
Sex Drives Functional Changes in the Progression and Regression of Liver Fibrosis
 , , and
, , and
Abstract
:1. Introduction
2. Results
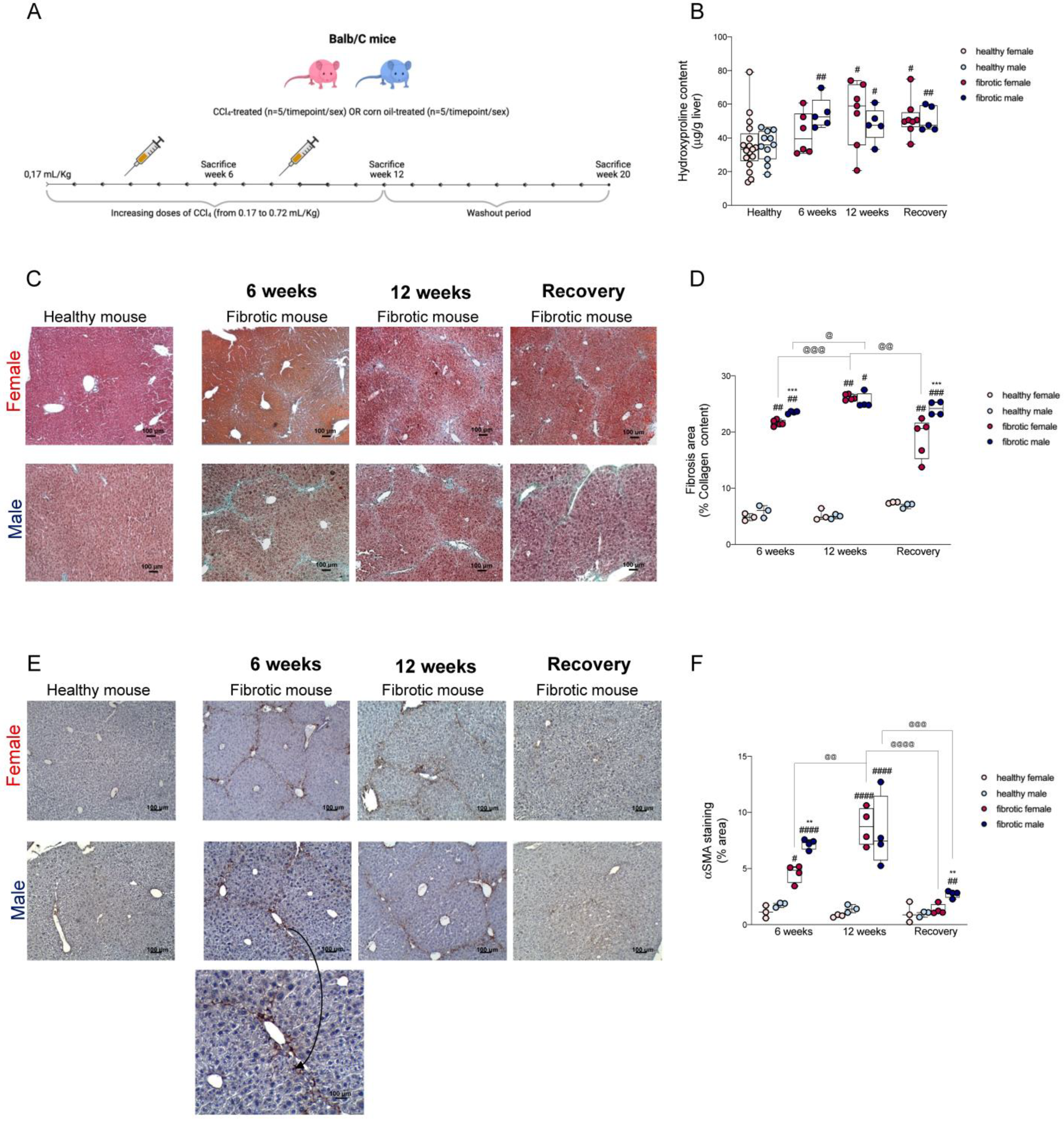
2.1. The Progression of Fibrosis and Regeneration Differed between Males and Females Due to the Differences in the Activation of Hepatic Stellate Cells
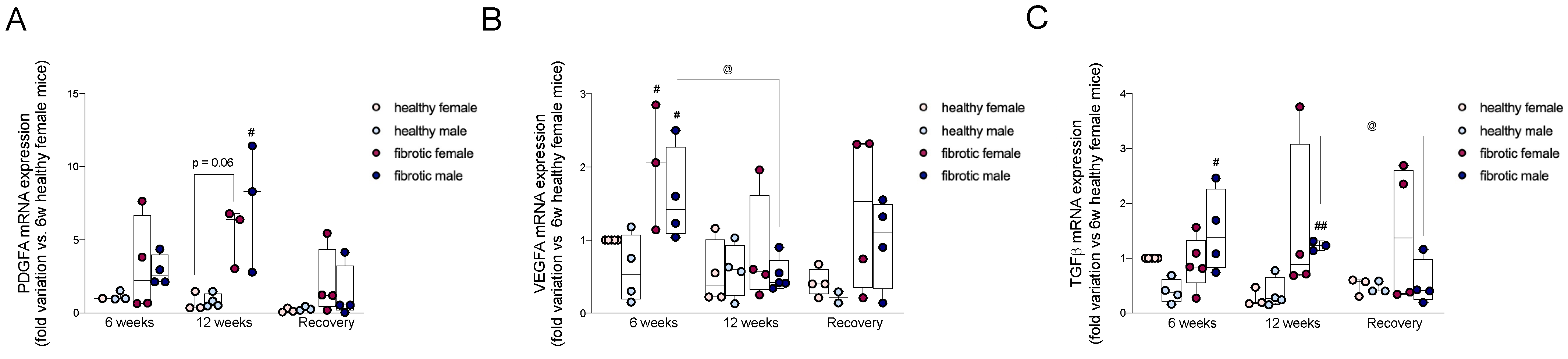
2.2. Pro-Fibrogenic TGF-β, VEGF-A, and PDGF-A Showed Different Patterns of Expression during Different Stages of Liver Damage and Regeneration
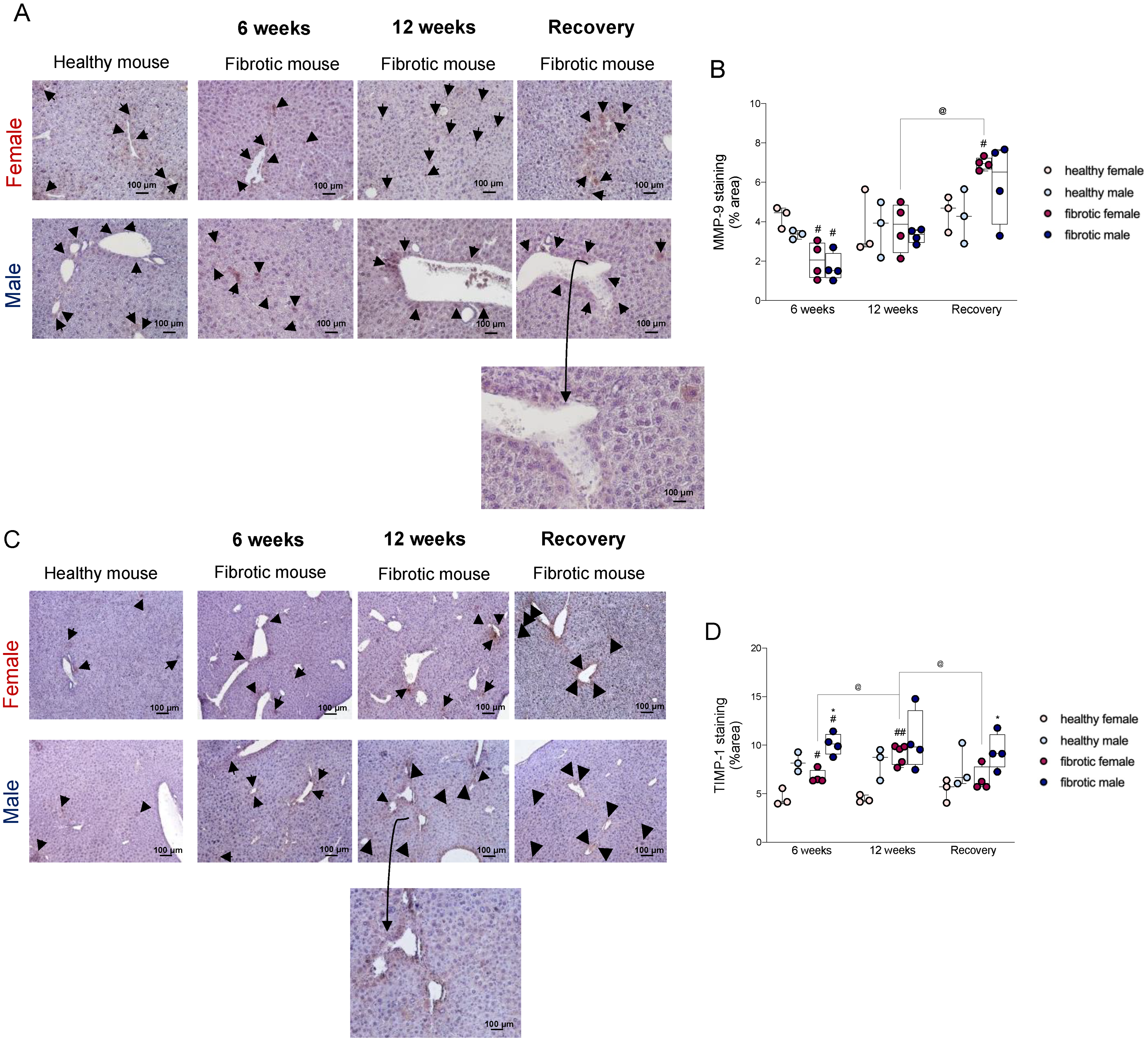
2.3. Sex Influenced the Disequilibrium of the Expression of the MMP9-TIMP1 Protein
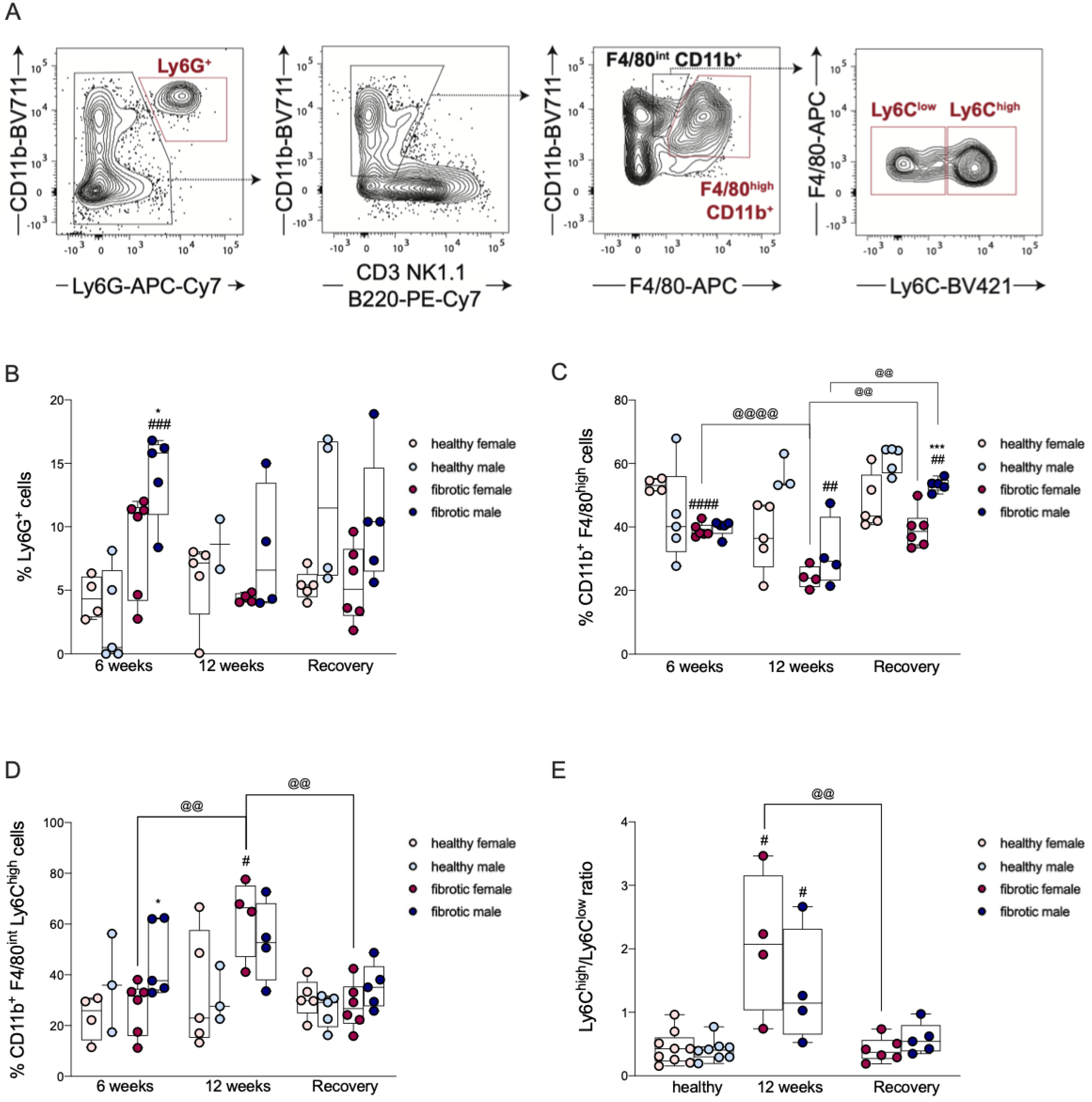
2.4. The Infiltration of Neutrophils Was Driven by Sex-Dependent Mechanisms
2.5. Transient Reduction in Liver-Resident Macrophages Was Associated with Liver Injury
2.6. The Recruitment of Ly6Chigh MoMφs Was Sex-Dependent and Correlated with Liver Damage
2.7. The Expression of the Genes of Pro-Inflammatory Cytokines Was Differently Regulated in Males and Females during Chronic Damage
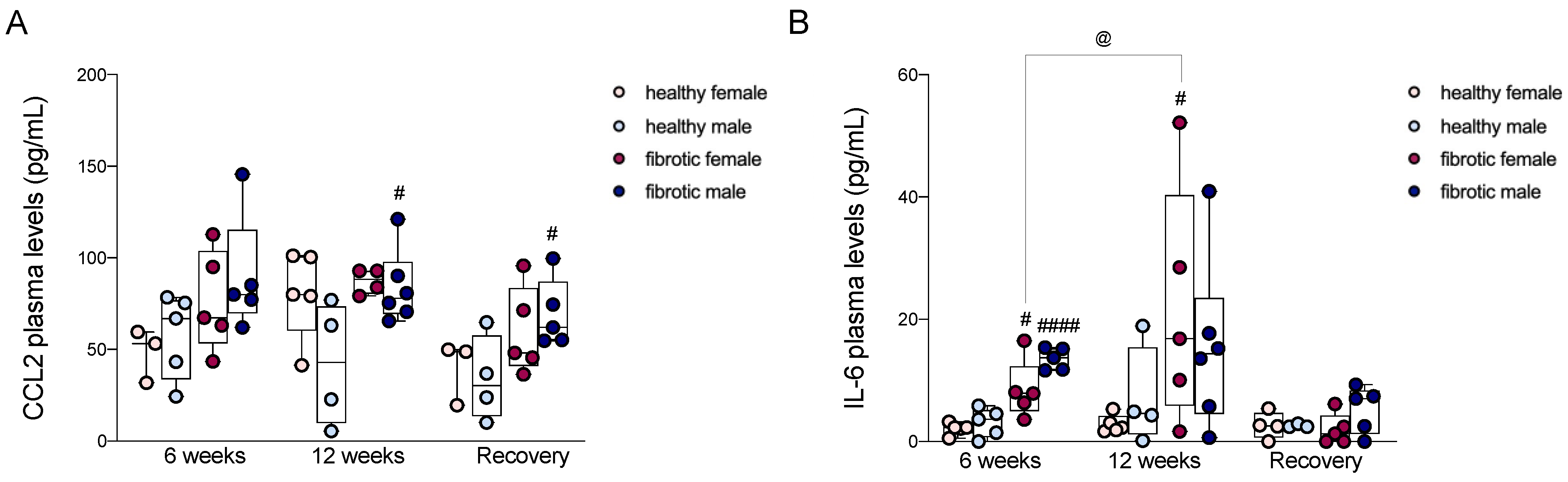
2.8. Circulating Levels of CCL2 and IL-6 Were Strongly Associated with Liver Fibrosis
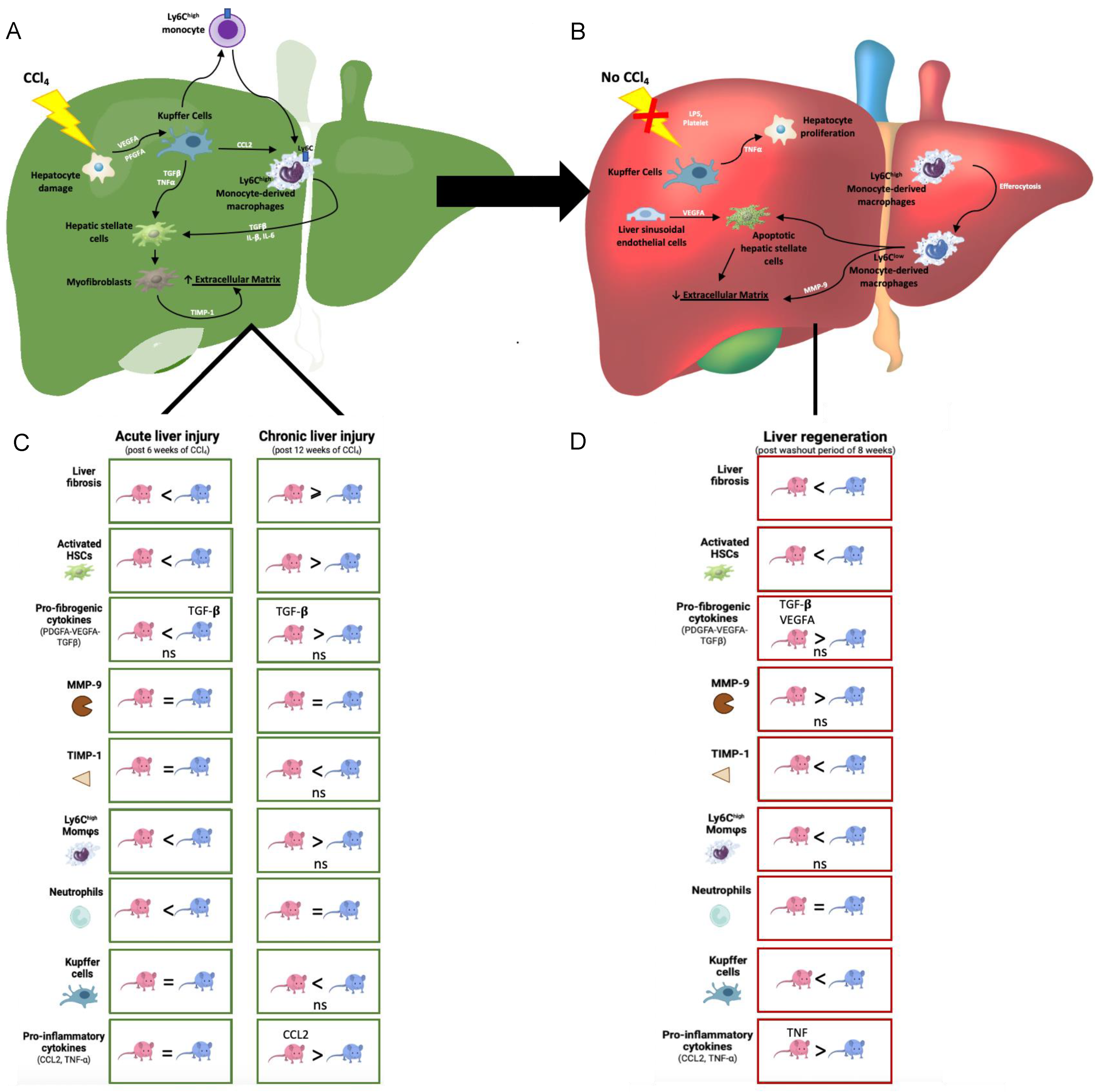
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Model of Liver Injury
4.3. Flow Cytometry (FCM)
4.4. Hydroxyproline Assay
4.5. Histological Evaluation of Liver Fibrosis
4.6. Immunohistochemical Analysis
4.7. Quantification of the Expression of mRNA via qRT-PCR
4.8. Quantification of CCL2 and IL-6 Plasma Levels
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef]
- Parola, M.; Pinzani, M. Liver fibrosis: Pathophysiology, pathogenetic targets and clinical issues. Mol. Asp. Med. 2019, 65, 37–55. [Google Scholar] [CrossRef]
- Russo, F.P.; Parola, M. Stem cells in liver failure. Best Pract. Res. Clin. Gastroenterol. 2012, 26, 35–45. [Google Scholar] [CrossRef]
- Kisseleva, T.; Brenner, D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 151–166. [Google Scholar] [CrossRef]
- Bert, F. vcLiver Fibrosis: Difficulties in Diagnostic and Treatment: A Review. Gastroenterol. Med. Res. 2017, 1, GMR.000502. [Google Scholar] [CrossRef]
- Aydın, M.M.; Akçalı, K.C. Liver fibrosis. Turk. J. Gastroenterol. 2018, 29, 14–21. [Google Scholar] [CrossRef]
- Dhar, D.; Baglieri, J.; Kisseleva, T.; Brenner, D.A. Mechanisms of liver fibrosis and its role in liver cancer. Exp. Biol. Med. 2020, 245, 96–108. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Liu, R.-X.; Hou, F.; Cui, L.-J.; Li, C.-Y.; Chi, C.; Yi, E.; Wen, Y.; Yin, C.-H. Fibronectin expression is critical for liver fibrogenesis in vivo and in vitro. Mol. Med. Rep. 2016, 14, 3669–3675. [Google Scholar] [CrossRef]
- Campana, L.; Esser, H.; Huch, M.; Forbes, S. Liver regeneration and inflammation: From fundamental science to clinical applications. Nat. Rev. Mol. Cell Biol. 2021, 22, 608–624. [Google Scholar] [CrossRef]
- Kratofil, R.M.; Kubes, P.; Deniset, J.F. Monocyte Conversion During Inflammation and Injury. Arter. Thromb. Vasc. Biol. 2017, 37, 35–42. [Google Scholar] [CrossRef]
- Sun, M.; Kisseleva, T. Reversibility of Liver Fibrosis. Clin. Res. Hepatol. Gastroenterol. 2015, 39, S60–S63. [Google Scholar] [CrossRef]
- Guy, J.; Peters, M.G. Liver disease in women: The influence of gender on epidemiology, natural history, and patient outcomes. Gastroenterol. Hepatol. 2013, 9, 633–639. [Google Scholar]
- Sayaf, K.; Gabbia, D.; Russo, F.P.; De Martin, S. The Role of Sex in Acute and Chronic Liver Damage. Int. J. Mol. Sci. 2022, 23, 10654. [Google Scholar] [CrossRef]
- Marcos, R.; Lopes, C.; Malhão, F.; Correia-Gomes, C.; Fonseca, S.; Lima, M.; Gebhardt, R.; Rocha, E. Stereological assessment of sexual dimorphism in the rat liver reveals differences in hepatocytes and Kupffer cells but not hepatic stellate cells. J. Anat. 2016, 228, 996–1005. [Google Scholar] [CrossRef]
- Bizzaro, D.; Crescenzi, M.; Di Liddo, R.; Arcidiacono, D.; Cappon, A.; Bertalot, T.; Amodio, V.; Tasso, A.; Stefani, A.; Bertazzo, V.; et al. Sex-dependent differences in inflammatory responses during liver regeneration in a murine model of acute liver injury. Clin. Sci. 2018, 132, 255–272. [Google Scholar] [CrossRef]
- Gabbia, D.; Carpi, S.; Sarcognato, S.; Zanotto, I.; Sayaf, K.; Colognesi, M.; Polini, B.; Digiacomo, M.; Macchia, M.; Nieri, P.; et al. The phenolic compounds tyrosol and hydroxytyrosol counteract liver fibrogenesis via the transcriptional modulation of NADPH oxidases and oxidative stress-related miRNAs. Biomed. Pharmacother. 2023, 157, 114014. [Google Scholar] [CrossRef]
- Zhang, W.; Conway, S.J.; Liu, Y.; Snider, P.; Chen, H.; Gao, H.; Liu, Y.; Isidan, K.; Lopez, K.J.; Campana, G.; et al. Heterogeneity of Hepatic Stellate Cells in Fibrogenesis of the Liver: Insights from Single-Cell Transcriptomic Analysis in Liver Injury. Cells 2021, 10, 2129. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. Transforming growth factor–β in tissue fibrosis. J. Exp. Med. 2020, 217, e20190103. [Google Scholar] [CrossRef]
- Yang, L.; Kwon, J.; Popov, Y.; Gajdos, G.B.; Ordog, T.; Brekken, R.A.; Mukhopadhyay, D.; Schuppan, D.; Bi, Y.; Simonetto, D.; et al. Vascular Endothelial Growth Factor Promotes Fibrosis Resolution and Repair in Mice. Gastroenterology 2014, 146, 1339–1350.e1. [Google Scholar] [CrossRef]
- Ying, H.-Z.; Chen, Q.; Zhang, W.-Y.; Zhang, H.-H.; Ma, Y.; Zhang, S.-Z.; Fang, J.; Yu, C.-H. PDGF signaling pathway in hepatic fibrosis pathogenesis and therapeutics (Review). Mol. Med. Rep. 2017, 16, 7879–7889. [Google Scholar] [CrossRef]
- Kolios, G.; Valatas, V.; Kouroumalis, E. Role of Kupffer cells in the pathogenesis of liver disease. World J. Gastroenterol. 2006, 12, 7413–7420. [Google Scholar] [CrossRef] [PubMed]
- Geervliet, E.; Bansal, R. Matrix Metalloproteinases as Potential Biomarkers and Therapeutic Targets in Liver Diseases. Cells 2020, 9, 1212. [Google Scholar] [CrossRef] [PubMed]
- Roderfeld, M.; Weiskirchen, R.; Wagner, S.; Berres, M.-L.; Henkel, C.; Grötzinger, J.; Gressner, A.M.; Matern, S.; Roeb, E. Inhibition of hepatic fibrogenesis by matrix metalloproteinase-9 mutants in mice. FASEB J. 2006, 20, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Wang, H.; Jiang, Y.; Zhang, Y.; Bao, Y.; Zhang, X.; Zhang, J.; Guo, H.; Yang, F.; Luan, Y.; et al. Ameliorative effects of tannic acid on carbon tetrachloride-induced liver fibrosis in vivo and in vitro. J. Pharmacol. Sci. 2016, 130, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Wang, F.-S.; Xu, R. Neutrophils in liver diseases: Pathogenesis and therapeutic targets. Cell Mol. Immunol. 2021, 18, 38–44. [Google Scholar] [CrossRef]
- Tang, J.; Yan, Z.; Feng, Q.; Yu, L.; Wang, H. The Roles of Neutrophils in the Pathogenesis of Liver Diseases. Front. Immunol. 2021, 12, 625472. [Google Scholar] [CrossRef]
- Alazawi, W.; Knolle, P.A. Interfering with Kupffer cell replenishment: New insights into liver injury. J. Hepatol. 2018, 68, 635–637. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Pan, G.; Xiang, L.; Luo, D.; Shao, J. Occurrences and Functions of Ly6Chi and Ly6Clo Macrophages in Health and Disease. Front. Immunol. 2022, 13, 901672. [Google Scholar] [CrossRef]
- Xi, S.; Zheng, X.; Li, X.; Jiang, Y.; Wu, Y.; Gong, J.; Jie, Y.; Li, Z.; Cao, J.; Sha, L.; et al. Activated Hepatic Stellate Cells Induce Infiltration and Formation of CD163+ Macrophages via CCL2/CCR2 Pathway. Front. Med. 2021, 8, 627927. [Google Scholar] [CrossRef]
- Tiegs, G.; Horst, A.K. TNF in the liver: Targeting a central player in inflammation. Semin. Immunopathol. 2022, 44, 445–459. [Google Scholar] [CrossRef]
- Yang, Y.M.; Seki, E. TNFα in Liver Fibrosis. Curr. Pathobiol. Rep. 2015, 3, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Schwabe, R.F.; Brenner, D.A. Mechanisms of Liver Injury. I. TNF-α-induced liver injury: Role of IKK, JNK, and ROS pathways. Am. J. Physiol.-Gastrointest. Liver Physiol. 2006, 290, G583–G589. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.Y.; Lu, R.H.; Tsai, Y.T.; Lin, H.C.; Hou, M.C.; Li, C.P.; Liao, T.M.; Lin, L.F.; Wang, S.S.; Lee, S.D. Plasma Interleukin-6 Levels in Patients with Cirrhosis Relationship to Endotoxemia, Tumor Necrosis Factor-α, and Hyperdynamic Circulation. Scand. J. Gastroenterol. 1996, 31, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Queck, A.; Bode, H.; Uschner, F.E.; Brol, M.J.; Graf, C.; Schulz, M.; Jansen, C.; Praktiknjo, M.; Schierwagen, R.; Klein, S.; et al. Systemic MCP-1 Levels Derive Mainly From Injured Liver and Are Associated With Complications in Cirrhosis. Front. Immunol. 2020, 11, 354. [Google Scholar] [CrossRef]
- Wu, J.; Zern, M.A. Hepatic stellate cells: A target for the treatment of liver fibrosis. J. Gastroenterol. 2000, 35, 665–672. [Google Scholar] [CrossRef]
- Borst, K.; Graalmann, T.; Kalinke, U. Reply to: “Lack of Kupffer cell depletion in diethylnitrosamine-induced hepatic inflammation”. J. Hepatol. 2019, 70, 815–816. [Google Scholar] [CrossRef]
- Ramirez-Pedraza, M.; Fernández, M. Interplay Between Macrophages and Angiogenesis: A Double-Edged Sword in Liver Disease. Front. Immunol. 2019, 10, 2882. [Google Scholar] [CrossRef]
- Secchi, M.F.; Crescenzi, M.; Masola, V.; Russo, F.P.; Floreani, A.; Onisto, M. Heparanase and macrophage interplay in the onset of liver fibrosis. Sci. Rep. 2017, 7, 14956. [Google Scholar] [CrossRef]
- Wen, Y.; Lambrecht, J.; Ju, C.; Tacke, F. Hepatic macrophages in liver homeostasis and diseases-diversity, plasticity and therapeutic opportunities. Cell Mol. Immunol. 2021, 18, 45–56. [Google Scholar] [CrossRef]
- Cheng, D.; Chai, J.; Wang, H.; Fu, L.; Peng, S.; Ni, X. Hepatic macrophages: Key players in the development and progression of liver fibrosis. Liver Int. 2021, 41, 2279–2294. [Google Scholar] [CrossRef]
- Cordero-Espinoza, L.; Huch, M. The balancing act of the liver: Tissue regeneration versus fibrosis. J. Clin. Investig. 2018, 128, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Cervera, A.; Soehnlein, O.; Kenne, E. Neutrophils in chronic inflammatory diseases. Cell Mol. Immunol. 2022, 19, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Kessler, S.M.; Hoppstädter, J.; Hosseini, K.; Laggai, S.; Haybaeck, J.; Kiemer, A.K. Lack of Kupffer cell depletion in diethylnitrosamine-induced hepatic inflammation. J. Hepatol. 2019, 70, 813–815. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, P.; Pellicoro, A.; Vernon, M.A.; Boulter, L.; Aucott, R.L.; Ali, A.; Hartland, S.N.; Snowdon, V.K.; Cappon, A.; Gordon-Walker, T.T.; et al. Differential Ly-6C expression identifies the recruited macrophage phenotype, which orchestrates the regression of murine liver fibrosis. Proc. Natl. Acad. Sci. USA 2012, 109, E3186–E3195. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (3′-5′) |
|---|---|---|
| TNFα | CCCACGTCGTAGCAAACCA | TGTCTTTGAGATCCATGCCGT |
| TGFβ | GTGGAAATCAACGGGATCAGC | GTTGGTATCCAGGGCTCTCC |
| VEGFA | ACTGGACCCTGGCTTTACTG | CTCTCCTTCTGTCGTGGGTG |
| CCL2 | CCACAACCACCTCAAGCACT | AGGCATCACAGTCCGAGTCA |
| PDGFA | CTGTGTTCCTCTGCCCCTTT | TGTCATGTCTCCATGCTGCC |
| β-ACTIN | AGCAAGCAGGAGGATGAG | AAAACGCAGCTCAGTAACAGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayaf, K.; Zanotto, I.; Gabbia, D.; Alberti, D.; Pasqual, G.; Zaramella, A.; Fantin, A.; De Martin, S.; Russo, F.P. Sex Drives Functional Changes in the Progression and Regression of Liver Fibrosis. Int. J. Mol. Sci. 2023, 24, 16452. https://doi.org/10.3390/ijms242216452
Sayaf K, Zanotto I, Gabbia D, Alberti D, Pasqual G, Zaramella A, Fantin A, De Martin S, Russo FP. Sex Drives Functional Changes in the Progression and Regression of Liver Fibrosis. International Journal of Molecular Sciences. 2023; 24(22):16452. https://doi.org/10.3390/ijms242216452
Chicago/Turabian StyleSayaf, Katia, Ilaria Zanotto, Daniela Gabbia, Dafne Alberti, Giulia Pasqual, Alice Zaramella, Alberto Fantin, Sara De Martin, and Francesco Paolo Russo. 2023. "Sex Drives Functional Changes in the Progression and Regression of Liver Fibrosis" International Journal of Molecular Sciences 24, no. 22: 16452. https://doi.org/10.3390/ijms242216452