induced damage | [78] |
| UCP2 | Transgenic mice with targeted expression of UCP2 in the liver | Acute liver injury | Expression of UCP2 in mouse liver increases susceptibility to acute liver injury induced by lipopolysaccharide and galactosamine | [79] |
| UCP3 | Mice overexpressing human UCP-3 in skeletal muscle (UCP-3tg) | Mice were hyperphagic but weighed less | [80] | |
| UCP3 | Mice overexpressing human UCP-3 in skeletal muscle (UCP-3tg) | UCP-3tg showed increase in β-oxidation in the MTE-1-dependent manner | [81] | |
| UCP3 | Mice overexpressing human UCP-3 in skeletal muscle (UCP-3tg) | UCP-3tg showed increase in muscle mitochondrial inefficiency and decrease in ATP synthesis | [82] | |
| UCP5 | UCP5-transfected cell lines of heart and kidney | The mitochondrial ROS production was decreased | [83] | |
| UCP5 | SH-SY5Y neuroblastoma cells stably overexpressing human UCP5 | PD model | UCP5 overexpression protected against MPP(+)- and dopamine-induced toxicity | [84] |
| Model Object | Gene | Donor Organism | Complex Bypass | ROS Effect | Model of Disease | Reference |
|---|---|---|---|---|---|---|
| Cell lines | ||||||
| Flp-InTM T-RExTM-293 cells | AOX | C. intestinalis | CIV dysfunction. Cyanide-induced inhibition | Decrease in antimycin-induced superoxide overproduction | [97] | |
| COX10-depleted HEK-293-derived AOX-transgenic cells from Hakkaart et al., 2005 | AOX | C. intestinalis | CIV dysfunction. shRNA targeted against COX10 | [92] | ||
| COX-defective fibroblasts | AOX | C. intestinalis | CIV dysfunction. Deleterious COX15 gene mutation | Reduction in the superoxide production in COX15– cells in the presence of antimycin | Hypertrophic cardiomyopathy | [92] |
| HEK293 Flp-In cells | AOX | C. intestinalis | Alzheimer’s disease | [94] | ||
| Complex I defective fibroblasts | NDH-2 | Arabidopsis thaliana | CI dysfunction. CI-defective fibroblasts | The normalization of SOD activity | [106] | |
| Drosophila | ||||||
| w1118 Drosophila | AOX | C. intestinalis | CIV dysfunctions. Partial knockdown of COXVIc and complex IV assembly factor Surf1 | Leigh syndrome | [95] | |
| w1118 Drosophila | AOX | C. intestinalis | Reduction in the ROS production | Parkinson’s disease | [95] | |
| w1118 Drosophila with knockdown of different complex IV subunits | AOX | C. intestinalis | CIV dysfunctions. Knockdown of different CIV subunits | |||
| w1118 Drosophila | AOX | C. intestinalis | AOX abrogates induction of oxidative stress markers in a Drosophila AD model | Alzheimer’s disease | [94] | |
| w1118 Drosophila | Ndi1 | S. cerevisiae | CI dysfunction. Rotenone-induced inhibition, paraquat-induced inhibition | Mitigation of mitochondrial ROS production, oxidative damage | [105] | |
| w1118 Drosophila | Ndi1 | S. cerevisiae | CI dysfunction. Rotenone-induced inhibition | Ndi1 expression in neurons, reducing ROS levels | [102] | |
| UAS-dCIA30 Drosophila | Ndi1 | S. cerevisiae | CI dysfunction. Reduced expression of CI assembly factor | [103] | ||
| w1118 Drosophila | Ndi1 | Saccharomyces cerevisiae | CI dysfunction. Rotenone-induced inhibition | Reduction in whole tissue ROS levels | [104] | |
| Drosophila lines 24,861 and 24,871 | Ndx | C. intestinalis | Increased resistance to menadione-induced ROS production | [100] | ||
| Mice | ||||||
| Mice (strain not specified) | AOX | C. intestinalis | CIV dysfunction. Cyanide-induced inhibition | Decrease in antimycin- and cyanide-induced superoxide overproduction | [93] | |
| AOXRosa26 mice, C57Bl/6J strain background | AOX | C. intestinalis | CIV dysfunction. Cyanide-induced inhibition Azide-induced inhibition. CIII dysfunction. Antimicyn-induced inhibition | Decrease in H2O2 production in succinate-supported mitochondria | [99] | |
| AOXRosa26 mice, C57Bl/6J strain background | AOX | C. intestinalis | CII dysfunction. Cigarette smoke condensate-induced inhibition. CIV dysfunction. Cigarette smoke condensate-induced inhibition | Decreases superoxide production | [96] | |
| (cIII)-deficient Bcs1lp.S78G knock-in mice AOX backcross with transgenic mice | AOX | C. intestinalis | CIII dysfunction. (CIII)-deficient Bcs1lp.S78G knock-in mice | Lethal mitochondrial cardiomyopathy | [98] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gureev, A.P.; Alimova, A.A.; Silachev, D.N.; Plotnikov, E.Y. Noncoupled Mitochondrial Respiration as Therapeutic Approach for the Treatment of Metabolic Diseases: Focus on Transgenic Animal Models. Int. J. Mol. Sci. 2023, 24, 16491. https://doi.org/10.3390/ijms242216491
Gureev AP, Alimova AA, Silachev DN, Plotnikov EY. Noncoupled Mitochondrial Respiration as Therapeutic Approach for the Treatment of Metabolic Diseases: Focus on Transgenic Animal Models. International Journal of Molecular Sciences. 2023; 24(22):16491. https://doi.org/10.3390/ijms242216491
Chicago/Turabian StyleGureev, Artem P., Alina A. Alimova, Denis N. Silachev, and Egor Y. Plotnikov. 2023. "Noncoupled Mitochondrial Respiration as Therapeutic Approach for the Treatment of Metabolic Diseases: Focus on Transgenic Animal Models" International Journal of Molecular Sciences 24, no. 22: 16491. https://doi.org/10.3390/ijms242216491