
Osa-miR11117 Targets OsPAO4 to Regulate Rice Immunity against the Blast Fungus Magnaporthe oryzae
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
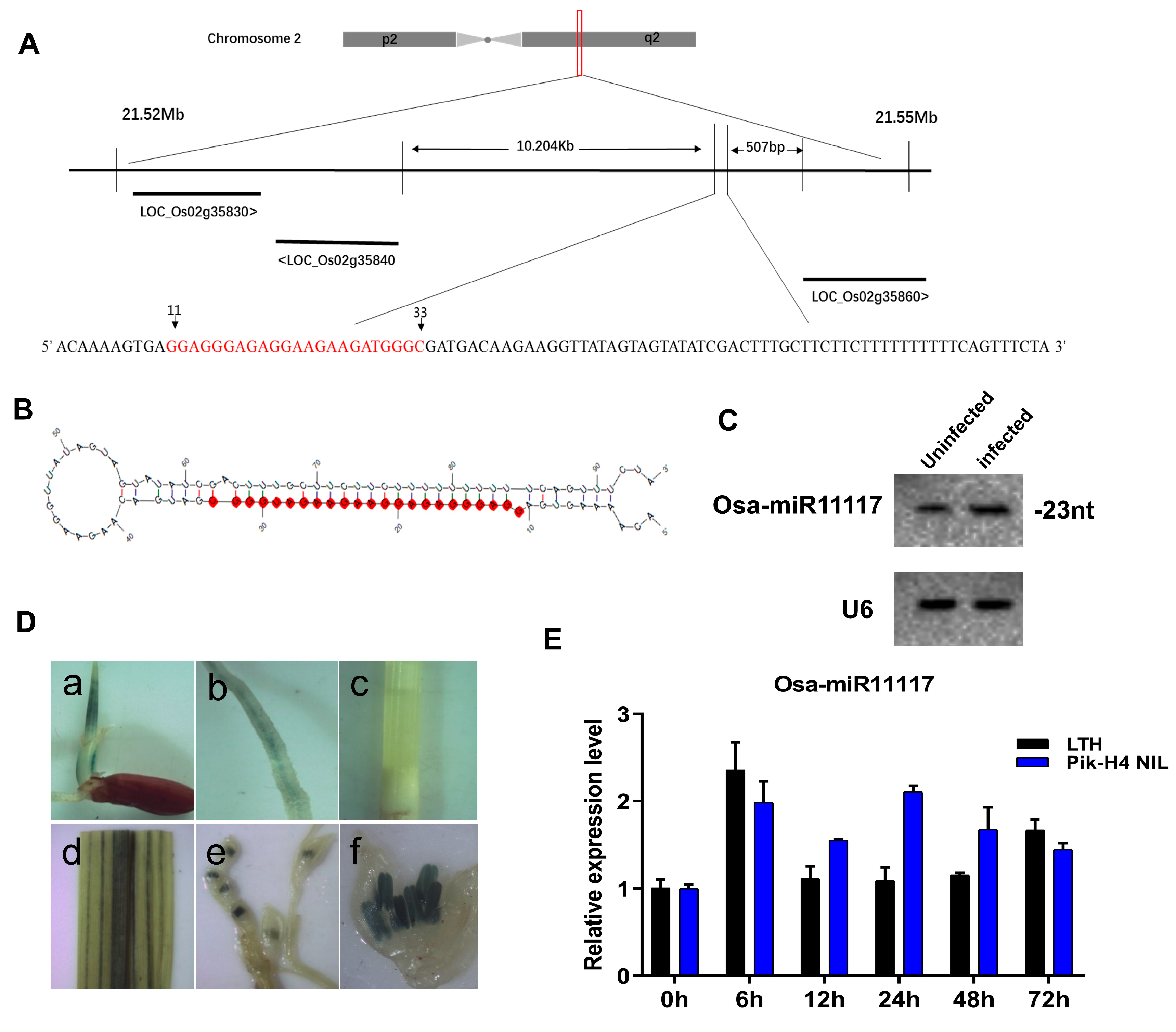
2.1. Osa-miR11117 Is Responsive to Blast Fungus
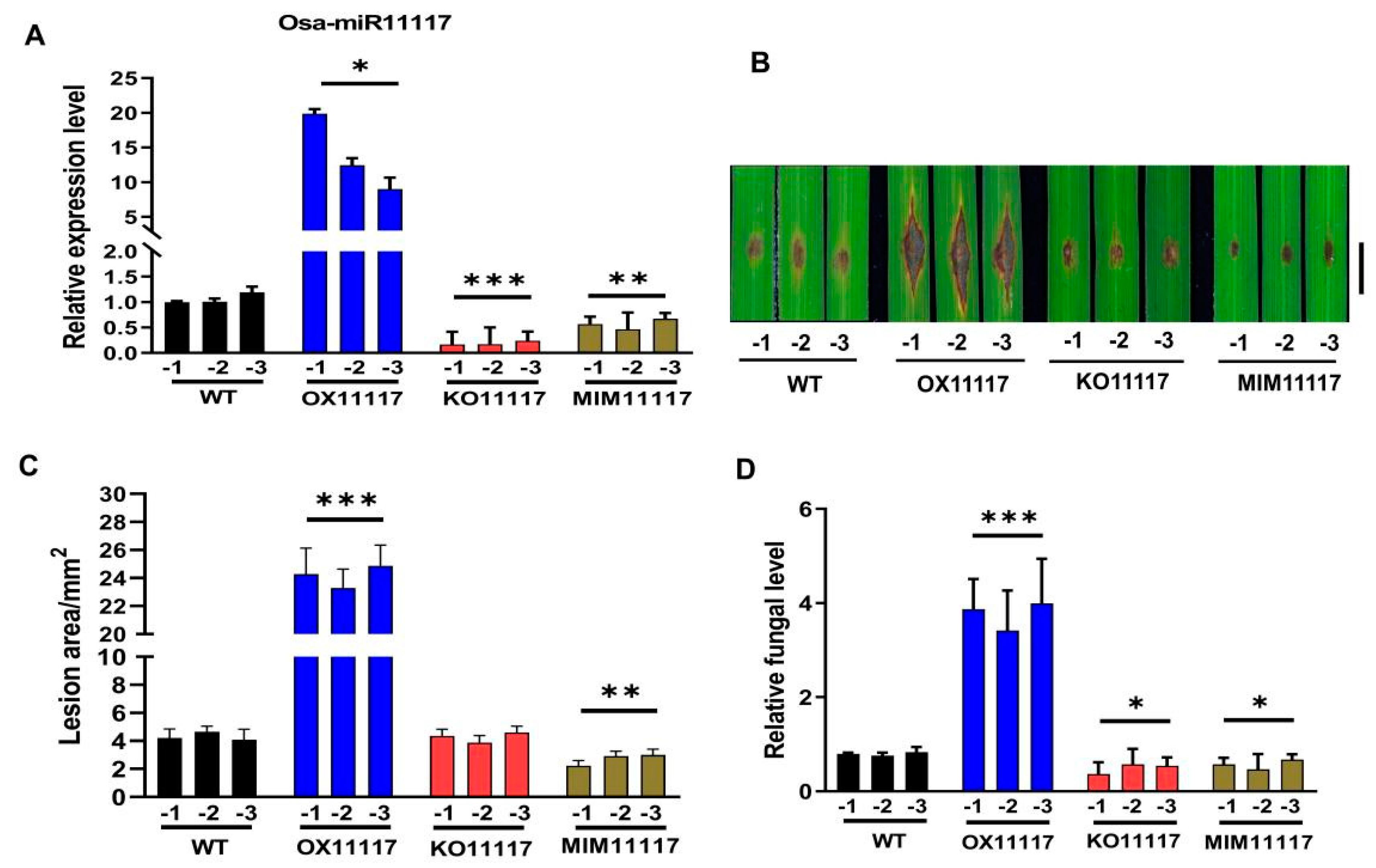
2.2. Overexpression of Osa-miR11117 Enhances Susceptibility to Infection by M. oryzae
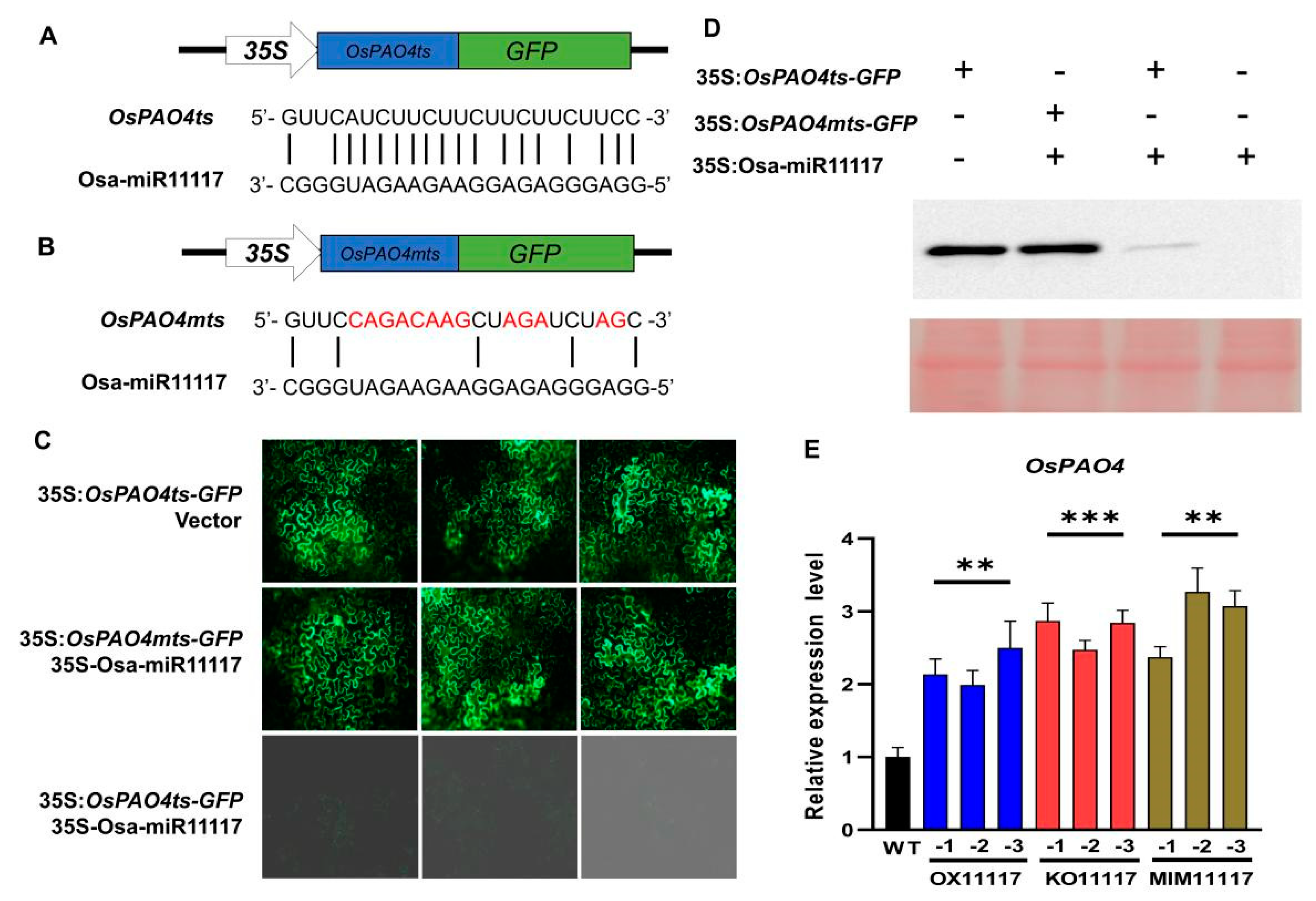
2.3. OsPAO4 Is One of the Target Genes of Osa-miR11117
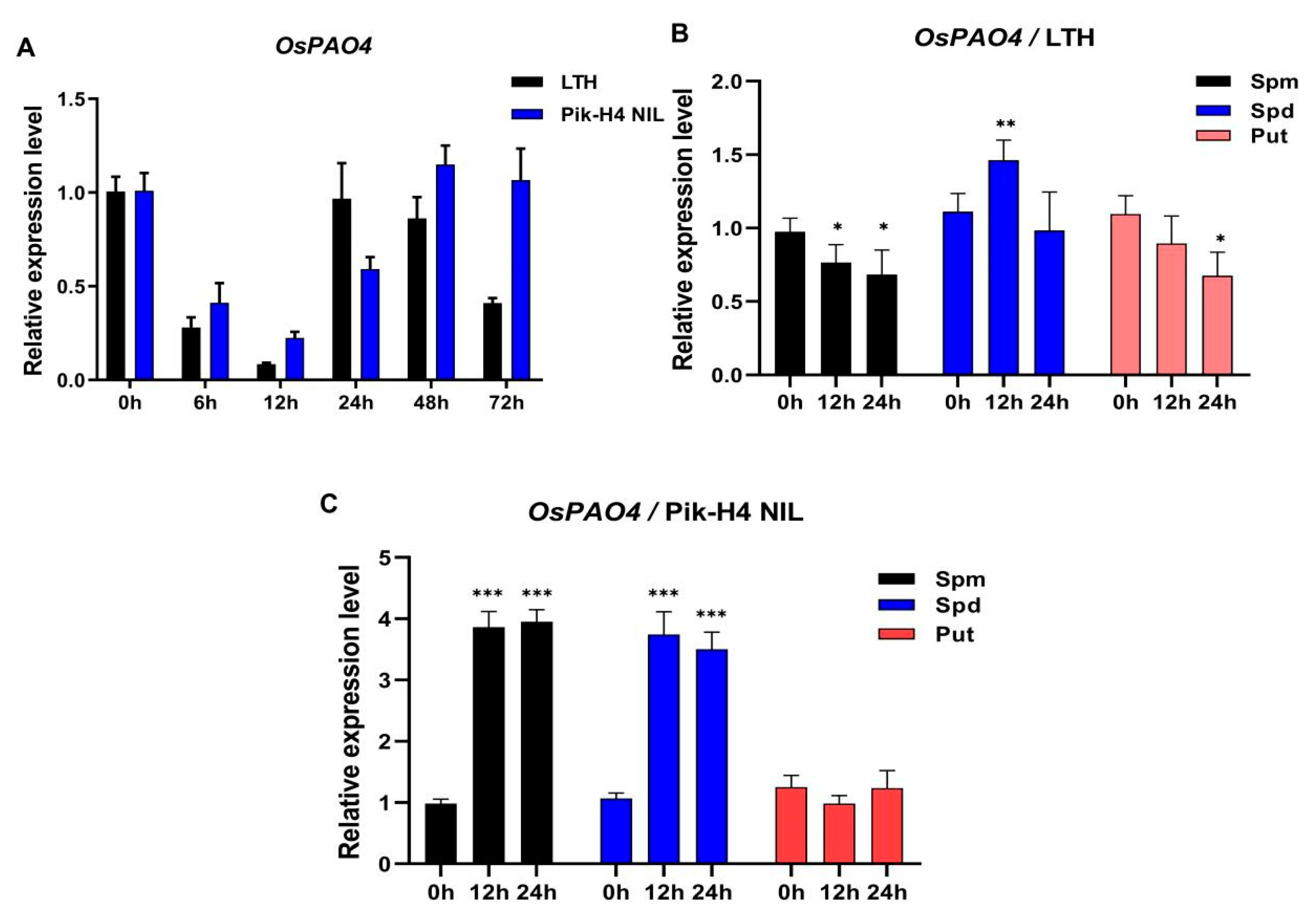
2.4. OsPAO4 Is One M. oryzae and PA Responsive PAO Gene
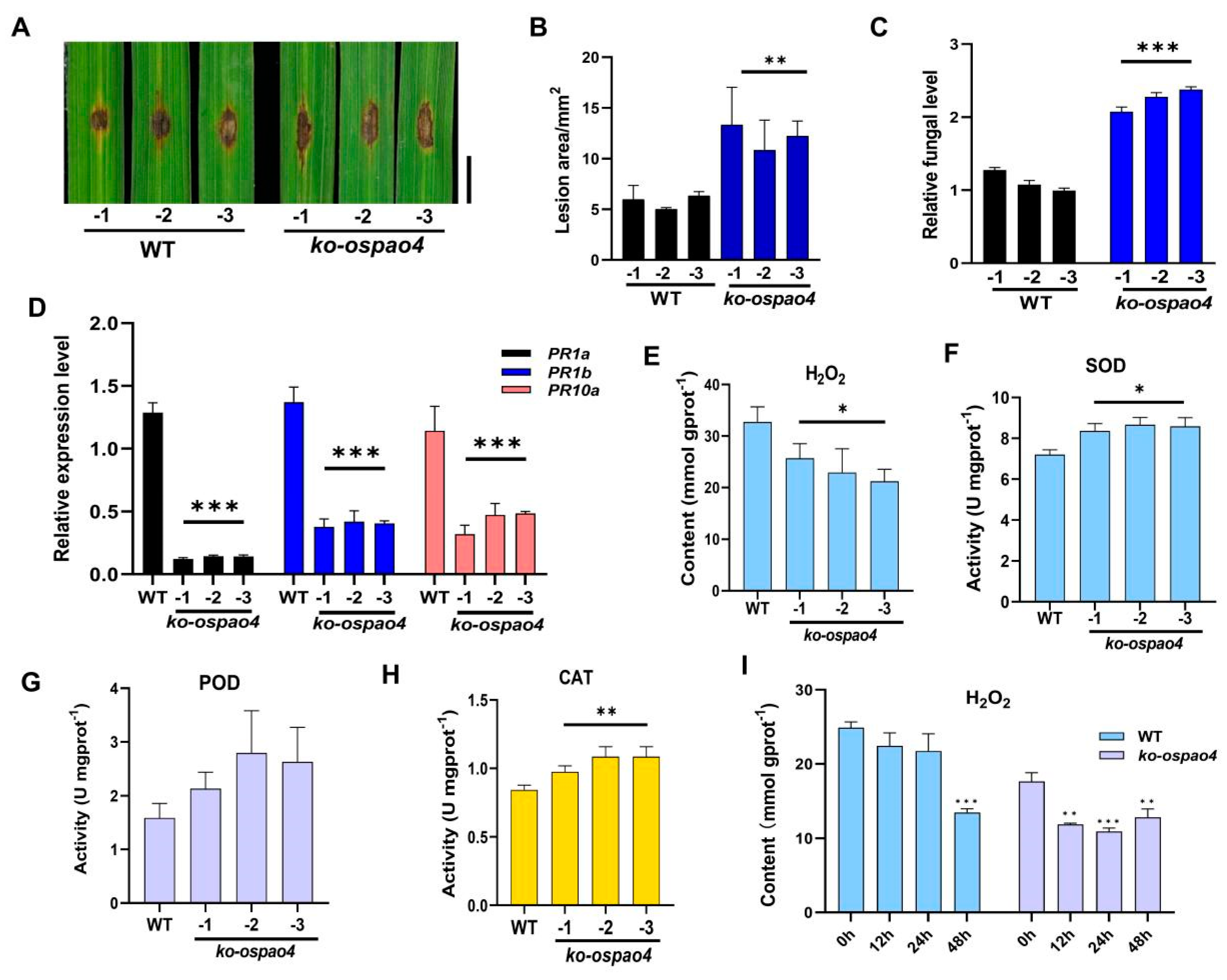
2.5. Knockout of OsPAO4 Enhances the Susceptibility to Infection by M. oryzae
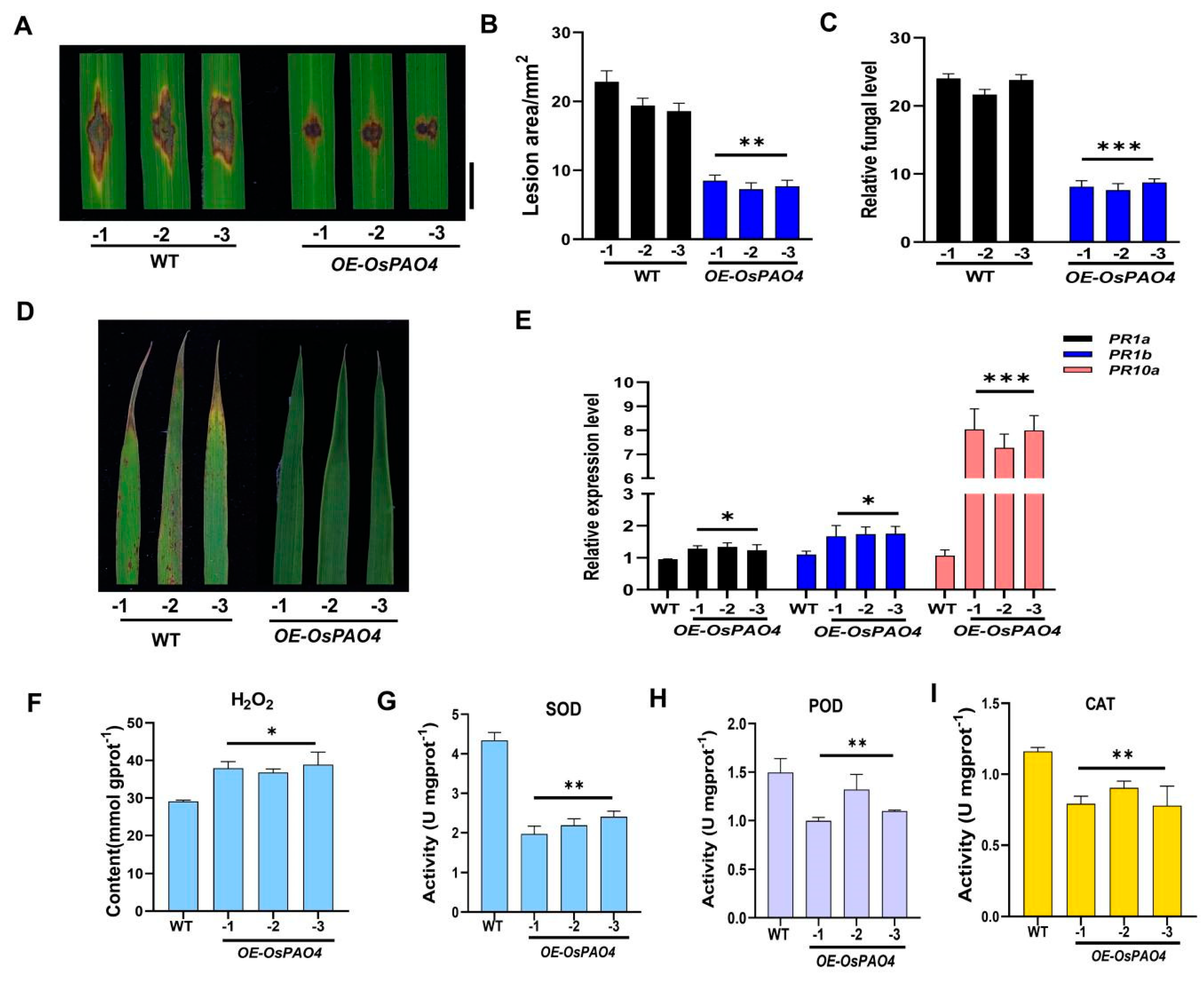
2.6. Overexpression of OsPAO4 Enhances the Resistance against M. oryzae
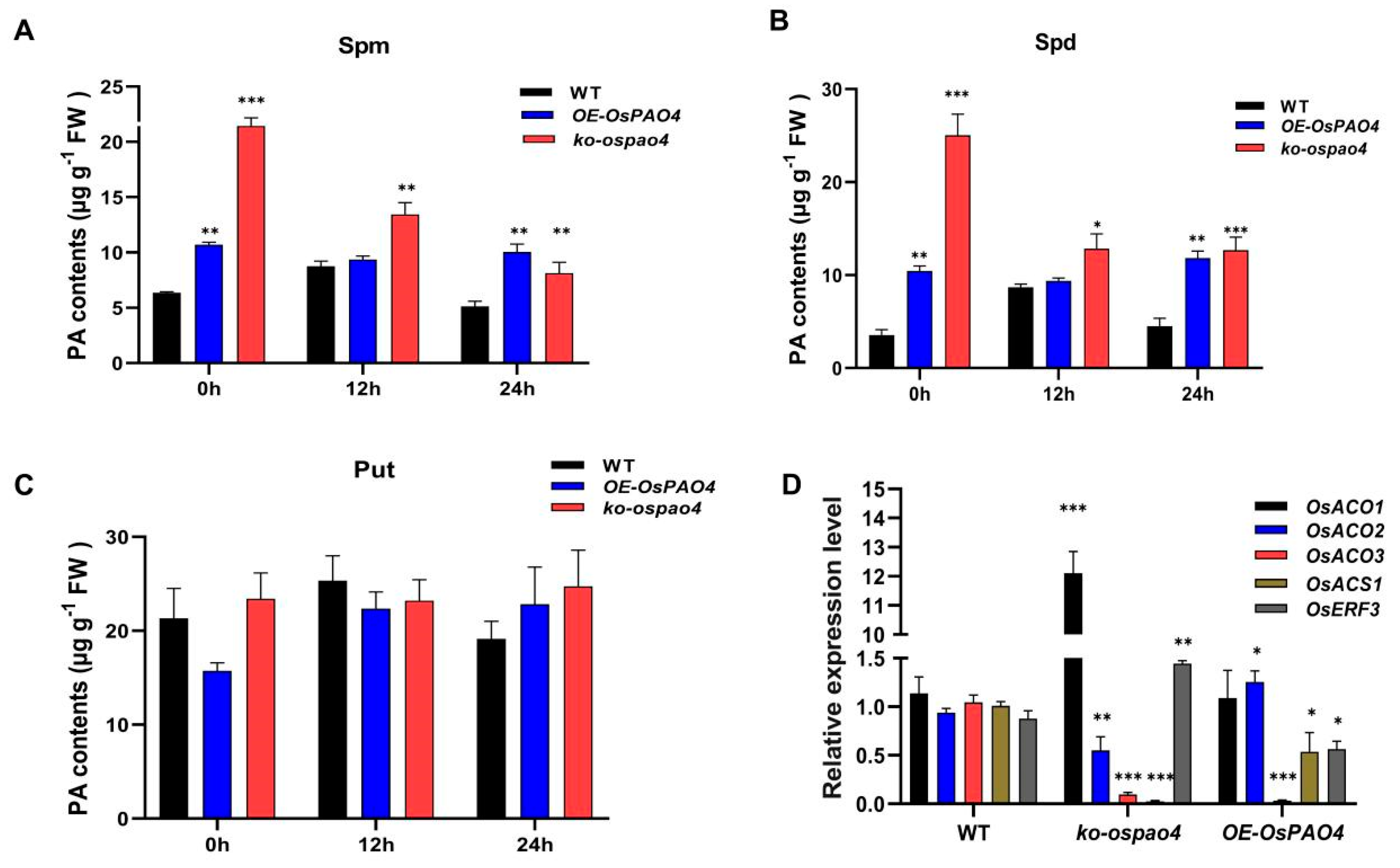
2.7. OsPAO4 Regulates Rice Blast Resistance by Controlling Both the Accumulation of PA and the Ethylene Signaling Pathway
3. Discussion
3.1. Impact of Osa-miR11117 on Rice Blast Resistance
3.2. OsPAO4 Is a Direct Target of Osa-miR11117
3.3. OsPAO4 Positively Regulates Rice Blast Resistance
3.4. Association of OsPAO4 with Polyamine Metabolism and Ethylene Pathway
4. Methods
4.1. Plant Materials and Growth Conditions
4.2. Plasmid Construction and Genetic Transformation
4.3. Pathogen Growth, Infection, and PAs Treatment
4.4. GUS Staining Assay
4.5. RNA Isolation, Quantitative RT-PCR, and Northern Blot
4.6. Agrobacterium-Mediated Transient Expression Analysis in N. benthamiana
4.7. H2O2 Measurement and Enzymatic Activity Assay
4.8. Extraction and Analysis of PAs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ramirez-Prado, J.S.; Abulfaraj, A.A.; Rayapuram, N.; Benhamed, M.; Hirt, H. Plant Immunity: From Signaling to Epigenetic Control of Defense. Trends Plant Sci. 2018, 23, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.K.; Wu, J.G.; Lii, Y.F.; Li, Y.; Jin, H.L. Contribution of Small RNA Pathway Components in Plant Immunity. Mol. Plant Microbe 2013, 26, 617–625. [Google Scholar] [CrossRef]
- Deng, Y.T.; Liu, M.L.; Li, X.F.; Li, F. microRNA-mediated R gene regulation: Molecular scabbards for double-edged swords. Sci. China Life Sci. 2018, 61, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Broughton, J.P.; Lovci, M.T.; Huang, J.L.; Yeo, G.W.; Pasquinelli, A.E. Pairing beyond the Seed Supports MicroRNA Targeting Specificity. Mol. Cell 2016, 64, 320–333. [Google Scholar] [CrossRef]
- Katiyar-Agarwal, S.; Jin, H.L. Role of Small RNAs in Host-Microbe Interactions. Annu. Rev. Phytopathol. 2010, 48, 225–246. [Google Scholar] [CrossRef]
- Zhang, F.Y.; Yang, J.W.; Zhang, N.; Wu, J.H.; Si, H.J. Roles of microRNAs in abiotic stress response and characteristics regulation of plant. Front. Plant Sci. 2022, 13, 919243. [Google Scholar] [CrossRef]
- Gupta, O.P.; Sharma, P.; Gupta, R.K.; Sharma, I. Current status on role of miRNAs during plant–fungus interaction. Physiol. Mol. Plant Pathol. 2014, 85, 1–7. [Google Scholar] [CrossRef]
- Li, Y.; Lu, Y.G.; Shi, Y.; Wu, L.; Xu, Y.J.; Huang, F.; Guo, X.Y.; Zhang, Y.; Fan, J.; Zhao, J.Q.; et al. Multiple Rice MicroRNAs Are Involved in Immunity against the Blast Fungus Magnaporthe oryzae. Plant Physiol. 2014, 164, 1077–1092. [Google Scholar] [CrossRef]
- Navarro, L.; Dunoyer, P.; Jay, F.; Arnold, B.; Dharmasiri, N.; Estelle, M.; Voinnet, O.; Jones, J.D.G. A plant miRNA contributes to antibacterial resistance by repressing auxin signaling. Science 2006, 312, 436–439. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Q.Q.; Zhang, J.G.; Wu, L.; Qi, Y.J.; Zhou, J.M. Identification of MicroRNAs Involved in Pathogen-Associated Molecular Pattern-Triggered Plant Innate Immunity. Plant Physiol. 2010, 152, 2222–2231. [Google Scholar] [CrossRef]
- Zhai, J.X.; Jeong, D.H.; De Paoli, E.; Park, S.; Rosen, B.D.; Li, Y.P.; Gonzalez, A.J.; Yan, Z.; Kitto, S.L.; Grusak, M.A.; et al. MicroRNAs as master regulators of the plant NB-LRR defense gene family via the production of phased, trans-acting siRNAs. Gene Dev. 2011, 25, 2540–2553. [Google Scholar] [CrossRef]
- Zhu, Q.H.; Fan, L.J.; Liu, Y.; Xu, H.; Llewellyn, D.; Wilson, I. miR482 Regulation of NBS-LRR Defense Genes during Fungal Pathogen Infection in Cotton. PLoS ONE 2013, 8, e84390. [Google Scholar] [CrossRef] [PubMed]
- Gerlin, L.; Baroukh, C.; Genin, S. Polyamines: Double agents in disease and plant immunity. Trends Plant Sci. 2021, 26, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Walters, D.R. Polyamines and plant disease. Phytochemistry 2003, 64, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Marina, M.; Maiale, S.J.; Rossi, F.R.; Romero, M.F.; Rivas, E.I.; Garriz, A.; Ruiz, O.A.; Pieckenstain, F.L. Apoplastic polyamine oxidation plays different roles in local responses of tobacco to infection by the necrotrophic fungus Sclerotinia sclerotiorum and the biotrophic bacterium Pseudomonas viridiflava. Plant Physiol. 2008, 147, 2164–2178. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Atanasov, K.E.; Arafaty, N.; Murillo, E.; Tiburcio, A.F.; Zeier, J.; Alcázar, R. Putrescine elicits ROS-dependent activation of the salicylic acid pathway in Arabidopsis thaliana. Plant Cell Environ. 2020, 43, 2755–2768. [Google Scholar] [CrossRef]
- Chávez-Martínez, A.I.; Ortega-Amaro, M.A.; Torres, M.; Serrano, M.; Jiménez-Bremont, J.F. Arabidopsis adc-silenced line exhibits differential defense responses to Botrytis cinerea and Pseudomonas syringae infection. Plant Physiol. Biochem. 2020, 156, 494–503. [Google Scholar] [CrossRef]
- Seifi, H.S.; Shelp, B.J. Spermine Differentially Refines Plant Defense Responses Against Biotic and Abiotic Stresses. Front. Plant Sci. 2019, 10, 117. [Google Scholar] [CrossRef]
- Mo, H.J.; Wang, X.F.; Zhang, Y.; Yang, J.; Ma, Z.Y. Cotton is involved in stem elongation and the plant defense response to through thermospermine alteration. Plant Cell Rep. 2015, 34, 1975–1985. [Google Scholar] [CrossRef]
- Wu, D.S.; von Roepenack-Lahaye, E.; Buntru, M.; de Lange, O.; Schandry, N.; Pérez-Quintero, A.L.; Weinberg, Z.; Lowe-Power, T.M.; Szurek, B.; Michael, A.J.; et al. A Plant Pathogen Type III Effector Protein Subverts Translational Regulation to Boost Host Polyamine Levels. Cell Host Microbe 2019, 26, 638–649. [Google Scholar] [CrossRef]
- Liu, C.X.; Atanasov, K.E.; Tiburcio, A.F.; Alcazar, R. The Polyamine Putrescine Contributes to H2O2 and RbohD/F-Dependent Positive Feedback Loop in Arabidopsis PAMP-Triggered Immunity. Front. Plant Sci. 2019, 10, 894. [Google Scholar] [CrossRef] [PubMed]
- Benkő, P.; Gémes, K.; Fehér, A. Polyamine Oxidase-Generated Reactive Oxygen Species in Plant Development and Adaptation: The Polyamine Oxidase—NADPH Oxidase Nexus. Antioxidants 2022, 11, 2488. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cao, X.L.; Zhu, Y.; Yang, X.M.; Zhang, K.N.; Xiao, Z.Y.; Wang, H.; Zhao, J.H.; Zhang, L.L.; Li, G.B.; et al. Osa-miR398b boosts H2O2 production and rice blast disease-resistance via multiple superoxide dismutases. New Phytol. 2019, 222, 1507–1522. [Google Scholar] [CrossRef]
- Wang, Z.; Xia, Y.; Lin, S.; Wang, Y.; Guo, B.; Song, X.; Ding, S.; Zheng, L.; Feng, R.; Chen, S.; et al. Osa-miR164a targets OsNAC60 and negatively regulates rice immunity against the blast fungus Magnaporthe oryzae. Plant J. 2018, 95, 584–597. [Google Scholar] [CrossRef]
- Zhao, Z.X.; Feng, Q.; Cao, X.L.; Zhu, Y.; Wang, H.; Chandran, V.; Fan, J.; Zhao, J.Q.; Pu, M.; Li, Y.; et al. Facilitates infection of in rice. J. Integr. Plant Biol. 2020, 62, 702–715. [Google Scholar] [CrossRef]
- Wang, H.; Li, Y.; Chern, M.; Zhu, Y.; Zhang, L.L.; Lu, J.H.; Li, X.P.; Dang, W.Q.; Ma, X.C.; Yang, Z.R.; et al. Suppression of rice miR168 improves yield, flowering time and immunity. Nat. Plants 2021, 7, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Li, W.T.; Zhu, Z.W.; Chern, M.S.; Yin, J.J.; Yang, C.; Ran, L.; Cheng, M.P.; He, M.; Wang, K.; Wang, J.; et al. A Natural Allele of a Transcription Factor in Rice Confers Broad-Spectrum Blast Resistance. Cell 2017, 170, 114–126.e15. [Google Scholar] [CrossRef]
- Zhang, X.; Bao, Y.L.; Shan, D.Q.; Wang, Z.H.; Song, X.N.; Wang, Z.Y.; Wang, J.S.; He, L.Q.; Wu, L.; Zhang, Z.G.; et al. Induces the Expression of a MicroRNA to Suppress the Immune Response in Rice. Plant Physiol. 2018, 177, 352–368. [Google Scholar] [CrossRef] [PubMed]
- Chandran, V.; Wang, H.; Gao, F.; Cao, X.-L.; Chen, Y.-P.; Li, G.-B.; Zhu, Y.; Yang, X.-M.; Zhang, L.-L.; Zhao, Z.-X.; et al. miR396-OsGRFs Module Balances Growth and Rice Blast Disease-Resistance. Front. Plant Sci. 2019, 9, 1999. [Google Scholar] [CrossRef]
- Xiao, Z.; Wang, Q.; Zhao, S.; Wang, H.; Li, J.; Fan, J.; Li, Y.; Wang, W.J.A.P.S. MiR444b. 2 regulates resistance to Magnaporthe oryzae and tillering in rice. Acta Phytopathol. Sin. 2017, 47, 511–522. [Google Scholar]
- Li, Y.; Wang, L.F.; Bhutto, S.H.; He, X.R.; Yang, X.M.; Zhou, X.H.; Lin, X.Y.; Rajput, A.A.; Li, G.B.; Zhao, J.H.; et al. Blocking miR530 Improves Rice Resistance, Yield, and Maturity. Front. Plant Sci. 2021, 12, 729560. [Google Scholar] [CrossRef]
- Zhang, L.L.; Huang, Y.Y.; Zheng, Y.P.; Liu, X.X.; Zhou, S.X.; Yang, X.M.; Liu, S.L.; Li, Y.; Li, J.L.; Zhao, S.L.; et al. Osa-miR535 targets SQUAMOSA promoter binding protein-like 4 to regulate blast disease resistance in rice. Plant J. 2022, 110, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, Y.P.; Zhou, X.H.; Yang, X.M.; He, X.R.; Feng, Q.; Zhu, Y.; Li, G.B.; Wang, H.; Zhao, J.H.; et al. Rice miR1432 Fine-Tunes the Balance of Yield and Blast Disease Resistance via Different Modules. Rice 2021, 14, 87. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, T.T.; He, X.R.; Zhu, Y.; Feng, Q.; Yang, X.M.; Zhou, X.H.; Li, G.B.; Ji, Y.P.; Zhao, J.H.; et al. Blocking Osa-miR1871 enhances rice resistance against Magnaporthe oryzae and yield. Plant Biotechnol. J. 2022, 20, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.X.; Zhu, Y.; Wang, L.F.; Zheng, Y.P.; Chen, J.F.; Li, T.T.; Yang, X.M.; Wang, H.; Li, X.P.; Ma, X.C.; et al. Fine-tunes rice immunity against and yield traits. J. Integr. Plant Biol. 2020, 62, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.F.; Zhao, Z.X.; Li, Y.; Li, T.T.; Zhu, Y.; Yang, X.M.; Zhou, S.X.; Wang, H.; Zhao, J.Q.; Pu, M.; et al. Fine-Tuning Roles of Osa-miR159a in Rice Immunity Against Magnaporthe oryzae and Development. Rice 2021, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Wang, H.; Yang, X.M.; Hu, Z.W.; Zhou, X.H.; Xiang, L.; Xiong, X.Y.; He, X.R.; Zhu, Y.; Li, G.B.; et al. Osa-miR160a confers broad-spectrum resistance to fungal and bacterial pathogens in rice. New Phytol. 2022, 236, 2216–2232. [Google Scholar] [CrossRef]
- Li, X.-P.; Ma, X.-C.; Wang, H.; Zhu, Y.; Liu, X.-X.; Li, T.-T.; Zheng, Y.-P.; Zhao, J.-Q.; Zhang, J.-W.; Huang, Y.-Y.; et al. Osa-miR162a fine-tunes rice resistance to Magnaporthe oryzae and Yield. Rice 2020, 13, 38. [Google Scholar] [CrossRef]
- Salvador-Guirao, R.; Hsing, Y.I.; Segundo, B.S. The Polycistronic miR166k-166h Positively Regulates Rice Immunity via Post-transcriptional Control of EIN2. Front. Plant Sci. 2018, 9, 337. [Google Scholar] [CrossRef]
- Sánchez-Sanuy, F.; Peris-Peris, C.; Tomiyama, S.; Okada, K.; Hsing, Y.I.; San Segundo, B.; Campo, S. Enhances transcriptional priming in defense responses against the rice blast fungus. BMC Plant Biol. 2019, 19, 563. [Google Scholar] [CrossRef]
- Dong, S.Y.; Zhang, J.X.; Sun, D.Y.; Liu, H.; Yang, Q.Y.; Wang, H.; Chen, Z.Q.; Wang, J.F. Identification of Magnaporthe oryzae-elicited rice novel miRNAs and their targets by miRNA and degradome sequencing. Eur. J. Plant Pathol. 2018, 151, 629–647. [Google Scholar] [CrossRef]
- Markham, N.R.; Zuker, M. UNAFold: Software for nucleic acid folding and hybridization. In Bioinformatics; Methods in Molecular Biology; Keith, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2008; pp. 3–31. [Google Scholar]
- Lv, Y.S.; Shao, G.N.; Jiao, G.A.; Sheng, Z.H.; Xie, L.H.; Hu, S.K.; Tang, S.Q.; Wei, X.J.; Hu, P.S. Targeted mutagenesis of POLYAMINE OXIDASE 5 that negatively regulates mesocotyl elongation enables the generation of direct-seeding rice with improved grain yield. Mol. Plant 2021, 14, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Kim, D.W.; Watanabe, K.; Sasaki, A.; Niitsu, M.; Berberich, T.; Kusano, T.; Takahashi, Y. Constitutively and highly expressed Oryza sativa polyamine oxidases localize in peroxisomes and catalyze polyamine back conversion. Amino Acids 2012, 42, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Junhua, L.; Xuemei, Y.; Jinfeng, C.; Tingting, L.; Zijin, H.; Ying, X.; Jinlu, L.; Jiqun, Z.; Mei, P.; Hui, F.; et al. Osa-miR439 Negatively Regulates Rice Immunity Against Magnaporthe oryzae. Rice Sci. 2021, 28, 156–165. [Google Scholar] [CrossRef]
- Shang, R.F.; Lee, S.; Senavirathne, G.; Lai, E.C. microRNAs in action: Biogenesis, function and regulation. Nat. Rev. Genet. 2023, 1–18. [Google Scholar] [CrossRef]
- Jia, Y.F.; Li, Q.L.; Li, Y.Y.; Zhai, W.X.; Jiang, G.H.; Li, C.R. Inducible Enrichment of Osa-miR1432 Confers Rice Bacterial Blight Resistance through Suppressing OsCaML2. Int. J. Mol. Sci. 2021, 22, 11367. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; He, X.W.; Wang, X.X.; Zhang, S.X.; Guo, X.Q. ghr-miR5272a-mediated regulation of gene transcription contributes to the immune response in cotton. J. Exp. Bot. 2017, 68, 5895–5906. [Google Scholar] [CrossRef]
- Casero, R.A.; Murray Stewart, T.; Pegg, A.E. Polyamine metabolism and cancer: Treatments, challenges and opportunities. Nat. Rev. Cancer 2018, 18, 681–695. [Google Scholar] [CrossRef]
- Fincato, P.; Moschou, P.N.; Spedaletti, V.; Tavazza, R.; Angelini, R.; Federico, R.; Roubelakis-Angelakis, K.A.; Tavladoraki, P. Functional diversity inside the gene family. J. Exp. Bot. 2011, 62, 1155–1168. [Google Scholar] [CrossRef]
- Seifi, H.S.; Zarei, A.; Hsiang, T.; Shelp, B.J. Spermine Is a Potent Plant Defense Activator Against Gray Mold Disease on Solanum lycopersicum, Phaseolus vulgaris, and Arabidopsis thaliana. Phytopathology 2019, 109, 1367–1377. [Google Scholar] [CrossRef]
- Pandey, S.; Ranade, S.A.; Nagar, P.K.; Kumar, N. Role of polyamines and ethylene as modulators of plant senescence. J. Biosci. 2000, 25, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.W.; Zhang, J.F.; Quan, R.D.; Pan, X.W.; Wan, L.Y.; Huang, R.F. EAR motif mutation of rice OsERF3 alters the regulation of ethylene biosynthesis and drought tolerance. Planta 2013, 237, 1443–1451. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Dong, S.Y.; Li, M.; Gu, F.W.; Yang, G.L.; Guo, T.; Chen, Z.Q.; Wang, J.F. The Class III peroxidase gene OsPrx30, transcriptionally modulated by the AT-hook protein OsATH1, mediates rice bacterial blight-induced ROS accumulation. J. Integr. Plant Biol. 2021, 63, 393–408. [Google Scholar] [CrossRef]
- Liu, H.; Dong, S.Y.; Gu, F.W.; Liu, W.; Yang, G.L.; Huang, M.; Xiao, W.M.; Liu, Y.Z.; Guo, T.; Wang, H.; et al. NBS-LRR Protein Pik-H4 Interacts with OsBIHD1 to Balance Rice Blast Resistance and Growth by Coordinating Ethylene-Brassinosteroid Pathway. Front. Plant Sci. 2017, 8, 127. [Google Scholar] [CrossRef] [PubMed]
- Park, C.-H.; Chen, S.; Shirsekar, G.; Zhou, B.; Khang, C.H.; Songkumarn, P.; Afzal, A.J.; Ning, Y.; Wang, R.; Bellizzi, M.; et al. The Magnaporthe oryzae Effector AvrPiz-t Targets the RING E3 Ubiquitin Ligase APIP6 to Suppress Pathogen-Associated Molecular Pattern–Triggered Immunity in Rice. Plant Cell 2012, 24, 4748–4762. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Varkonyi-Gasic, E.; Wu, R.M.; Wood, M.; Walton, E.F.; Hellens, R.P. Protocol: A highly sensitive RT-PCR method for detection and quantification of microRNAs. Plant Methods 2007, 3, 1–12. [Google Scholar] [CrossRef]
- Zapata, P.J.; Serrano, M.; Pretel, M.T.; Amoros, A.; Botella, A. Changes in ethylene evolution and polyamine profiles of seedlings of nine cultivars of Lactuca sativa L. in response to salt stress during germination. Plant Sci. 2003, 164, 557–563. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, S.; Hou, Y.; Huang, Q.; Wu, P.; Han, Z.; Wei, D.; Xie, H.; Gu, F.; Chen, C.; Wang, J. Osa-miR11117 Targets OsPAO4 to Regulate Rice Immunity against the Blast Fungus Magnaporthe oryzae. Int. J. Mol. Sci. 2023, 24, 16052. https://doi.org/10.3390/ijms242216052
Gao S, Hou Y, Huang Q, Wu P, Han Z, Wei D, Xie H, Gu F, Chen C, Wang J. Osa-miR11117 Targets OsPAO4 to Regulate Rice Immunity against the Blast Fungus Magnaporthe oryzae. International Journal of Molecular Sciences. 2023; 24(22):16052. https://doi.org/10.3390/ijms242216052
Chicago/Turabian StyleGao, Shang, Yan Hou, Qiwei Huang, Pengzhi Wu, Zhikai Han, Danhong Wei, Huabin Xie, Fengwei Gu, Chun Chen, and Jiafeng Wang. 2023. "Osa-miR11117 Targets OsPAO4 to Regulate Rice Immunity against the Blast Fungus Magnaporthe oryzae" International Journal of Molecular Sciences 24, no. 22: 16052. https://doi.org/10.3390/ijms242216052