
Genome-Wide Identification and Expression Analysis of the Copper Transporter (COPT/Ctr) Gene Family in Kandelia obovata, a Typical Mangrove Plant

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Copper Treatments Affect the Growth of Kandelia obovata Seedlings
2.2. Identification of COPT Family Members in Kandelia obovata
2.3. Variation across the COPT Family in Terms of Domain and 3D Structure
2.4. COPT Protein Phylogenetic Relationships
2.5. COPT Gene Structure and Conserved Motif Investigation
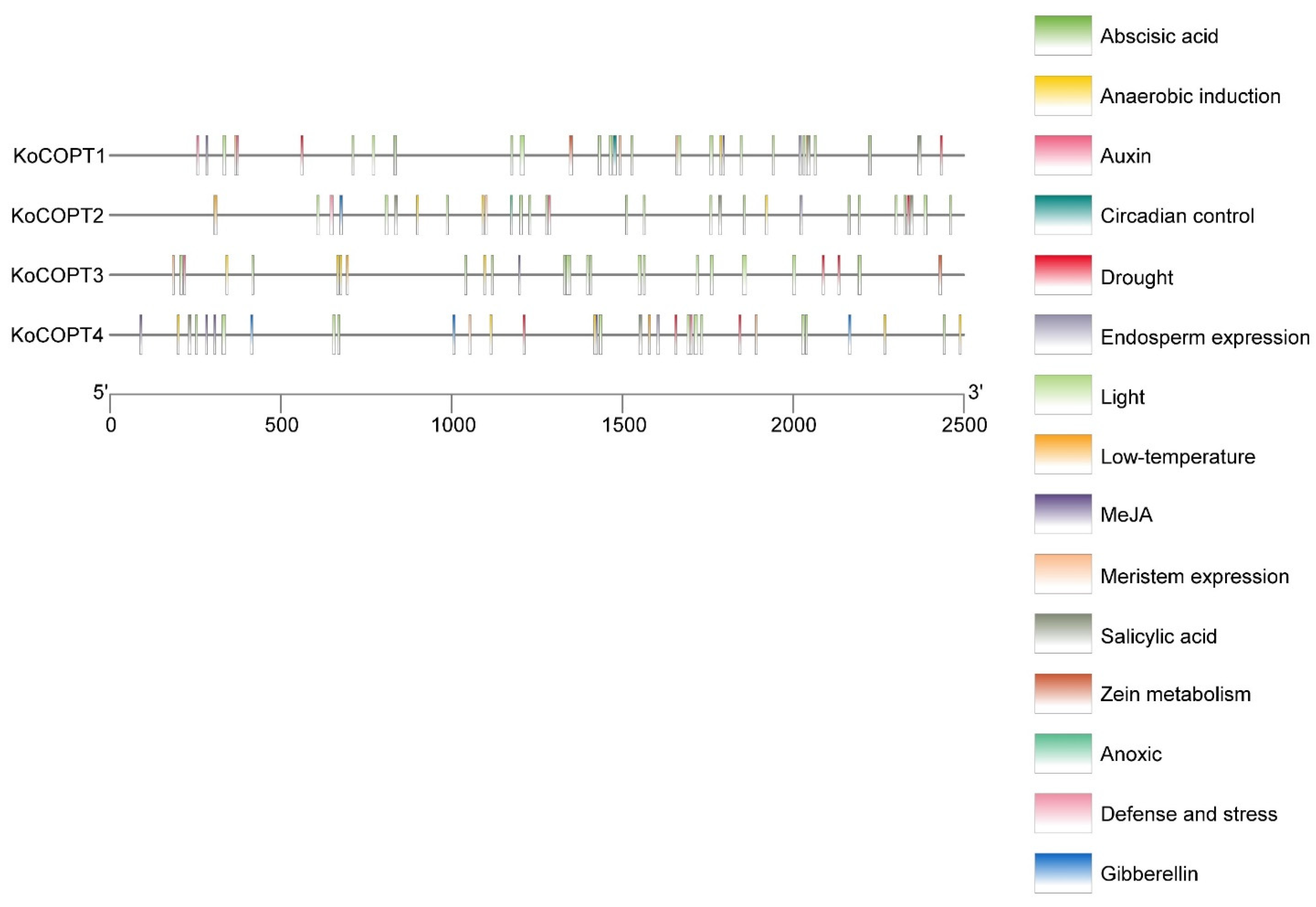
2.6. Identification of Cis-Regulatory Elements in the Promoters of Four COPT Genes
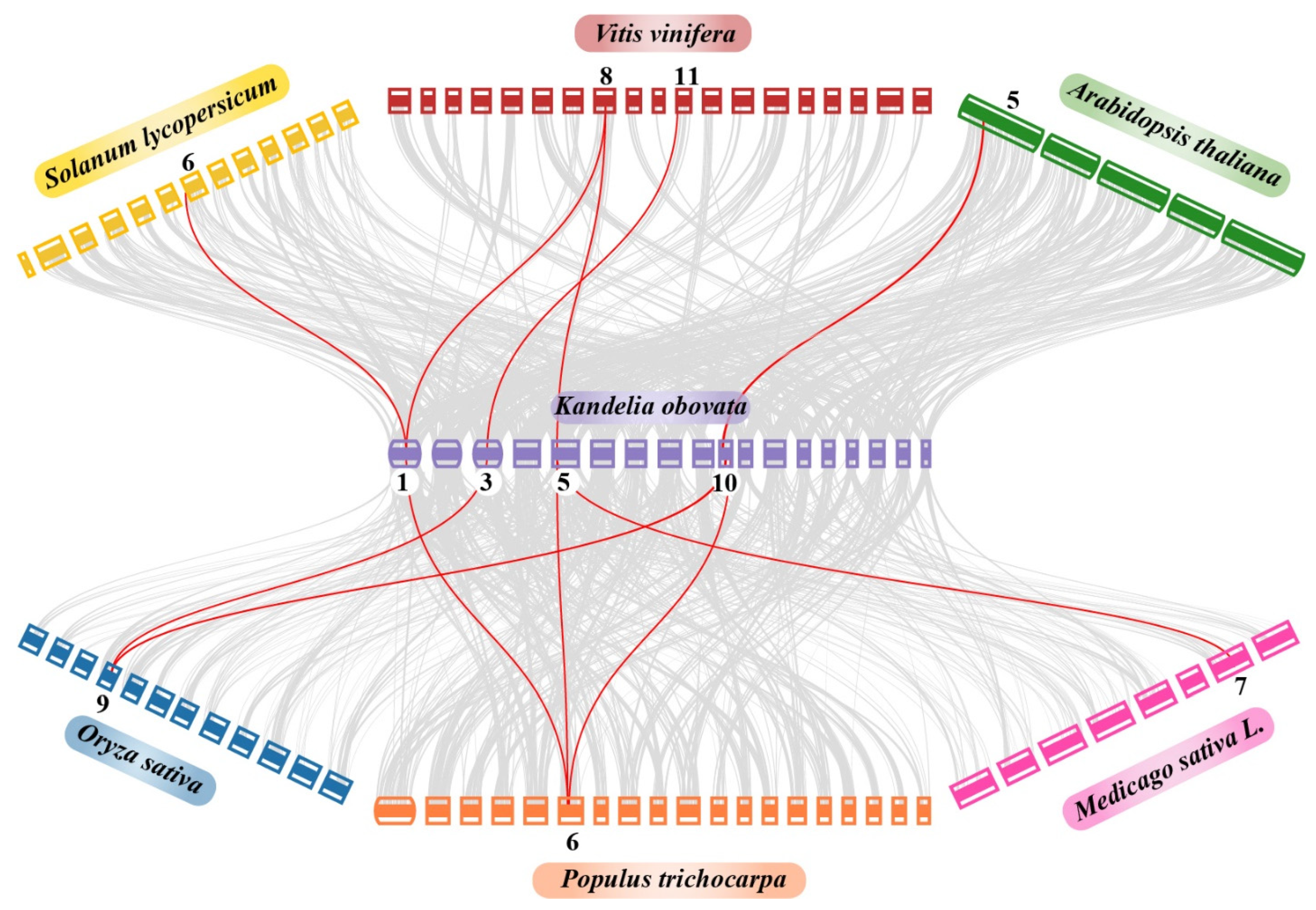
2.7. Synteny and Duplication Analysis of the COPT Family
2.8. Expression Analysis of KoCOPT Genes under Different Copper Stress
3. Discussion
4. Materials and Methods
4.1. Identification and Characterization of COPT Genes in Kandelia obovata
4.2. Chromosomal Distribution of COPT Genes in Kandelia obovata
4.3. Phylogenetic Tree Construction
4.4. Gene Structure and Significant Motif Analyses of the COPT Family Members
4.5. Analysis of the COPT Family’s Promoter Sequences in Kandelia obovata
4.6. Synteny and Duplication Analysis
4.7. Three-Dimensional Structure and Subcellular Localization
- (1)
- ProtComp 9.0: http://linux1.softberry.com/berry.phtml?topic=protcomppl&group=programs&subgroup=proloc (accessed on 8 April 2023);
- (2)
- CELLO server: cello.life.nctu.edu.tw/, accessed on 8 April 2023).
4.8. Plant Material and Environmental Conditions
4.9. Quantitative Real-Time PCR Assays
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, M.; Li, X.; Xiao, J.; Wang, S. Molecular and Functional Analyses of COPT/Ctr-Type Copper Transporter-like Gene Family in Rice. BMC Plant Biol. 2011, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Fu, Q.; Zheng, J.; Zhang, A.; Wang, H. Transcriptomic and Metabolomic Analyses Reveal That Melatonin Promotes Melon Root Development under Copper Stress by Inhibiting Jasmonic Acid Biosynthesis. Hortic. Res. 2020, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Cano-Gauci, D.F.; Sarkar, B. Reversible Zinc Exchange between Metallothionein and the Estrogen Receptor Zinc Finger. FEBS Lett. 1996, 386, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Peñarrubia, L.; Andrés-Colás, N.; Moreno, J.; Puig, S. Regulation of Copper Transport in Arabidopsis Thaliana: A Biochemical Oscillator? J. Biol. Inorg. Chem. 2010, 15, 29–36. [Google Scholar] [CrossRef]
- Burkhead, J.L.; Gogolin Reynolds, K.A.; Abdel-Ghany, S.E.; Cohu, C.M.; Pilon, M. Copper Homeostasis. New Phytol. 2009, 182, 799–816. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, J.; Wang, W.; Li, D.; Hu, X.; Wang, H.; Wei, M.; Liu, Q.; Wang, Z.; Li, C. Genome-Wide Identification and Expression Profiling of the Copper Transporter Gene Family in Populus Trichocarpa. Plant Physiol. Biochem. 2015, 97, 451–460. [Google Scholar] [CrossRef]
- Puig, S.; Thiele, D.J. Molecular Mechanisms of Copper Uptake and Distribution. Curr. Opin. Chem. Biol. 2002, 6, 171–180. [Google Scholar] [CrossRef]
- Beaudoin, J.; Thiele, D.J.; Labbé, S.; Puig, S. Dissection of the Relative Contribution of the Schizosaccharomyces Pombe Ctr4 and Ctr5 Proteins to the Copper Transport and Cell Surface Delivery Functions. Microbiology 2011, 157, 1021–1031. [Google Scholar] [CrossRef]
- De Feo, C.J.; Aller, S.G.; Siluvai, G.S.; Blackburn, N.J.; Unger, V.M. Three-Dimensional Structure of the Human Copper Transporter HCTR1. Proc. Natl. Acad. Sci. USA 2009, 106, 4237–4242. [Google Scholar] [CrossRef]
- Dancis, A.; Yuan, D.S.; Haile, D.; Askwith, C.; Eide, D.; Moehle, C.; Kaplan, J.; Klausner, R.D. Molecular Characterization of a Copper Transport Protein in S. Cerevisiae: An Unexpected Role for Copper in Iron Transport. Cell 1994, 76, 393–402. [Google Scholar] [CrossRef]
- Labbé, S.; Thiele, D.J. Pipes and Wiring: The Regulation of Copper Uptake and Distribution in Yeast. Trends Microbiol. 1999, 7, 500–505. [Google Scholar] [CrossRef] [PubMed]
- Sancenón, V.; Puig, S.; Mira, H.; Thiele, D.J.; Peñarrubia, L. Identification of a Copper Transporter Family in Arabidopsis Thaliana. Plant Mol. Biol. 2003, 51, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Sancenón, V.; Puig, S.; Mateu-Andrés, I.; Dorcey, E.; Thiele, D.J.; Peñarrubia, L. The Arabidopsis Copper Transporter COPT1 Functions in Root Elongation and Pollen Development. J. Biol. Chem. 2004, 279, 15348–15355. [Google Scholar] [CrossRef] [PubMed]
- Perea-García, A.; Garcia-Molina, A.; Andrés-Colás, N.; Vera-Sirera, F.; Pérez-Amador, M.A.; Puig, S.; Peñarrubia, L. Arabidopsis Copper Transport Protein COPT2 Participates in the Cross Talk between Iron Deficiency Responses and Low-Phosphate Signaling. Plant Physiol. 2013, 162, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Klaumann, S.; Nickolaus, S.D.; Fürst, S.H.; Starck, S.; Schneider, S.; Ekkehard Neuhaus, H.; Trentmann, O. The Tonoplast Copper Transporter COPT5 Acts as an Exporter and Is Required for Interorgan Allocation of Copper in Arabidopsis Thaliana. New Phytol. 2011, 192, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Molina, A.; Andrés-Colás, N.; Perea-García, A.; Neumann, U.; Dodani, S.C.; Huijser, P.; Peñarrubia, L.; Puig, S. The Arabidopsis COPT6 Transport Protein Functions in Copper Distribution under Copper-Deficient Conditions. Plant Cell Physiol. 2013, 54, 1378–1390. [Google Scholar] [CrossRef]
- Martins, V.; Hanana, M.; Blumwald, E.; Gerós, H. Copper Transport and Compartmentation in Grape Cells. Plant Cell Physiol. 2012, 53, 1866–1880. [Google Scholar] [CrossRef]
- Martins, V.; Bassil, E.; Hanana, M.; Blumwald, E.; Gerós, H. Copper Homeostasis in Grapevine: Functional Characterization of the Vitis Vinifera Copper Transporter 1. Planta 2014, 240, 91–101. [Google Scholar] [CrossRef]
- Nizam, A.; Meera, S.P.; Kumar, A. Genetic and Molecular Mechanisms Underlying Mangrove Adaptations to Intertidal Environments. iScience 2022, 25, 103547. [Google Scholar] [CrossRef]
- Krauss, K.W.; Mckee, K.L.; Lovelock, C.E.; Cahoon, D.R.; Saintilan, N.; Reef, R.; Chen, L. How Mangrove Forests Adjust to Rising Sea Level. New Phytol. 2014, 202, 19–34. [Google Scholar] [CrossRef]
- Sun, M.M.; Liu, X.; Huang, X.J.; Yang, J.J.; Qin, P.T.; Zhou, H.; Jiang, M.G.; Liao, H.Z. Genome-Wide Identification and Expression Analysis of the NAC Gene Family in Kandelia Obovata, a Typical Mangrove Plant. Curr. Issues Mol. Biol. 2022, 44, 5622–5637. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.J.; Sun, W.H.; Tsai, W.C.; Xiang, S.; Lai, X.K.; Chen, D.Q.; Liu, X.D.; Wang, Y.F.; Le, Y.X.; Chen, S.M.; et al. Chromosome-Scale Assembly of the Kandelia Obovata Genome. Hortic. Res. 2020, 7, 75. [Google Scholar] [CrossRef] [PubMed]
- Sheue, C.R.; Liu, H.Y.; Yong, J.W.H. Kandelia Obovata (Rhizophoraceae), a New Mangrove Species from Eastern Asia. Taxon 2003, 52, 287–294. [Google Scholar] [CrossRef]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and Distribution of Mangrove Forests of the World Using Earth Observation Satellite Data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Wardiatno, Y.; Mardiansyah; Prartono, T.; Tsuchiya, M. Possible Food Sources of Macrozoobenthos in the Manko Mangrove Ecosystem, Okinawa (Japan): A Stable Isotope Analysis Approach. Trop. Life Sci. Res. 2015, 26, 53–65. [Google Scholar]
- Zhou, Q.; Tu, C.; Fu, C.; Li, Y.; Zhang, H.; Xiong, K.; Zhao, X.; Li, L.; Waniek, J.J.; Luo, Y. Characteristics and Distribution of Microplastics in the Coastal Mangrove Sediments of China. Sci. Total Environ. 2020, 703, 134807. [Google Scholar] [CrossRef]
- Li, T.; Ye, Y. Dynamics of Decomposition and Nutrient Release of Leaf Litter in Kandelia Obovata Mangrove Forests with Different Ages in Jiulongjiang Estuary, China. Ecol. Eng. 2014, 73, 454–460. [Google Scholar] [CrossRef]
- Wang, C.W.; Wong, S.L.; Liao, T.S.; Weng, J.H.; Chen, M.N.; Huang, M.Y.; Chen, C.I. Photosynthesis in Response to Salinity and Submergence in Two Rhizophoraceae Mangroves Adapted to Different Tidal Elevations. Tree Physiol. 2022, 42, 1016–1028. [Google Scholar] [CrossRef]
- Shen, X.; Li, R.; Chai, M.; Cheng, S.; Tam, N.F.Y.; Han, J. Does Combined Heavy Metal Stress Enhance Iron Plaque Formation and Heavy Metal Bioaccumulation in Kandelia Obovata? Environ. Exp. Bot. 2021, 186, 104463. [Google Scholar] [CrossRef]
- Bodin, N.; N’Gom-Kâ, R.; Kâ, S.; Thiaw, O.T.; Tito de Morais, L.; Le Loc’h, F.; Rozuel-Chartier, E.; Auger, D.; Chiffoleau, J.F. Assessment of Trace Metal Contamination in Mangrove Ecosystems from Senegal, West Africa. Chemosphere 2013, 90, 150–157. [Google Scholar] [CrossRef]
- Abubakar, U.S.; Zulkifli, S.Z.; Ismail, A. Heavy Metals Bioavailability and Pollution Indices Evaluation in the Mangrove Surface Sediment of Sungai Puloh, Malaysia. Environ. Earth Sci. 2018, 77, 225. [Google Scholar] [CrossRef]
- Cheng, S.; Tam, N.F.Y.; Li, R.; Shen, X.; Niu, Z.; Chai, M.; Qiu, G.Y. Temporal Variations in Physiological Responses of Kandelia Obovata Seedlings Exposed to Multiple Heavy Metals. Mar. Pollut. Bull. 2017, 124, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Wenjiao, Z. Effects of Zinc Stress on Growth and Antioxidant Enzyme Responses of Kandelia Obovata Seedlings. Toxicol. Environ. Chem. 2015, 97, 1190–1201. [Google Scholar] [CrossRef]
- Shen, X.; Li, R.; Chai, M.; Cheng, S.; Niu, Z.; Qiu, G.Y. Interactive Effects of Single, Binary and Trinary Trace Metals (Lead, Zinc and Copper) on the Physiological Responses of Kandelia Obovata Seedlings. Environ. Geochem. Health 2019, 41, 135–148. [Google Scholar] [CrossRef]
- Huang, G.Y.; Wang, Y.S. Physiological and Biochemical Responses in the Leaves of Two Mangrove Plant Seedlings (Kandelia Candel and Bruguiera Gymnorrhiza) Exposed to Multiple Heavy Metals. J. Hazard. Mater. 2010, 182, 848–854. [Google Scholar] [CrossRef]
- Sanz, A.; Pike, S.; Khan, M.A.; Carrió-Seguí, À.; Mendoza-Cózatl, D.G.; Peñarrubia, L.; Gassmann, W. Copper Uptake Mechanism of Arabidopsis Thaliana High-Affinity COPT Transporters. Protoplasma 2019, 256, 161–170. [Google Scholar] [CrossRef]
- Wang, Q.; Wei, N.; Jin, X.; Min, X.; Ma, Y.; Liu, W. Molecular Characterization of the COPT/Ctr-Type Copper Transporter Family under Heavy Metal Stress in Alfalfa. Int. J. Biol. Macromol. 2021, 181, 644–652. [Google Scholar] [CrossRef]
- Wang, H.; Du, H.; Li, H.; Huang, Y.; Ding, J.; Liu, C.; Wang, N.; Lan, H.; Zhang, S. Identification and Functional Characterization of the ZmCOPT Copper Transporter Family in Maize. PLoS ONE 2018, 13, e0199081. [Google Scholar] [CrossRef]
- Chai, M.; Li, R.; Shen, X.; Yu, L.; Han, J. Multiple Heavy Metals Affect Root Response, Iron Plaque Formation, and Metal Bioaccumulation of Kandelia Obovata. Sci. Rep. 2022, 12, 14389. [Google Scholar] [CrossRef]
- Hirokawa, T.; Boon-Chieng, S.; Mitaku, S. SOSUI: Classification and Secondary Structure Prediction System for Membrane Proteins. Bioinformatics 1998, 14, 378–379. [Google Scholar] [CrossRef]
- Yaghobi, M.; Heidari, P. Genome-Wide Analysis of Aquaporin Gene Family in Triticum Turgidum and Its Expression Profile in Response to Salt Stress. Genes 2023, 14, 202. [Google Scholar] [CrossRef] [PubMed]
- Hashemipetroudi, S.H.; Arab, M. Genome-Wide Analysis of the Laccase (LAC) Gene Family in Aeluropus Littoralis: A Focus on Identi Fi Cation, Evolution and Expression Patterns in Response to Abiotic Stresses and ABA Treatment. Front. Plant Sci. 2023, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hanada, K.; Zou, C.; Lehti-Shiu, M.D.; Shinozaki, K.; Shiu, S.H. Importance of Lineage-Specific Expansion of Plant Tandem Duplicates in the Adaptive Response to Environmental Stimuli. Plant Physiol. 2008, 148, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Q.; Zheng, M.; Chang, W.; Ashraf, M.F.; Khan, R.; Asim, M.; Riaz, M.W.; Alwahibi, M.S.; Elshikh, M.S.; Zhang, R.; et al. Genome-Wide Identification and Expression Analysis of SnRK2 Gene Family in Dormant Vegetative Buds of Liriodendron Chinense in Response to Abscisic Acid, Chilling, and Photoperiod. Genes 2022, 13, 1305. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Q.; Zheng, M.; Furqan, M.; Khan, R.; Yasir, M.; Farooq, S.; Zhang, R.; Wu, J. Genome-Wide Identification, Characterization and Expression Analysis of the ABA Receptor PYL Gene Family in Response to ABA, Photoperiod, and Chilling in Vegetative Buds of Liriodendron Chinense. Sci. Hortic. 2022, 303, 111200. [Google Scholar] [CrossRef]
- Asim, M.; Hussain, Q.; Wang, X.; Sun, Y.; Liu, H.; Khan, R.; Du, S.; Shi, Y.; Zhang, Y. Mathematical Modeling Reveals That Sucrose Regulates Leaf Senescence via Dynamic Sugar Signaling Pathways. Int. J. Mol. Sci. 2022, 23, 6498. [Google Scholar] [CrossRef]
- Khan, R.; Ma, X.; Hussain, Q.; Asim, M.; Iqbal, A.; Ren, X.; Shah, S.; Chen, K.; Shi, Y. Application of 2,4-Epibrassinolide Improves Drought Tolerance in Tobacco through Physiological and Biochemical Mechanisms. Biology 2022, 11, 1192. [Google Scholar] [CrossRef]
- Sajjad, M.; Ahmad, A.; Riaz, M.W.; Hussain, Q.; Yasir, M.; Lu, M. Recent Genome Resequencing Paraded COBRA-Like Gene Family Roles in Abiotic Stress and Wood Formation in Poplar. Front. Plant Sci. 2023, 14, 1–20. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Location | AA 1 | Chains 2 | MW 3/kDd | pI 4 | GRAVY 5 | Aliphatic Index | Instability |
|---|---|---|---|---|---|---|---|---|---|
| KoCOPT1 | geneMaker00016256 | Chr1 8617182–8617661 | 159 | + | 17.09 | 9.72 | 0.309 | 82.2 | 47.68 |
| KoCOPT2 | geneMaker00000230 | Chr3 5129069–5129389 | 106 | - | 11.18 | 8.71 | 0.498 | 105.85 | 48.23 |
| KoCOPT3 | geneMaker00004869 | Chr5 1473358–1473795 | 145 | - | 15.64 | 6.82 | 0.566 | 88.83 | 34.84 |
| KoCOPT4 | geneMaker00015010 | Chr10 3212193–3212594 | 133 | - | 14.78 | 8.82 | 0.805 | 124.66 | 49.66 |
| Name | Method | Ka | Ks | Ka/Ks | Divergence-Time (MYA) | Duplicated type |
|---|---|---|---|---|---|---|
| KoCOPT1 & KoCOP3 | MS | 1.06 | 0.86 | 1.23 | 28.65 | Segmental |
| Gene Name | Primer Name | Sequence (5′ to 3′) |
|---|---|---|
| KoCOPT1 | Forward | AAGATGATGCCACACCTCGG |
| Reverse | CAACCCGGGAAGAGGAACTC | |
| KoCOPT2 | Forward | AACGCCGCAATCGGTTATTT |
| Reverse | CAAATCGACAACCTCGCCATC | |
| KoCOPT3 | Forward | CTTCAGCCAAGTCACCGTCT |
| Reverse | AGGAACGGTGCTATTTCGGT | |
| KoCOPT4 | Forward | CACTTTGCTCGCCTGTTTCC |
| Reverse | ACCCAATTGCCAGTCCTACG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, Q.; Ye, T.; Li, S.; Nkoh, J.N.; Zhou, Q.; Shang, C. Genome-Wide Identification and Expression Analysis of the Copper Transporter (COPT/Ctr) Gene Family in Kandelia obovata, a Typical Mangrove Plant. Int. J. Mol. Sci. 2023, 24, 15579. https://doi.org/10.3390/ijms242115579
Hussain Q, Ye T, Li S, Nkoh JN, Zhou Q, Shang C. Genome-Wide Identification and Expression Analysis of the Copper Transporter (COPT/Ctr) Gene Family in Kandelia obovata, a Typical Mangrove Plant. International Journal of Molecular Sciences. 2023; 24(21):15579. https://doi.org/10.3390/ijms242115579
Chicago/Turabian StyleHussain, Quaid, Ting Ye, Sihui Li, Jackson Nkoh Nkoh, Qiao Zhou, and Chenjing Shang. 2023. "Genome-Wide Identification and Expression Analysis of the Copper Transporter (COPT/Ctr) Gene Family in Kandelia obovata, a Typical Mangrove Plant" International Journal of Molecular Sciences 24, no. 21: 15579. https://doi.org/10.3390/ijms242115579