

The Peptide AWRK6 Alleviates Lipid Accumulation in Hepatocytes by Inhibiting miR-5100 Targeting G6PC

Abstract
:1. Introduction
2. Results
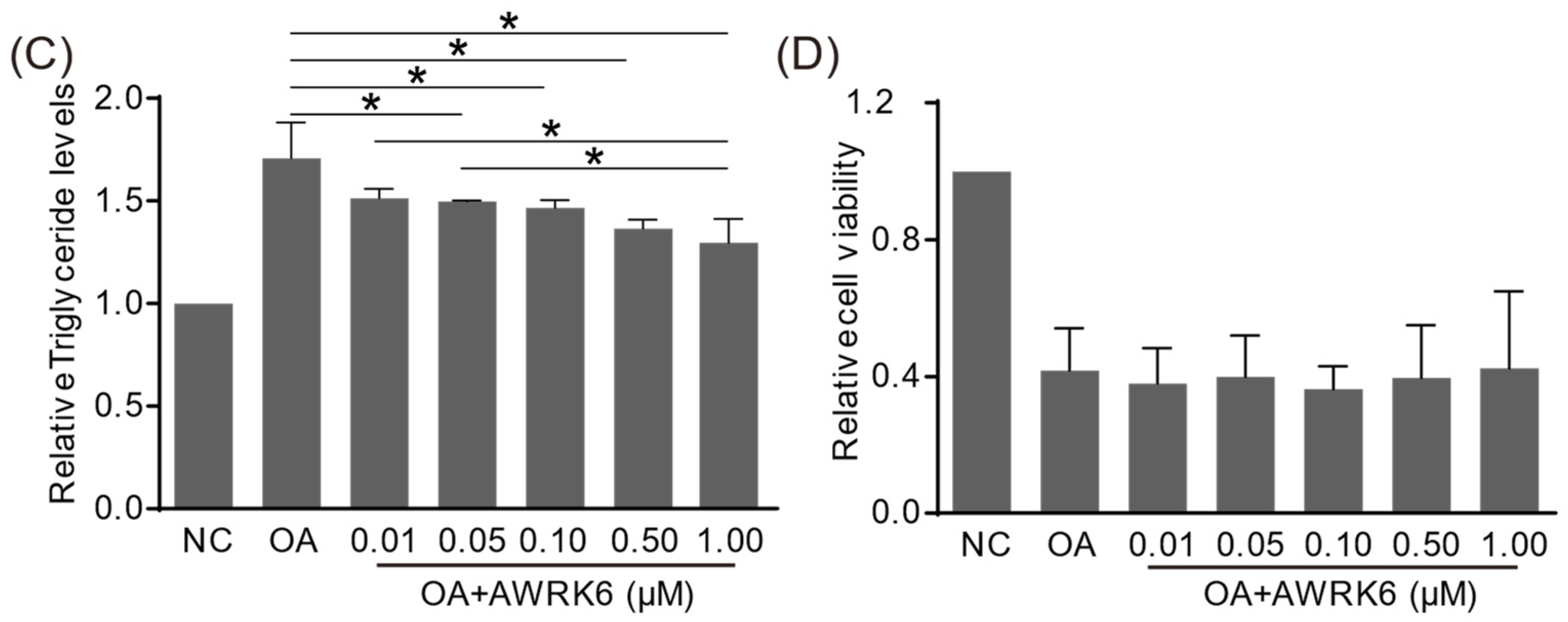
2.1. AWRK6 Alleviated Lipid Accumulation Induced by Oleic Acid
2.2. AWRK6 Reduced Oleic Acid-Elevated miR-5100 and miR-505
2.3. miR-5100 Mediated Alleviation of Lipid Accumulation by AWRK6
2.4. G6PC Was a Direct Target of miR-5100
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Viability Assay
4.3. Triglyceride Determination
4.4. Real-Time PCR
4.5. Western Blotting
4.6. Dual-Luciferase Reporter Assay
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Riazi, K.; Azhari, H.; Charette, J.H.; Underwood, F.E.; King, J.A.; Afshar, E.E.; Swain, M.G.; Congly, S.E.; Kaplan, G.G.; Shaheen, A.A. The prevalence and incidence of NAFLD worldwide: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2022, 7, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, J.V.; Mark, H.E.; Anstee, Q.M.; Arab, J.P.; Batterham, R.L.; Castera, L.; Cortez-Pinto, H.; Crespo, J.; Cusi, K.; Dirac, M.A.; et al. Advancing the global public health agenda for NAFLD: A consensus statement. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 60–78. [Google Scholar] [CrossRef] [PubMed]
- Neuschwander-Tetri, B.A. Therapeutic Landscape for NAFLD in 2020. Gastroenterology 2020, 158, 1984–1998.e1983. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.; Finck, B.N. Emerging therapeutic approaches for the treatment of NAFLD and type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2021, 17, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Gjorgjieva, M.; Sobolewski, C.; Dolicka, D.; Correia de Sousa, M.; Foti, M. miRNAs and NAFLD: From pathophysiology to therapy. Gut 2019, 68, 2065–2079. [Google Scholar] [CrossRef] [PubMed]
- Hochreuter, M.Y.; Dall, M.; Treebak, J.T.; Barres, R. MicroRNAs in non-alcoholic fatty liver disease: Progress and perspectives. Mol. Metab. 2022, 65, 101581. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Sun, Y.; Li, Y.; Zhang, H.; Yu, W.; Li, Y.; Xin, Y.; Alsareii, S.A.; Wang, Q.; Zhang, D. A synthetic peptide AWRK6 ameliorates metabolic associated fatty liver disease: Involvement of lipid and glucose homeostasis. Peptides 2021, 143, 170597. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. MiRNA Regulation in the Improvement of Lipid Accumulation in Metabolic Associated Fatty Liver Disease by AWRK6; Liaoning University: Shenyang, China, 2022. [Google Scholar] [CrossRef]
- Loomba, R.; Friedman, S.L.; Shulman, G.I. Mechanisms and disease consequences of nonalcoholic fatty liver disease. Cell 2021, 184, 2537–2564. [Google Scholar] [CrossRef] [PubMed]
- Monteillet, L.; Gjorgjieva, M.; Silva, M.; Verzieux, V.; Imikirene, L.; Duchampt, A.; Guillou, H.; Mithieux, G.; Rajas, F. Intracellular lipids are an independent cause of liver injury and chronic kidney disease in non alcoholic fatty liver disease-like context. Mol. Metab. 2018, 16, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Latorre, J.; Moreno-Navarrete, J.M.; Mercader, J.M.; Sabater, M.; Rovira, O.; Girones, J.; Ricart, W.; Fernandez-Real, J.M.; Ortega, F.J. Decreased lipid metabolism but increased FA biosynthesis are coupled with changes in liver microRNAs in obese subjects with NAFLD. Int. J. Obes. 2017, 41, 620–630. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; You, H.J.; Lee, G.; Lee, S.H.; Yoo, T.; Choi, M.; Joo, S.K.; Park, J.H.; Chang, M.S.; Lee, D.H.; et al. Interaction effect between NAFLD severity and high carbohydrate diet on gut microbiome alteration and hepatic de novo lipogenesis. Gut Microbes 2022, 14, 2078612. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Yi, H.; Feng, J.; Zhang, Z.; Zhou, J.; Zhou, L.; Zhou, J.; Li, M.; Li, Q. Intermittent hypoxia disrupts glucose homeostasis in liver cells in an insulin-dependent and independent manner. Cell. Physiol. Biochem 2018, 47, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- Kobori, T.; Tanaka, C.; Tameishi, M.; Urashima, Y.; Ito, T.; Obata, T. Role of Ezrin/Radixin/Moesin in the surface localization of programmed cell death ligand-1 in human colon adenocarcinoma LS180 cells. Pharmaceuticals 2021, 14, 864. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) |
|---|---|
| miR-5100 RT | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACGGTACA |
| miR-5100 forward | GCTTCAGATCCCAGCGGT |
| miR-505 RT | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGGAAA |
| miR-505 forward | GCCGTCAACACTTGCTGG |
| miRNA reverse | AGTGCAGGGTCCGAGGTATT |
| U6 RT and reverse | CGAATTTGCGTGTCATCCT |
| U6 forward | CTCGCTTCGGCAGCACATA |
| G6PC forward [13] | CTACTACAGCAACACTTCCGTG |
| G6PC reverse [13] | GGTCGGCTTTATCTTTCCCTGA |
| β-actin forward [14] | TGGCACCCAGCACAATGAA |
| β-actin reverse [14] | CTAAGTCATAGTCCGCCTAGAAGCA |
| Name | Sequence (5′-3′) |
|---|---|
| site 1 wildtype forward | CAATTACTATATTTTAGATGCAATGTGATCTGAAGTTTCTAATTCTGGCCC |
| site 1 wildtype reverse | TCGAGGGCCAGAATTAGAAACTTCAGATCACATTGCATCTAAAATATAGTAATTGAGCT |
| site 1 mutant forward | CAATTACTATATTTTAGATGCAATGTGAGAGTAAGTTTCTAATTCTGGCCC |
| site 1 mutant reverse | TCGAGGGCCAGAATTAGAAACTTACTCTCACATTGCATCTAAAATATAGTAATTGAGCT |
| site 2 wildtype forward | CGTTACATTTGAAGACATGTCTGCATATCTGAAATTCCAGCCCTAATTAAC |
| site 2 wildtype reverse | TCGAGTTAATTAGGGCTGGAATTTCAGATATGCAGACATGTCTTCAAATGTAACGAGCT |
| site 2 mutant forward | CGTTACATTTGAAGACATGTCTGCATAGAGTAAATTCCAGCCCTAATTAAC |
| site 2 mutant reverse | TCGAGTTAATTAGGGCTGGAATTTACTCTATGCAGACATGTCTTCAAATGTAACGAGCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Liu, Y.; Wang, Q.; Jin, L.; Zhang, D. The Peptide AWRK6 Alleviates Lipid Accumulation in Hepatocytes by Inhibiting miR-5100 Targeting G6PC. Int. J. Mol. Sci. 2023, 24, 16141. https://doi.org/10.3390/ijms242216141
Liu J, Liu Y, Wang Q, Jin L, Zhang D. The Peptide AWRK6 Alleviates Lipid Accumulation in Hepatocytes by Inhibiting miR-5100 Targeting G6PC. International Journal of Molecular Sciences. 2023; 24(22):16141. https://doi.org/10.3390/ijms242216141
Chicago/Turabian StyleLiu, Jiaxin, Ying Liu, Qiuyu Wang, Lili Jin, and Dianbao Zhang. 2023. "The Peptide AWRK6 Alleviates Lipid Accumulation in Hepatocytes by Inhibiting miR-5100 Targeting G6PC" International Journal of Molecular Sciences 24, no. 22: 16141. https://doi.org/10.3390/ijms242216141