
Searching for Atherosclerosis Biomarkers by Proteomics: A Focus on Lesion Pathogenesis and Vulnerability



Abstract
:1. Introduction

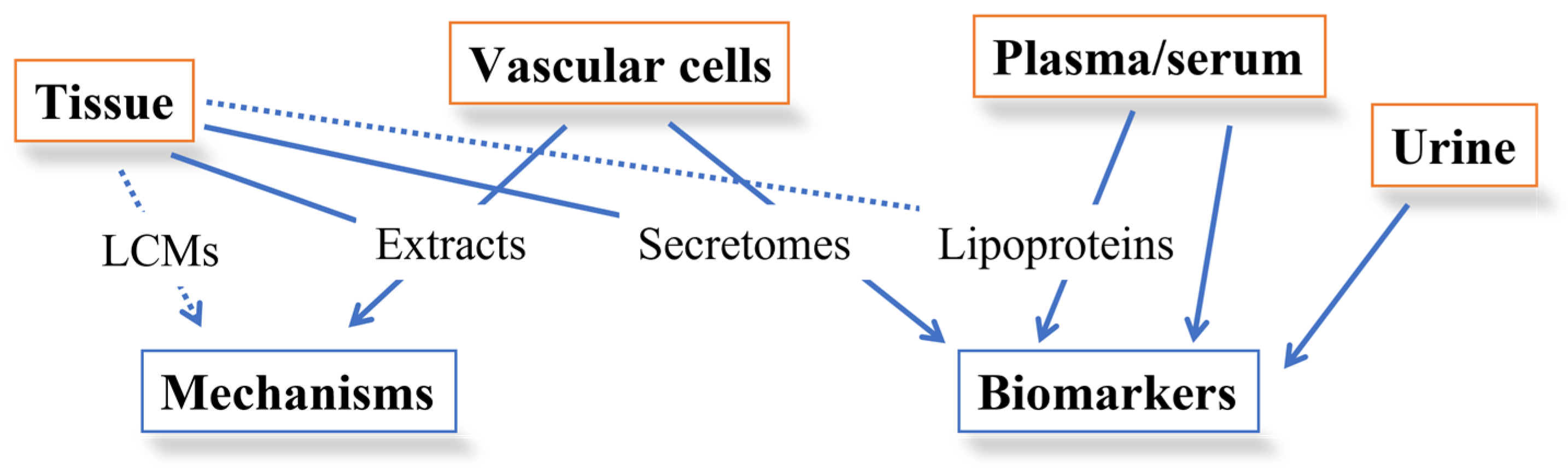
2. Application of MS-Based Technologies to the Study of Lipoproteins Involvement in Atherogenesis
2.1. Lipoproteomics
2.2. Lipidomics
3. Atherosclerotic Plaque Dissection through Proteomics
3.1. Studies on Human Plaque Extracts
3.2. Studies on Human Plaque Secretomes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Proteomics | Main Findings | References |
|---|---|---|---|
| Secretome from normal carotid artery segments vs. non complicated plaque vs. complicated plaque with thrombus | 2DE coupled with MALDI TOF MS | The more complicated the lesion, the higher the number of secreted proteins, suggesting the production of specific proteins relating to the complexity of the atherosclerotic lesion | [175] |
| Secretome from 35 atherosclerotic endarterectomies (10 femoral, 25 carotids) vs. 36 control endarteries (24 mammary, 12 radial) | 2DE coupled with LC-MS/MS | Decreased secretion of HSP27 in complicated atherosclerotic plaques | [176] |
| 21 carotid arteries secretomes—stenosing complicated zone (incubated in the presence or absence of atorvastatin) vs. the adjacent fibrous zone | 2DE coupled with MALDI TOF MS/Q TOF MS | ↑ 24 proteins ↓ 20 proteins; 66% of the proteins differentially released by atherosclerotic plaques reverted to control values after the administration of atorvastatin | [177] |
| Secretomes from 7 carotid arteries vs. 7 mammary arteries | SELDI TOF MS | ↓ soluble tumor necrosis factor-like weak inducer of apoptosis (sTWEAK); sTWEAK concentrations negatively correlated with the carotid intima-media thickness (r 0.4; p 0.001) | [178] |
| Secretomes from three biological replicates of human atherosclerotic coronary arteries (APC), preatherosclerotic coronaries (PC) and mammaries (M) | 1DE coupled with LC MS/MS | In total, 64 proteins were identified in the 3 replicates of at least one of the 3 groups and 15 secreted proteins have not been previously reported in plasma. Four proteins were significantly released in higher amounts by mammary tissue: gelsolin, vinculin, lamin A/C and phosphoglucomutase 5 | [179] |
| Secretomes from carotid endarterectomy specimens of 14 patients | LC MS/MS | In total, 463 proteins were identified, of which 31 proteins were differentially secreted between plaques and the corresponding downstream segments | [21] |
| Secretomes from 12 carotid atherosclerotic plaque vs. 10 nonatherosclerotic mammary artery | Tandem immunoaffinity depletion, iTRAQ labeling, and nano LC-MS/MS | In total, 162 proteins were quantified, of which 25 were differentially expressed in secretome between carotid atherosclerotic plaque and non-diseased mammary artery. ↑ neutrophil defensin 1, apolipoprotein E, clusterin, and zinc-alpha-2-glycoprotein in CAP secretomes | [180] |
3.3. Studies on Animal Models
| Samples | Proteomics | Main Findings | References |
|---|---|---|---|
| Aortas from 10-week-old, 12-month-old, and 18-month-old apolipoprotein E-/- mice | 2DE coupled with LC-MS/MS | In total, 79 protein species were identified as altered during various stages of atherogenesis. Immunoglobulin deposition, redox imbalance, and impaired energy metabolism preceded lesion formation in apolipoprotein E-/- mice | [29] |
| Atherosclerotic aorta vs. normal aorta from Wistar rats (Atherosclerosis was induced by a single dose of vitamin D3 followed by a high fat diet) | 2DE coupled with MALDI TOF MS | Found 46 differentially expressed proteins | [183] |
| Aorta from apo E-/- mice vs. WT mice | Affinity proteomic strategy for in situ isolation and differential mapping of vascular endothelial proteins: perfusion with biotin, capturing with streptavidin chromatography, SDS-PAGE followed by LC-MS/MS | Here, 454 proteins identified, and 81 differentially expressed proteins involved in immune and inflammatory responses, cell adhesion, and lipid metabolism | [30] |
| Rat aorta cultured under two levels of shear stress, 5 and 15 dyn/cm2 | 2DE coupled with MALDI TOF MS | ↓ Rho-GDIα in low shear stress vessels | [185] |
| Aortas from Lipoprotein(a) transgenic mice vs WT mice | 2-DE followed by MALDI-TOF/TOF MS | Here, 34 differentially expressed proteins found involved in energy, redox, and lipid metabolism | [186] |
| Ascending arch of thoracic aorta from 24 male albino rabbits assigned randomly into 4 groups: control, cholesterol, cholesterol plus vitamin E and vitamin E groups | LC MS/MS | Differential expression of proteins following cholesterol and also vitamin E treatments | [33] |
| Secretomes of coronary arteries from pigs fed on standard or high-cholesterol diet | LC MS/MS | TNFα was identified as an associated plasma marker, oxLDL and HDL as relevant lipoproteins; macrophage function-related antioxidant Catalase enzyme, lysosome-associated Cathepsin D, S100-A10, and Transforming growth factor-beta-induced protein ig-h3 were identified and selected as associated to atherogenesis outcome | [36] |
| Low-dose streptozotocin-induced diabetic mouse model (10 animals) | LC MS/MS | Dysregulation of molecules involved in myogenesis, vascularization, hypertension, hypertrophy (associated with thickening of the aortic wall), and a substantial reduction in fatty acid storage. A novel finding is the pronounced downregulation of glycogen synthase kinase-3β (Gsk3β) and the upregulation of molecules linked to the tricarboxylic acid cycle (e.g., aspartate aminotransferase (Got2) and hydroxyacid-oxoacid transhydrogenase (Adhfe1)). In addition, pathways involving primary alcohols and amino acid breakdown are altered, potentially leading to ketone-body production. Conclusions: streptozotocin-induced diabetes mellitus in animals leads to a reduction in fatty acid biosynthesis and an upregulation of an alternative ketone-body formation pathway | [189] |
| Plasma and aortic tissue from apo E deficient and wild-type mice | ICAT followed by nanoLC-MS/MS | Here, 13 proteins in the plasma and 36 in the arterial tissues showed significant difference in abundance. These proteins were found to be components of inflammation, thrombus formation, and vascular remodeling, suggesting drastic and integrative alterations in accordance with atherosclerosis development | [31] |
| Cholesterol-fed rabbits | LC MS/MS iTRAQ | Here, 453 unique proteins were identified and quantified; 67 proteins differentially expressed, 62 higher (e.g., apolipoproteins, extracellular matrix adhesion proteins, glycolytic enzymes, heat shock proteins and proteins involved in immune defense) and 5 lower in ascending aortas from HCD-fed rabbits compared to controls | [34] |
| Secretomes of coronary arteries from pigs fed on standard or high-cholesterol diet | LC MS/MS | A wide coronary artery map of secreted proteins has been obtained in high-fat (HF) diet-induced ATS swine model and a significantly different expression of many proteins related to VSMC activation/migration has been identified | [37] |
| Aortas from apo E knock-out mice | LC MS/MS | Here, 5117 proteins were identified, 182 of which changed their expression status in response to vessel maturation and atherosclerotic plaque development. In the insoluble ECM proteome, 65 proteins significantly changed, including relevant collagens, matrix metalloproteinases and macrophage derived proteins | [32] |
| Aortas from SR-uPA+/0 (n = 6) vs. SR-uPA0/0 (n = 6) apo E-/- mice; ruptured (n = 6) and stable (n = 6) areas of human carotid plaques | LC MS/MS | Here, 775 unique proteins were identified; overexpression of urokinase-type plasminogen activator may cause plaque rupture by activating proteolytic and proinflammatory mechanisms leading to ECM and basement-membrane protein depletion, as well as by upregulating the intracellular pathways related to cell–cell adhesion and cell–matrix adhesion. Several biochemical features in common between ruptured human carotid plaques and SR-uPA+/0 mouse aortas | [188] |
4. Conclusions
Author Contributions
Funding
 —NextGenerationEU) for their financial support.
—NextGenerationEU) for their financial support.Data Availability Statement
Conflicts of Interest
References
- Mendis, S.; Nordet, P.; Fernandez-Britto, J.E.; Sternby, N. Atherosclerosis in children and young adults: An overview of the world health organization and international society and federation of cardiology study on pathobiological determinants of atherosclerosis in youth study (1985–1995). Prev. Control 2005, 1, 3–15. [Google Scholar] [CrossRef]
- Hansson, G.K.; Libby, P.; Tabas, I. Inflammation and plaque vulnerability. J. Intern. Med. 2015, 278, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Vilahur, G. Thrombosis formation on atherosclerotic lesions and plaque rupture. J. Intern. Med. 2014, 276, 618–632. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Buring, J.E.; Badimon, L.; Hansson, G.K.; Deanfield, J.; Bittencourt, M.S.; Tokgozoglu, L.; Lewis, E.F. Atherosclerosis. Nat. Rev. Dis. Primers 2019, 5, 56. [Google Scholar] [CrossRef]
- Formato, M.; Farina, M.; Spirito, R.; Maggioni, M.; Guarino, A.; Cherchi, G.M.; Biglioli, P.; Edelstein, C.; Scanu, A.M. Evidence for a proinflammatory and proteolytic environment in plaques from endarterectomy segments of human carotid arteries. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 129–135. [Google Scholar] [CrossRef]
- Lepedda, A.J.; Cigliano, A.; Cherchi, G.M.; Spirito, R.; Maggioni, M.; Carta, F.; Turrini, F.; Edelstein, C.; Scanu, A.M.; Formato, M. A proteomic approach to differentiate histologically classified stable and unstable plaques from human carotid arteries. Atherosclerosis 2009, 203, 112–118. [Google Scholar] [CrossRef]
- Lepedda, A.J.; Zinellu, A.; Nieddu, G.; Zinellu, E.; Carru, C.; Spirito, R.; Guarino, A.; De Muro, P.; Formato, M. Protein sulfhydryl group oxidation and mixed-disulfide modifications in stable and unstable human carotid plaques. Oxidative Med. Cell. Longev. 2013, 2013, 403973. [Google Scholar] [CrossRef]
- Lepedda, A.J.; Formato, M. Oxidative modifications in advanced atherosclerotic plaques: A focus on in situ protein sulfhydryl group oxidation. Oxidative Med. Cell. Longev. 2020, 2020, 6169825. [Google Scholar] [CrossRef]
- Puig, N.; Jimenez-Xarrie, E.; Camps-Renom, P.; Benitez, S. Search for reliable circulating biomarkers to predict carotid plaque vulnerability. Int. J. Mol. Sci. 2020, 21, 8236. [Google Scholar] [CrossRef]
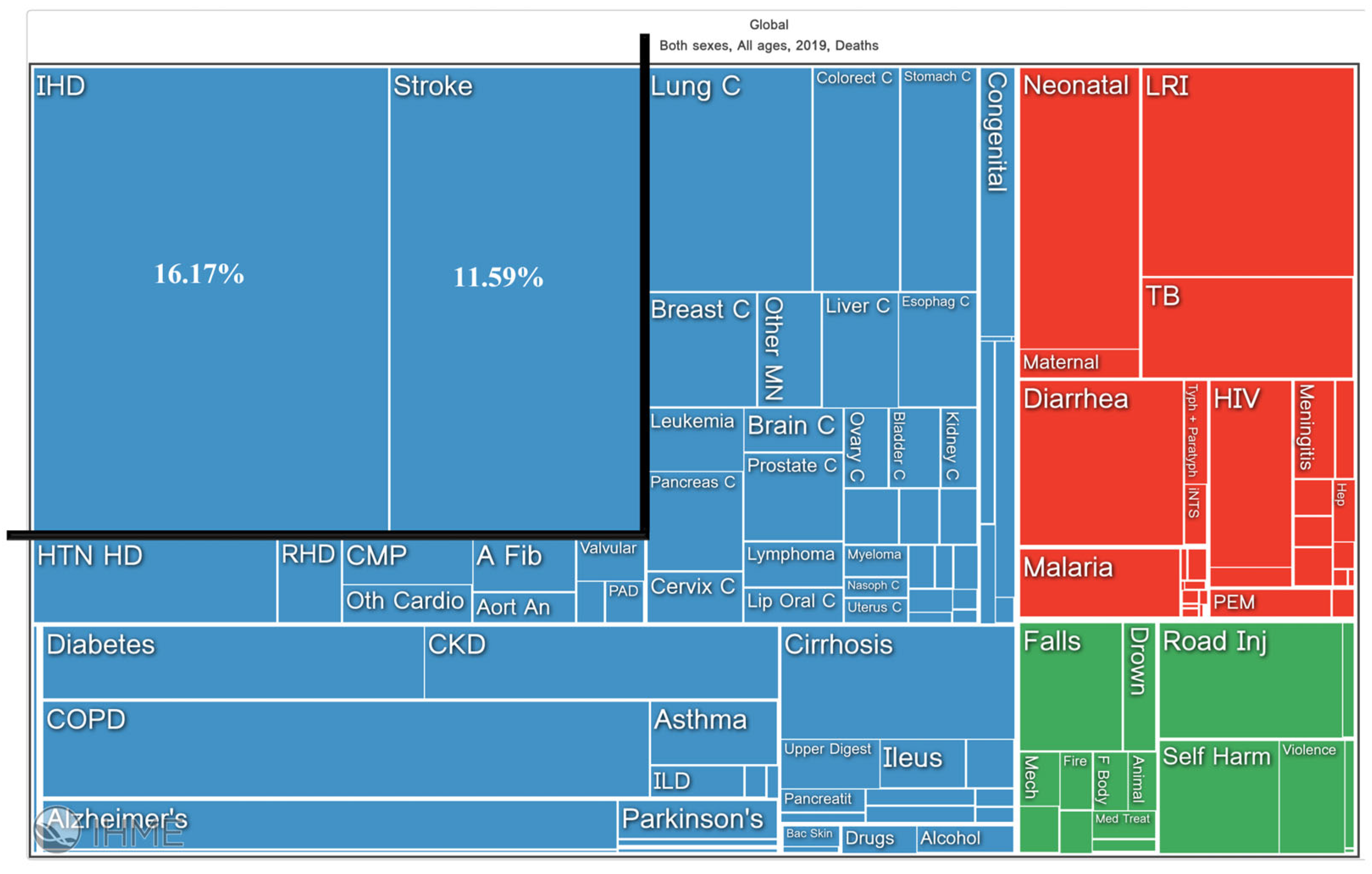
- Available online: https://vizhub.healthdata.org/gbd-compare/InstituteforHealthMetricsandEvaluation (accessed on 14 July 2023).
- Herrington, D.M.; Mao, C.; Parker, S.J.; Fu, Z.; Yu, G.; Chen, L.; Venkatraman, V.; Fu, Y.; Wang, Y.; Howard, T.D.; et al. Proteomic architecture of human coronary and aortic atherosclerosis. Circulation 2018, 137, 2741–2756. [Google Scholar] [CrossRef]
- Tunon, J.; Martin-Ventura, J.L.; Blanco-Colio, L.M.; Lorenzo, O.; Lopez, J.A.; Egido, J. Proteomic strategies in the search of new biomarkers in atherothrombosis. J. Am. Coll. Cardiol. 2010, 55, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- Martinet, W.; Schrijvers, D.M.; De Meyer, G.R.; Herman, A.G.; Kockx, M.M. Western array analysis of human atherosclerotic plaques: Downregulation of apoptosis-linked gene 2. Cardiovasc. Res. 2003, 60, 259–267. [Google Scholar] [CrossRef]
- Slevin, M.; Elasbali, A.B.; Miguel Turu, M.; Krupinski, J.; Badimon, L.; Gaffney, J. Identification of differential protein expression associated with development of unstable human carotid plaques. Am. J. Pathol. 2006, 168, 1004–1021. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; Zullo, A.; Picascia, A.; Infante, T.; Mancini, F.P. Recent advances in proteomic technologies applied to cardiovascular disease. J. Cell. Biochem. 2013, 114, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.Y.; Rui, Y.C.; Yang, P.Y.; Yu, Y.L. Proteomic analysis of foam cells. Methods Mol. Biol. 2007, 357, 297–305. [Google Scholar] [PubMed]
- Yu, J.; Fu, J.; Zhang, X.; Cui, X.; Cheng, M. The integration of metabolomic and proteomic analyses revealed alterations in inflammatory-related protein metabolites in endothelial progenitor cells subjected to oscillatory shear stress. Front. Physiol. 2022, 13, 825966. [Google Scholar] [CrossRef] [PubMed]
- Eligini, S.; Gianazza, E.; Mallia, A.; Ghilardi, S.; Banfi, C. Macrophage phenotyping in atherosclerosis by proteomics. Int. J. Mol. Sci. 2023, 24, 2613. [Google Scholar] [CrossRef]
- Eslava-Alcon, S.; Extremera-Garcia, M.J.; Gonzalez-Rovira, A.; Rosal-Vela, A.; Rojas-Torres, M.; Beltran-Camacho, L.; Sanchez-Gomar, I.; Jimenez-Palomares, M.; Alonso-Pinero, J.A.; Conejero, R.; et al. Molecular signatures of atherosclerotic plaques: An up-dated panel of protein related markers. J. Proteom. 2020, 221, 103757. [Google Scholar] [CrossRef]
- De la Cuesta, F.; Alvarez-Llamas, G.; Maroto, A.S.; Barderas, M.G.; Vivanco, F. Laser microdissection and saturation labeling dige method for the analysis of human arteries. Methods Mol. Biol. 2013, 1000, 21–32. [Google Scholar]
- Rocchiccioli, S.; Pelosi, G.; Rosini, S.; Marconi, M.; Viglione, F.; Citti, L.; Ferrari, M.; Trivella, M.G.; Cecchettini, A. Secreted proteins from carotid endarterectomy: An untargeted approach to disclose molecular clues of plaque progression. J. Transl. Med. 2013, 11, 260. [Google Scholar] [CrossRef]
- Lygirou, V.; Latosinska, A.; Makridakis, M.; Mullen, W.; Delles, C.; Schanstra, J.P.; Zoidakis, J.; Pieske, B.; Mischak, H.; Vlahou, A. Plasma proteomic analysis reveals altered protein abundances in cardiovascular disease. J. Transl. Med. 2018, 16, 104. [Google Scholar] [CrossRef] [PubMed]
- Delles, C.; Diez, J.; Dominiczak, A.F. Urinary proteomics in cardiovascular disease: Achievements, limits and hopes. Proteom. Clin. Appl. 2011, 5, 222–232. [Google Scholar] [CrossRef]
- Hoofnagle, A.N.; Heinecke, J.W. Lipoproteomics: Using mass spectrometry-based proteomics to explore the assembly, structure, and function of lipoproteins. J. Lipid Res. 2009, 50, 1967–1975. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.S.; Tan, L.; Long, J.L.; Davidson, W.S. Proteomic diversity of high density lipoproteins: Our emerging understanding of its importance in lipid transport and beyond. J. Lipid Res. 2013, 54, 2575–2585. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.D.; Burger, P.C. Platelets in inflammation and thrombosis. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 2131–2137. [Google Scholar] [CrossRef]
- Thomas, M.R.; Storey, R.F. The role of platelets in inflammation. Thromb. Haemost. 2015, 114, 449–458. [Google Scholar]
- Gutmann, C.; Joshi, A.; Mayr, M. Platelet “-omics” in health and cardiovascular disease. Atherosclerosis 2020, 307, 87–96. [Google Scholar] [CrossRef]
- Mayr, M.; Chung, Y.L.; Mayr, U.; Yin, X.; Ly, L.; Troy, H.; Fredericks, S.; Hu, Y.; Griffiths, J.R.; Xu, Q. Proteomic and metabolomic analyses of atherosclerotic vessels from apolipoprotein e-deficient mice reveal alterations in inflammation, oxidative stress, and energy metabolism. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2135–2142. [Google Scholar] [CrossRef]
- Wu, J.; Liu, W.; Sousa, E.; Qiu, Y.; Pittman, D.D.; Maganti, V.; Feldman, J.; Gill, D.; Lu, Z.; Dorner, A.J.; et al. Proteomic identification of endothelial proteins isolated in situ from atherosclerotic aorta via systemic perfusion. J. Proteome Res. 2007, 6, 4728–4736. [Google Scholar] [CrossRef]
- Hanzawa, H.; Sakamoto, T.; Kaneko, A.; Manri, N.; Zhao, Y.; Zhao, S.; Tamaki, N.; Kuge, Y. Combined plasma and tissue proteomic study of atherogenic model mouse: Approach to elucidate molecular determinants in atherosclerosis development. J. Proteome Res. 2015, 14, 4257–4269. [Google Scholar] [CrossRef]
- Wierer, M.; Prestel, M.; Schiller, H.B.; Yan, G.; Schaab, C.; Azghandi, S.; Werner, J.; Kessler, T.; Malik, R.; Murgia, M.; et al. Compartment-resolved proteomic analysis of mouse aorta during atherosclerotic plaque formation reveals osteoclast-specific protein expression. Mol. Cell. Proteom. 2018, 17, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Kaga, E.; Karademir, B.; Baykal, A.T.; Ozer, N.K. Identification of differentially expressed proteins in atherosclerotic aorta and effect of vitamin e. J. Proteom. 2013, 92, 260–273. [Google Scholar] [CrossRef]
- Xu, J.; Jullig, M.; Middleditch, M.J.; Cooper, G.J. Modelling atherosclerosis by proteomics: Molecular changes in the ascending aortas of cholesterol-fed rabbits. Atherosclerosis 2015, 242, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Rather, R.A.; Malik, V.S.; Trikha, D.; Bhat, O.; Dhawan, V. Aqueous terminalia arjuna extract modulates expression of key atherosclerosis-related proteins in a hypercholesterolemic rabbit: A proteomic-based study. Proteomics. Clin. Appl. 2016, 10, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, G.; Rocchiccioli, S.; Cecchettini, A.; Viglione, F.; Puntoni, M.; Parodi, O.; Capobianco, E.; Trivella, M.G. Inflammation blood and tissue factors of plaque growth in an experimental model evidenced by a systems approach. Front. Genet. 2014, 5, 70. [Google Scholar] [CrossRef] [PubMed]
- Rocchiccioli, S.; Cecchettini, A.; Ucciferri, N.; Terreni, M.; Viglione, F.; Trivella, M.G.; Citti, L.; Parodi, O.; Pelosi, G. Site-specific secretome map evidences vsmc-related markers of coronary atherosclerosis grade and extent in the hypercholesterolemic swine. Dis. Markers 2015, 2015, 465242. [Google Scholar] [CrossRef]
- Ekroos, K.; Janis, M.; Tarasov, K.; Hurme, R.; Laaksonen, R. Lipidomics: A tool for studies of atherosclerosis. Curr. Atheroscler. Rep. 2010, 12, 273–281. [Google Scholar] [CrossRef]
- Stegemann, C.; Pechlaner, R.; Willeit, P.; Langley, S.R.; Mangino, M.; Mayr, U.; Menni, C.; Moayyeri, A.; Santer, P.; Rungger, G.; et al. Lipidomics profiling and risk of cardiovascular disease in the prospective population-based bruneck study. Circulation 2014, 129, 1821–1831. [Google Scholar] [CrossRef]
- Wang, D.; Yu, B.; Li, Q.; Guo, Y.; Koike, T.; Koike, Y.; Wu, Q.; Zhang, J.; Mao, L.; Tang, X.; et al. HDL quality features revealed by proteome–lipidome connectivity are associated with atherosclerotic disease. J. Mol. Cell Biol. 2022, 14, mjac004. [Google Scholar] [CrossRef]
- Stegemann, C.; Drozdov, I.; Shalhoub, J.; Humphries, J.; Ladroue, C.; Didangelos, A.; Baumert, M.; Allen, M.; Davies, A.H.; Monaco, C.; et al. Comparative lipidomics profiling of human atherosclerotic plaques. Circ. Cardiovasc. Genet. 2011, 4, 232–242. [Google Scholar] [CrossRef]
- Moerman, A.M.; Visscher, M.; Slijkhuis, N.; Van Gaalen, K.; Heijs, B.; Klein, T.; Burgers, P.C.; De Rijke, Y.B.; Van Beusekom, H.M.M.; Luider, T.M.; et al. Lipid signature of advanced human carotid atherosclerosis assessed by mass spectrometry imaging. J. Lipid Res. 2021, 62, 100020. [Google Scholar] [CrossRef] [PubMed]
- Lepedda, A.J.; Zinellu, E.; Formato, M. Overview of Current Proteomic Approaches for Discovery of Vascular Biomarkers of Atherosclerosis. In Proteomics; Tsz-Kwong, M., Ricardo, J.F., Eds.; IntechOpen: Rijeka, Croatia, 2012; Chapter 1. [Google Scholar]
- Jonas, A.; Phillips, M.C. Chapter 17—Lipoprotein structure. In Biochemistry of Lipids, Lipoproteins and Membranes, 5th ed.; Vance, D.E., Vance, J.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 485–506. [Google Scholar]
- Vance, J.E.; Vance, D.E. Biochemistry of Lipids, Lipoproteins and Membranes; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Scanu, A.M. Lipoprotein(a) as a cardiovascular risk factor. Trends Cardiovasc. Med. 1991, 1, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Ferhatbegovic, L.; Mrsic, D.; Kusljugic, S.; Pojskic, B. LDL-c: The only causal risk factor for ascvd. Why is it still overlooked and underestimated? Curr. Atheroscler. Rep. 2022, 24, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.J.; Tabas, I. The response-to-retention hypothesis of early atherogenesis. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 551–561. [Google Scholar] [CrossRef]
- Ouimet, M.; Barrett, T.J.; Fisher, E.A. HDL and reverse cholesterol transport. Circ. Res. 2019, 124, 1505–1518. [Google Scholar] [CrossRef]
- Kajani, S.; Curley, S.; McGillicuddy, F.C. Unravelling HDL-looking beyond the cholesterol surface to the quality within. Int. J. Mol. Sci. 2018, 19, 1971. [Google Scholar] [CrossRef]
- Vaisar, T.; Pennathur, S.; Green, P.S.; Gharib, S.A.; Hoofnagle, A.N.; Cheung, M.C.; Byun, J.; Vuletic, S.; Kassim, S.; Singh, P.; et al. Shotgun proteomics implicates protease inhibition and complement activation in the antiinflammatory properties of HDL. J. Clin. Investig. 2007, 117, 746–756. [Google Scholar] [CrossRef]
- Holzer, M.; Ljubojevic-Holzer, S.; Souza Junior, D.R.; Stadler, J.T.; Rani, A.; Scharnagl, H.; Ronsein, G.E.; Marsche, G. HDL isolated by immunoaffinity, ultracentrifugation, or precipitation is compositionally and functionally distinct. J. Lipid Res. 2022, 63, 100307. [Google Scholar] [CrossRef]
- Havel, R.J.; Eder, H.A.; Bragdon, J.H. The distribution and chemical composition of ultracentrifugally separated lipoproteins in human serum. J. Clin. Investig. 1955, 34, 1345–1353. [Google Scholar] [CrossRef]
- Chary, A.; Hedayati, M. Review of laboratory methods to determine HDL and LDL subclasses and their clinical importance. Rev. Cardiovasc. Med. 2022, 23, 147. [Google Scholar] [CrossRef]
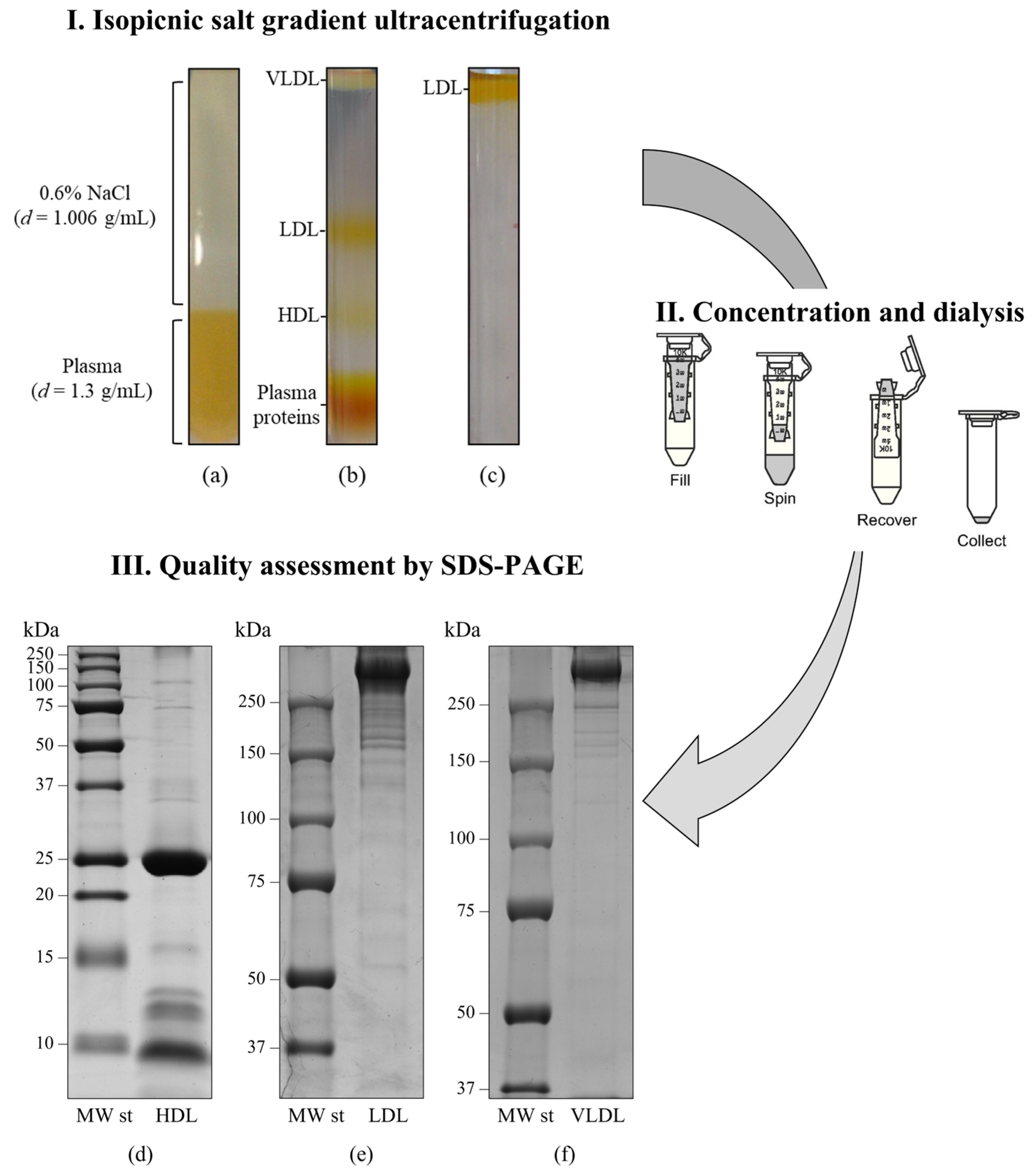
- Nieddu, G.; Michelucci, E.; Formato, M.; Ciampelli, C.; Obino, G.; Signore, G.; Di Giorgi, N.; Rocchiccioli, S.; Lepedda, A.J. Molecular characterization of plasma HDL, LDL, and VLDL lipids cargos from atherosclerotic patients with advanced carotid lesions: A preliminary report. Int. J. Mol. Sci. 2022, 23, 12449. [Google Scholar] [CrossRef] [PubMed]
- Finamore, F.; Nieddu, G.; Rocchiccioli, S.; Spirito, R.; Guarino, A.; Formato, M.; Lepedda, A.J. Apolipoprotein signature of HDL and LDL from atherosclerotic patients in relation with carotid plaque typology: A preliminary report. Biomedicines 2021, 9, 1156. [Google Scholar] [CrossRef] [PubMed]
- Mancone, C.; Amicone, L.; Fimia, G.M.; Bravo, E.; Piacentini, M.; Tripodi, M.; Alonzi, T. Proteomic analysis of human very low-density lipoprotein by two-dimensional gel electrophoresis and maldi-tof/tof. Proteomics 2007, 7, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Banfi, C.; Brioschi, M.; Barcella, S.; Wait, R.; Begum, S.; Galli, S.; Rizzi, A.; Tremoli, E. Proteomic analysis of human low-density lipoprotein reveals the presence of prenylcysteine lyase, a hydrogen peroxide-generating enzyme. Proteomics 2009, 9, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Davidson, W.S.; Silva, R.A.; Chantepie, S.; Lagor, W.R.; Chapman, M.J.; Kontush, A. Proteomic analysis of defined HDL subpopulations reveals particle-specific protein clusters: Relevance to antioxidative function. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 870–876. [Google Scholar] [CrossRef]
- Bondarenko, P.V.; Cockrill, S.L.; Watkins, L.K.; Cruzado, I.D.; Macfarlane, R.D. Mass spectral study of polymorphism of the apolipoproteins of very low density lipoprotein. J. Lipid Res. 1999, 40, 543–555. [Google Scholar] [CrossRef]
- Sun, H.Y.; Chen, S.F.; Lai, M.D.; Chang, T.T.; Chen, T.L.; Li, P.Y.; Shieh, D.B.; Young, K.C. Comparative proteomic profiling of plasma very-low-density and low-density lipoproteins. Clin. Chim. Acta Int. J. Clin. Chem. 2010, 411, 336–344. [Google Scholar] [CrossRef]
- Stahlman, M.; Davidsson, P.; Kanmert, I.; Rosengren, B.; Boren, J.; Fagerberg, B.; Camejo, G. Proteomics and lipids of lipoproteins isolated at low salt concentrations in d2o/sucrose or in kbr. J. Lipid Res. 2008, 49, 481–490. [Google Scholar] [CrossRef]
- Bottcher, A.; Schlosser, J.; Kronenberg, F.; Dieplinger, H.; Knipping, G.; Lackner, K.J.; Schmitz, G. Preparative free-solution isotachophoresis for separation of human plasma lipoproteins: Apolipoprotein and lipid composition of HDL subfractions. J. Lipid Res. 2000, 41, 905–915. [Google Scholar] [CrossRef]
- Collins, L.A.; Olivier, M. Quantitative comparison of lipoprotein fractions derived from human plasma and serum by liquid chromatography-tandem mass spectrometry. Proteome Sci. 2010, 8, 42. [Google Scholar] [CrossRef]
- Gordon, S.M.; Deng, J.; Lu, L.J.; Davidson, W.S. Proteomic characterization of human plasma high density lipoprotein fractionated by gel filtration chromatography. J. Proteome Res. 2010, 9, 5239–5249. [Google Scholar] [CrossRef]
- Javeed, R.; Hussain, D.; Jabeen, F.; Saeed, A.; Fatima, B.; Ashiq, M.N.; Najam-Ul-Haq, M. Enrichment of HDL proteome and phospholipidome from human serum via imac/moac affinity. Biomed. Chromatogr. 2020, 34, e4693. [Google Scholar] [CrossRef]
- Davidson, W.S.; Shah, A.S.; Sexmith, H.; Gordon, S.M. The HDL proteome watch: Compilation of studies leads to new insights on HDL function. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2022, 1867, 159072. [Google Scholar] [CrossRef]
- Dashty, M.; Motazacker, M.M.; Levels, J.; de Vries, M.; Mahmoudi, M.; Peppelenbosch, M.P.; Rezaee, F. Proteome of human plasma very low-density lipoprotein and low-density lipoprotein exhibits a link with coagulation and lipid metabolism. Thromb. Haemost. 2014, 111, 518–530. [Google Scholar] [CrossRef] [PubMed]
- Bancells, C.; Canals, F.; Benitez, S.; Colome, N.; Julve, J.; Ordonez-Llanos, J.; Sanchez-Quesada, J.L. Proteomic analysis of electronegative low-density lipoprotein. J. Lipid Res. 2010, 51, 3508–3515. [Google Scholar] [CrossRef]
- Heller, M.; Stalder, D.; Schlappritzi, E.; Hayn, G.; Matter, U.; Haeberli, A. Mass spectrometry-based analytical tools for the molecular protein characterization of human plasma lipoproteins. Proteomics 2005, 5, 2619–2630. [Google Scholar] [CrossRef] [PubMed]
- Collins, L.A.; Mirza, S.P.; Kissebah, A.H.; Olivier, M. Integrated approach for the comprehensive characterization of lipoproteins from human plasma using fplc and nano-hplc-tandem mass spectrometry. Physiol. Genom. 2010, 40, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, K.C.; Tio, R.A.; Zeebregts, C.J.; Bijlsma, M.F.; Zijlstra, F.; Badlou, B.; de Vries, M.; Ferreira, C.V.; Spek, C.A.; Peppelenbosch, M.P.; et al. Human plasma very low density lipoprotein carries indian hedgehog. J. Proteome Res. 2010, 9, 6052–6059. [Google Scholar] [CrossRef] [PubMed]
- Von Zychlinski, A.; Williams, M.; McCormick, S.; Kleffmann, T. Absolute quantification of apolipoproteins and associated proteins on human plasma lipoproteins. J. Proteom. 2014, 106, 181–190. [Google Scholar] [CrossRef]
- Von Zychlinski, A.; Kleffmann, T.; Williams, M.J.; McCormick, S.P. Proteomics of lipoprotein(a) identifies a protein complement associated with response to wounding. J. Proteom. 2011, 74, 2881–2891. [Google Scholar] [CrossRef]
- Bourgeois, R.; Girard, A.; Perrot, N.; Guertin, J.; Mitchell, P.L.; Couture, C.; Gotti, C.; Bourassa, S.; Poggio, P.; Mass, E.; et al. A comparative analysis of the lipoprotein(a) and low-density lipoprotein proteomic profiles combining mass spectrometry and mendelian randomization. CJC Open 2021, 3, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Mueller, P.A.; Yerkes, E.; Bergstrom, P.; Rosario, S.; Hay, J.; Pamir, N. A method for lipoprotein (a) isolation from a small volume of plasma with applications for clinical research. Sci. Rep. 2022, 12, 9138. [Google Scholar] [CrossRef] [PubMed]
- Green, P.S.; Vaisar, T.; Pennathur, S.; Kulstad, J.J.; Moore, A.B.; Marcovina, S.; Brunzell, J.; Knopp, R.H.; Zhao, X.Q.; Heinecke, J.W. Combined statin and niacin therapy remodels the high-density lipoprotein proteome. Circulation 2008, 118, 1259–1267. [Google Scholar] [CrossRef]
- Heller, M.; Schlappritzi, E.; Stalder, D.; Nuoffer, J.M.; Haeberli, A. Compositional protein analysis of high density lipoproteins in hypercholesterolemia by shotgun lc-ms/ms and probabilistic peptide scoring. Mol. Cell. Proteom. 2007, 6, 1059–1072. [Google Scholar] [CrossRef]
- Vaisar, T.; Mayer, P.; Nilsson, E.; Zhao, X.Q.; Knopp, R.; Prazen, B.J. HDL in humans with cardiovascular disease exhibits a proteomic signature. Clin. Chim. Acta Int. J. Clin. Chem. 2010, 411, 972–979. [Google Scholar] [CrossRef]
- Levels, J.H.; Geurts, P.; Karlsson, H.; Maree, R.; Ljunggren, S.; Fornander, L.; Wehenkel, L.; Lindahl, M.; Stroes, E.S.; Kuivenhoven, J.A.; et al. High-density lipoprotein proteome dynamics in human endotoxemia. Proteome Sci. 2011, 9, 34. [Google Scholar] [CrossRef]
- Cubedo, J.; Padro, T.; Garcia-Moll, X.; Pinto, X.; Cinca, J.; Badimon, L. Proteomic signature of apolipoprotein j in the early phase of new-onset myocardial infarction. J. Proteome Res. 2011, 10, 211–220. [Google Scholar] [CrossRef]
- Alwaili, K.; Bailey, D.; Awan, Z.; Bailey, S.D.; Ruel, I.; Hafiane, A.; Krimbou, L.; Laboissiere, S.; Genest, J. The HDL proteome in acute coronary syndromes shifts to an inflammatory profile. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2012, 1821, 405–415. [Google Scholar] [CrossRef]
- Riwanto, M.; Rohrer, L.; Roschitzki, B.; Besler, C.; Mocharla, P.; Mueller, M.; Perisa, D.; Heinrich, K.; Altwegg, L.; von Eckardstein, A.; et al. Altered activation of endothelial anti- and proapoptotic pathways by high-density lipoprotein from patients with coronary artery disease: Role of high-density lipoprotein-proteome remodeling. Circulation 2013, 127, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Lepedda, A.J.; Nieddu, G.; Zinellu, E.; De Muro, P.; Piredda, F.; Guarino, A.; Spirito, R.; Carta, F.; Turrini, F.; Formato, M. Proteomic analysis of plasma-purified VLDL, LDL, and HDL fractions from atherosclerotic patients undergoing carotid endarterectomy: Identification of serum amyloid a as a potential marker. Oxidative Med. Cell. Longev. 2013, 2013, 385214. [Google Scholar] [CrossRef]
- Mazur, M.T.; Cardasis, H.L. Quantitative analysis of apolipoproteins in human HDL by top-down differential mass spectrometry. Methods Mol. Biol. 2013, 1000, 115–137. [Google Scholar] [PubMed]
- Yan, L.R.; Wang, D.X.; Liu, H.; Zhang, X.X.; Zhao, H.; Hua, L.; Xu, P.; Li, Y.S. A pro-atherogenic HDL profile in coronary heart disease patients: An itraq labelling-based proteomic approach. PLoS ONE 2014, 9, e98368. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Liu, T.R.; Hu, S.W.; Tian, D.; Li, C.; Zhong, J.K.; Sun, H.G.; Luo, T.T.; Lai, W.Y.; Guo, Z.G. Acute coronary syndrome remodels the protein cargo and functions of high-density lipoprotein subfractions. PLoS ONE 2014, 9, e94264. [Google Scholar] [CrossRef] [PubMed]
- Yassine, H.N.; Jackson, A.M.; Borges, C.R.; Billheimer, D.; Koh, H.; Smith, D.; Reaven, P.; Lau, S.S.; Borchers, C.H. The application of multiple reaction monitoring and multi-analyte profiling to HDL proteins. Lipids Health Dis. 2014, 13, 8. [Google Scholar] [CrossRef]
- Jorge, I.; Burillo, E.; Mesa, R.; Baila-Rueda, L.; Moreno, M.; Trevisan-Herraz, M.; Silla-Castro, J.C.; Camafeita, E.; Ortega-Munoz, M.; Bonzon-Kulichenko, E.; et al. The human HDL proteome displays high inter-individual variability and is altered dynamically in response to angioplasty-induced atheroma plaque rupture. J. Proteom. 2014, 106, 61–73. [Google Scholar] [CrossRef]
- Krishnan, S.; Huang, J.; Lee, H.; Guerrero, A.; Berglund, L.; Anuurad, E.; Lebrilla, C.B.; Zivkovic, A.M. Combined high-density lipoprotein proteomic and glycomic profiles in patients at risk for coronary artery disease. J. Proteome Res. 2015, 14, 5109–5118. [Google Scholar] [CrossRef]
- Pedret, A.; Catalan, U.; Fernandez-Castillejo, S.; Farras, M.; Valls, R.M.; Rubio, L.; Canela, N.; Aragones, G.; Romeu, M.; Castaner, O.; et al. Impact of virgin olive oil and phenol-enriched virgin olive oils on the HDL proteome in hypercholesterolemic subjects: A double blind, randomized, controlled, cross-over clinical trial (VOHF study). PLoS ONE 2015, 10, e0129160. [Google Scholar] [CrossRef]
- Marsillach, J.; Becker, J.O.; Vaisar, T.; Hahn, B.H.; Brunzell, J.D.; Furlong, C.E.; de Boer, I.H.; McMahon, M.A.; Hoofnagle, A.N.; Group, D.E.R. Paraoxonase-3 is depleted from the high-density lipoproteins of autoimmune disease patients with subclinical atherosclerosis. J. Proteome Res. 2015, 14, 2046–2054. [Google Scholar] [CrossRef]
- Gordon, S.M.; Li, H.; Zhu, X.; Tso, P.; Reardon, C.A.; Shah, A.S.; Lu, L.J.; Davidson, W.S. Impact of genetic deletion of platform apolipoproteins on the size distribution of the murine lipoproteome. J. Proteom. 2016, 146, 184–194. [Google Scholar] [CrossRef]
- Oberbach, A.; Adams, V.; Schlichting, N.; Heinrich, M.; Kullnick, Y.; Lehmann, S.; Lehmann, S.; Feder, S.; Correia, J.C.; Mohr, F.W.; et al. Proteome profiles of HDL particles of patients with chronic heart failure are associated with immune response and also include bacteria proteins. Clin. Chim. Acta Int. J. Clin. Chem. 2016, 453, 114–122. [Google Scholar] [CrossRef]
- Burillo, E.; Jorge, I.; Martinez-Lopez, D.; Camafeita, E.; Blanco-Colio, L.M.; Trevisan-Herraz, M.; Ezkurdia, I.; Egido, J.; Michel, J.B.; Meilhac, O.; et al. Quantitative HDL proteomics identifies peroxiredoxin-6 as a biomarker of human abdominal aortic aneurysm. Sci. Rep. 2016, 6, 38477. [Google Scholar] [CrossRef] [PubMed]
- Ljunggren, S.A.; Helmfrid, I.; Norinder, U.; Fredriksson, M.; Wingren, G.; Karlsson, H.; Lindahl, M. Alterations in high-density lipoprotein proteome and function associated with persistent organic pollutants. Environ. Int. 2017, 98, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.M.; Chung, J.H.; Playford, M.P.; Dey, A.K.; Sviridov, D.; Seifuddin, F.; Chen, Y.C.; Pirooznia, M.; Chen, M.Y.; Mehta, N.N.; et al. High density lipoprotein proteome is associated with cardiovascular risk factors and atherosclerosis burden as evaluated by coronary ct angiography. Atherosclerosis 2018, 278, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Emmens, J.E.; Jones, D.J.L.; Cao, T.H.; Chan, D.C.S.; Romaine, S.P.R.; Quinn, P.A.; Anker, S.D.; Cleland, J.G.; Dickstein, K.; Filippatos, G.; et al. Proteomic diversity of high-density lipoprotein explains its association with clinical outcome in patients with heart failure. Eur. J. Heart Fail. 2018, 20, 260–267. [Google Scholar] [CrossRef]
- Natarajan, P.; Collier, T.S.; Jin, Z.; Lyass, A.; Li, Y.; Ibrahim, N.E.; Mukai, R.; McCarthy, C.P.; Massaro, J.M.; D’Agostino, R.B., Sr.; et al. Association of an HDL apolipoproteomic score with coronary atherosclerosis and cardiovascular death. J. Am. Coll. Cardiol. 2019, 73, 2135–2145. [Google Scholar] [CrossRef]
- Jin, Z.; Collier, T.S.; Dai, D.L.Y.; Chen, V.; Hollander, Z.; Ng, R.T.; McManus, B.M.; Balshaw, R.; Apostolidou, S.; Penn, M.S.; et al. Development and validation of apolipoprotein ai-associated lipoprotein proteome panel for the prediction of cholesterol efflux capacity and coronary artery disease. Clin. Chem. 2019, 65, 282–290. [Google Scholar] [CrossRef]
- Okada, T.; Ohama, T.; Takafuji, K.; Kanno, K.; Matsuda, H.; Sairyo, M.; Zhu, Y.; Saga, A.; Kobayashi, T.; Masuda, D.; et al. Shotgun proteomic analysis reveals proteome alterations in HDL of patients with cholesteryl ester transfer protein deficiency. J. Clin. Lipidol. 2019, 13, 317–325. [Google Scholar] [CrossRef]
- Plubell, D.L.; Fenton, A.M.; Rosario, S.; Bergstrom, P.; Wilmarth, P.A.; Clark, W.M.; Zakai, N.A.; Quinn, J.F.; Minnier, J.; Alkayed, N.J.; et al. High-density lipoprotein carries markers that track with recovery from stroke. Circ. Res. 2020, 127, 1274–1287. [Google Scholar] [CrossRef]
- Shao, B.; Mathew, A.V.; Thornock, C.; Pennathur, S.; Michigan Kidney Translational Core, C.I.G. Altered HDL proteome predicts incident cvd in chronic kidney disease patients. J. Lipid Res. 2021, 62, 100135. [Google Scholar] [CrossRef]
- Holzer, M.; Birner-Gruenberger, R.; Stojakovic, T.; El-Gamal, D.; Binder, V.; Wadsack, C.; Heinemann, A.; Marsche, G. Uremia alters HDL composition and function. J. Am. Soc. Nephrol. JASN 2011, 22, 1631–1641. [Google Scholar] [CrossRef]
- Mange, A.; Goux, A.; Badiou, S.; Patrier, L.; Canaud, B.; Maudelonde, T.; Cristol, J.P.; Solassol, J. HDL proteome in hemodialysis patients: A quantitative nanoflow liquid chromatography-tandem mass spectrometry approach. PLoS ONE 2012, 7, e34107. [Google Scholar] [CrossRef] [PubMed]
- Tolle, M.; Huang, T.; Schuchardt, M.; Jankowski, V.; Prufer, N.; Jankowski, J.; Tietge, U.J.; Zidek, W.; van der Giet, M. High-density lipoprotein loses its anti-inflammatory capacity by accumulation of pro-inflammatory-serum amyloid a. Cardiovasc. Res. 2012, 94, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Weichhart, T.; Kopecky, C.; Kubicek, M.; Haidinger, M.; Doller, D.; Katholnig, K.; Suarna, C.; Eller, P.; Tolle, M.; Gerner, C.; et al. Serum amyloid a in uremic HDL promotes inflammation. J. Am. Soc. Nephrol. JASN 2012, 23, 934–947. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.; de Boer, I.; Tang, C.; Mayer, P.S.; Zelnick, L.; Afkarian, M.; Heinecke, J.W.; Himmelfarb, J. A cluster of proteins implicated in kidney disease is increased in high-density lipoprotein isolated from hemodialysis subjects. J. Proteome Res. 2015, 14, 2792–2806. [Google Scholar] [CrossRef]
- Rubinow, K.B.; Henderson, C.M.; Robinson-Cohen, C.; Himmelfarb, J.; de Boer, I.H.; Vaisar, T.; Kestenbaum, B.; Hoofnagle, A.N. Kidney function is associated with an altered protein composition of high-density lipoprotein. Kidney Int. 2017, 92, 1526–1535. [Google Scholar] [CrossRef]
- Wang, K.; Zelnick, L.R.; Hoofnagle, A.N.; Vaisar, T.; Henderson, C.M.; Imrey, P.B.; Robinson-Cohen, C.; de Boer, I.H.; Shiu, Y.T.; Himmelfarb, J.; et al. Alteration of HDL protein composition with hemodialysis initiation. Clin. J. Am. Soc. Nephrol. CJASN 2018, 13, 1225–1233. [Google Scholar] [CrossRef]
- Florens, N.; Calzada, C.; Delolme, F.; Page, A.; Guebre Egziabher, F.; Juillard, L.; Soulage, A.C.O. Proteomic characterization of high-density lipoprotein particles from non-diabetic hemodialysis patients. Toxins 2019, 11, 671. [Google Scholar] [CrossRef]
- Manjunatha, S.; Distelmaier, K.; Dasari, S.; Carter, R.E.; Kudva, Y.C.; Nair, K.S. Functional and proteomic alterations of plasma high density lipoproteins in type 1 diabetes mellitus. Metab. Clin. Exp. 2016, 65, 1421–1431. [Google Scholar] [CrossRef]
- Shao, B.; Zelnick, L.R.; Wimberger, J.; Himmelfarb, J.; Brunzell, J.; Davidson, W.S.; Snell-Bergeon, J.K.; Bornfeldt, K.E.; de Boer, I.H.; Heinecke, J.W. Albuminuria, the high-density lipoprotein proteome, and coronary artery calcification in type 1 diabetes mellitus. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1483–1491. [Google Scholar] [CrossRef]
- Gourgari, E.; Ma, J.; Playford, M.P.; Mehta, N.N.; Goldman, R.; Remaley, A.T.; Gordon, S.M. Proteomic alterations of HDL in youth with type 1 diabetes and their associations with glycemic control: A case-control study. Cardiovasc. Diabetol. 2019, 18, 43. [Google Scholar] [CrossRef]
- Vaisar, T.; Kanter, J.E.; Wimberger, J.; Irwin, A.D.; Gauthier, J.; Wolfson, E.; Bahnam, V.; Wu, I.H.; Shah, H.; Keenan, H.A.; et al. High concentration of medium-sized HDL particles and enrichment in HDL paraoxonase 1 associate with protection from vascular complications in people with long-standing type 1 diabetes. Diabetes Care 2020, 43, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Kheniser, K.G.; Osme, A.; Kim, C.; Ilchenko, S.; Kasumov, T.; Kashyap, S.R. Temporal dynamics of high-density lipoprotein proteome in diet-controlled subjects with type 2 diabetes. Biomolecules 2020, 10, 520. [Google Scholar] [CrossRef] [PubMed]
- Gourgari, E.; Nadeau, K.J.; Pyle, L.; Playford, M.P.; Ma, J.; Mehta, N.N.; Remaley, A.T.; Gordon, S.M. Effect of metformin on the high-density lipoprotein proteome in youth with type 1 diabetes. Endocrinol. Diabetes Metab. 2021, 4, e00261. [Google Scholar] [CrossRef] [PubMed]
- Toyoshima, M.T.K.; Santana, M.F.M.; Silva, A.R.M.; Mello, G.B.; Santos-Bezerra, D.P.; Goes, M.F.S.; Bosco, A.A.; Caramelli, B.; Ronsein, G.E.; Correa-Giannella, M.L.; et al. Proteomics of high-density lipoprotein subfractions and subclinical atherosclerosis in type 1 diabetes mellitus: A case-control study. Diabetol. Metab. Syndr. 2023, 15, 42. [Google Scholar] [CrossRef]
- Khovidhunkit, W.; Duchateau, P.N.; Medzihradszky, K.F.; Moser, A.H.; Naya-Vigne, J.; Shigenaga, J.K.; Kane, J.P.; Grunfeld, C.; Feingold, K.R. Apolipoproteins a-iv and a-v are acute-phase proteins in mouse HDL. Atherosclerosis 2004, 176, 37–44. [Google Scholar] [CrossRef]
- O’Reilly, M.; Dillon, E.; Guo, W.; Finucane, O.; McMorrow, A.; Murphy, A.; Lyons, C.; Jones, D.; Ryan, M.; Gibney, M.; et al. High-density lipoprotein proteomic composition, and not efflux capacity, reflects differential modulation of reverse cholesterol transport by saturated and monounsaturated fat diets. Circulation 2016, 133, 1838–1850. [Google Scholar] [CrossRef]
- Cao, J.; Xu, Y.; Li, F.; Shang, L.; Fan, D.; Yu, H. Protein markers of dysfunctional HDL in scavenger receptor class b type i deficient mice. J. Transl. Med. 2018, 16, 155. [Google Scholar] [CrossRef]
- Camont, L.; Lhomme, M.; Rached, F.; Le Goff, W.; Negre-Salvayre, A.; Salvayre, R.; Calzada, C.; Lagarde, M.; Chapman, M.J.; Kontush, A. Small, dense high-density lipoprotein-3 particles are enriched in negatively charged phospholipids: Relevance to cellular cholesterol efflux, antioxidative, antithrombotic, anti-inflammatory, and antiapoptotic functionalities. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2715–2723. [Google Scholar] [CrossRef]
- Borodzicz, S.; Czarzasta, K.; Kuch, M.; Cudnoch-Jedrzejewska, A. Sphingolipids in cardiovascular diseases and metabolic disorders. Lipids Health Dis. 2015, 14, 55. [Google Scholar] [CrossRef]
- Dang, V.T.; Huang, A.; Zhong, L.H.; Shi, Y.; Werstuck, G.H. Comprehensive plasma metabolomic analyses of atherosclerotic progression reveal alterations in glycerophospholipid and sphingolipid metabolism in apolipoprotein e-deficient mice. Sci. Rep. 2016, 6, 35037. [Google Scholar] [CrossRef]
- Mantovani, A.; Bonapace, S.; Lunardi, G.; Canali, G.; Dugo, C.; Vinco, G.; Calabria, S.; Barbieri, E.; Laaksonen, R.; Bonnet, F.; et al. Associations between specific plasma ceramides and severity of coronary-artery stenosis assessed by coronary angiography. Diabetes Metab. 2020, 46, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Hilvo, M.; Meikle, P.J.; Pedersen, E.R.; Tell, G.S.; Dhar, I.; Brenner, H.; Schottker, B.; Laaperi, M.; Kauhanen, D.; Koistinen, K.M.; et al. Development and validation of a ceramide- and phospholipid-based cardiovascular risk estimation score for coronary artery disease patients. Eur. Heart J. 2020, 41, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Poss, A.M.; Maschek, J.A.; Cox, J.E.; Hauner, B.J.; Hopkins, P.N.; Hunt, S.C.; Holland, W.L.; Summers, S.A.; Playdon, M.C. Machine learning reveals serum sphingolipids as cholesterol-independent biomarkers of coronary artery disease. J. Clin. Investig. 2020, 130, 1363–1376. [Google Scholar] [CrossRef]
- Michelucci, E.; Giorgi, N.D.; Finamore, F.; Smit, J.M.; Scholte, A.; Signore, G.; Rocchiccioli, S. Lipid biomarkers in statin users with coronary artery disease annotated by coronary computed tomography angiography. Sci. Rep. 2021, 11, 12899. [Google Scholar] [CrossRef]
- Di Giorgi, N.; Michelucci, E.; Smit, J.M.; Scholte, A.; El Mahdiui, M.; Knuuti, J.; Buechel, R.R.; Teresinska, A.; Pizzi, M.N.; Roque, A.; et al. A specific plasma lipid signature associated with high triglycerides and low HDL cholesterol identifies residual cad risk in patients with chronic coronary syndrome. Atherosclerosis 2021, 339, 1–11. [Google Scholar] [CrossRef]
- Meikle, P.J.; Wong, G.; Tsorotes, D.; Barlow, C.K.; Weir, J.M.; Christopher, M.J.; MacIntosh, G.L.; Goudey, B.; Stern, L.; Kowalczyk, A.; et al. Plasma lipidomic analysis of stable and unstable coronary artery disease. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2723–2732. [Google Scholar] [CrossRef]
- Ellims, A.H.; Wong, G.; Weir, J.M.; Lew, P.; Meikle, P.J.; Taylor, A.J. Plasma lipidomic analysis predicts non-calcified coronary artery plaque in asymptomatic patients at intermediate risk of coronary artery disease. Eur. Heart J. Cardiovasc. Imaging 2014, 15, 908–916. [Google Scholar] [CrossRef]
- Cheng, J.M.; Suoniemi, M.; Kardys, I.; Vihervaara, T.; de Boer, S.P.; Akkerhuis, K.M.; Sysi-Aho, M.; Ekroos, K.; Garcia-Garcia, H.M.; Oemrawsingh, R.M.; et al. Plasma concentrations of molecular lipid species in relation to coronary plaque characteristics and cardiovascular outcome: Results of the atheroremo-ivus study. Atherosclerosis 2015, 243, 560–566. [Google Scholar] [CrossRef]
- Meikle, P.J.; Formosa, M.F.; Mellett, N.A.; Jayawardana, K.S.; Giles, C.; Bertovic, D.A.; Jennings, G.L.; Childs, W.; Reddy, M.; Carey, A.L.; et al. HDL phospholipids, but not cholesterol distinguish acute coronary syndrome from stable coronary artery disease. J. Am. Heart Assoc. 2019, 8, e011792. [Google Scholar] [CrossRef]
- Karjalainen, J.P.; Mononen, N.; Hutri-Kahonen, N.; Lehtimaki, M.; Hilvo, M.; Kauhanen, D.; Juonala, M.; Viikari, J.; Kahonen, M.; Raitakari, O.; et al. New evidence from plasma ceramides links apoe polymorphism to greater risk of coronary artery disease in finnish adults. J. Lipid Res. 2019, 60, 1622–1629. [Google Scholar] [CrossRef]
- Ding, M.; Rexrode, K.M. A review of lipidomics of cardiovascular disease highlights the importance of isolating lipoproteins. Metabolites 2020, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Denimal, D.; Pais de Barros, J.P.; Petit, J.M.; Bouillet, B.; Verges, B.; Duvillard, L. Significant abnormalities of the HDL phosphosphingolipidome in type 1 diabetes despite normal HDL cholesterol concentration. Atherosclerosis 2015, 241, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Kostara, C.E.; Ferrannini, E.; Bairaktari, E.T.; Papathanasiou, A.; Elisaf, M.; Tsimihodimos, V. Early signs of atherogenic features in the HDL lipidomes of normolipidemic patients newly diagnosed with type 2 diabetes. Int. J. Mol. Sci. 2020, 21, 8835. [Google Scholar] [CrossRef] [PubMed]
- Kostara, C.E.; Karakitsou, K.S.; Florentin, M.; Bairaktari, E.T.; Tsimihodimos, V. Progressive, qualitative, and quantitative alterations in HDL lipidome from healthy subjects to patients with prediabetes and type 2 diabetes. Metabolites 2022, 12, 683. [Google Scholar] [CrossRef] [PubMed]
- Denimal, D.; Nguyen, A.; Pais de Barros, J.P.; Bouillet, B.; Petit, J.M.; Verges, B.; Duvillard, L. Major changes in the sphingophospholipidome of HDL in non-diabetic patients with metabolic syndrome. Atherosclerosis 2016, 246, 106–114. [Google Scholar] [CrossRef]
- Mocciaro, G.; D’Amore, S.; Jenkins, B.; Kay, R.; Murgia, A.; Herrera-Marcos, L.V.; Neun, S.; Sowton, A.P.; Hall, Z.; Palma-Duran, S.A.; et al. Lipidomic approaches to study HDL metabolism in patients with central obesity diagnosed with metabolic syndrome. Int. J. Mol. Sci. 2022, 23, 6786. [Google Scholar] [CrossRef]
- Orsoni, A.; Therond, P.; Tan, R.; Giral, P.; Robillard, P.; Kontush, A.; Meikle, P.J.; Chapman, M.J. Statin action enriches HDL3 in polyunsaturated phospholipids and plasmalogens and reduces LDL-derived phospholipid hydroperoxides in atherogenic mixed dyslipidemia. J. Lipid Res. 2016, 57, 2073–2087. [Google Scholar] [CrossRef]
- Kostara, C.E.; Tsimihodimos, V.; Elisaf, M.S.; Bairaktari, E.T. Nmr-based lipid profiling of high density lipoprotein particles in healthy subjects with low, normal, and elevated HDL-cholesterol. J. Proteome Res. 2017, 16, 1605–1616. [Google Scholar] [CrossRef]
- Chen, Y.; Wen, S.; Jiang, M.; Zhu, Y.; Ding, L.; Shi, H.; Dong, P.; Yang, J.; Yang, Y. Atherosclerotic dyslipidemia revealed by plasma lipidomics on apoe(-/-) mice fed a high-fat diet. Atherosclerosis 2017, 262, 78–86. [Google Scholar] [CrossRef]
- Takeda, H.; Izumi, Y.; Tamura, S.; Koike, T.; Koike, Y.; Shiomi, M.; Bamba, T. Lipid profiling of serum and lipoprotein fractions in response to pitavastatin using an animal model of familial hypercholesterolemia. J. Proteome Res. 2020, 19, 1100–1108. [Google Scholar] [CrossRef]
- Meikle, P.J.; Wong, G.; Tan, R.; Giral, P.; Robillard, P.; Orsoni, A.; Hounslow, N.; Magliano, D.J.; Shaw, J.E.; Curran, J.E.; et al. Statin action favors normalization of the plasma lipidome in the atherogenic mixed dyslipidemia of mets: Potential relevance to statin-associated dysglycemia. J. Lipid Res. 2015, 56, 2381–2392. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Mundra, P.A.; Straznicky, N.E.; Nestel, P.J.; Wong, G.; Tan, R.; Huynh, K.; Ng, T.W.; Mellett, N.A.; Weir, J.M.; et al. Weight loss and exercise alter the high-density lipoprotein lipidome and improve high-density lipoprotein functionality in metabolic syndrome. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Chapman, M.J.; Orsoni, A.; Tan, R.; Mellett, N.A.; Nguyen, A.; Robillard, P.; Giral, P.; Therond, P.; Meikle, P.J. LDL subclass lipidomics in atherogenic dyslipidemia: Effect of statin therapy on bioactive lipids and dense LDL. J. Lipid Res. 2020, 61, 911–932. [Google Scholar] [CrossRef] [PubMed]
- Padro, T.; Vilahur, G.; Sanchez-Hernandez, J.; Hernandez, M.; Antonijoan, R.M.; Perez, A.; Badimon, L. Lipidomic changes of LDL in overweight and moderately hypercholesterolemic subjects taking phytosterol- and omega-3-supplemented milk. J. Lipid Res. 2015, 56, 1043–1056. [Google Scholar] [CrossRef]
- Sutter, I.; Velagapudi, S.; Othman, A.; Riwanto, M.; Manz, J.; Rohrer, L.; Rentsch, K.; Hornemann, T.; Landmesser, U.; von Eckardstein, A. Plasmalogens of high-density lipoproteins (HDL) are associated with coronary artery disease and anti-apoptotic activity of HDL. Atherosclerosis 2015, 241, 539–546. [Google Scholar] [CrossRef]
- Lee, J.H.; Yang, J.S.; Lee, S.H.; Moon, M.H. Analysis of lipoprotein-specific lipids in patients with acute coronary syndrome by asymmetrical flow field-flow fractionation and nanoflow liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1099, 56–63. [Google Scholar] [CrossRef]
- Avela, H.F.; Siren, H. Advances in lipidomics. Clin. Chim. Acta Int. J. Clin. Chem. 2020, 510, 123–141. [Google Scholar] [CrossRef]
- Hancock-Cerutti, W.; Lhomme, M.; Dauteuille, C.; Lecocq, S.; Chapman, M.J.; Rader, D.J.; Kontush, A.; Cuchel, M. Paradoxical coronary artery disease in humans with hyperalphalipoproteinemia is associated with distinct differences in the high-density lipoprotein phosphosphingolipidome. J. Clin. Lipidol. 2017, 11, 1192–1200.e1193. [Google Scholar] [CrossRef]
- Giammanco, A.; Noto, D.; Barbagallo, C.M.; Nardi, E.; Caldarella, R.; Ciaccio, M.; Averna, M.R.; Cefalu, A.B. Hyperalphalipoproteinemia and beyond: The role of HDL in cardiovascular diseases. Life 2021, 11, 581. [Google Scholar] [CrossRef]
- Stary, H.C. Natural history and histological classification of atherosclerotic lesions: An update. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1177–1178. [Google Scholar] [CrossRef]
- Virmani, R.; Kolodgie, F.D.; Burke, A.P.; Farb, A.; Schwartz, S.M. Lessons from sudden coronary death: A comprehensive morphological classification scheme for atherosclerotic lesions. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1262–1275. [Google Scholar] [CrossRef] [PubMed]
- You, S.A.; Archacki, S.R.; Angheloiu, G.; Moravec, C.S.; Rao, S.; Kinter, M.; Topol, E.J.; Wang, Q. Proteomic approach to coronary atherosclerosis shows ferritin light chain as a significant marker: Evidence consistent with iron hypothesis in atherosclerosis. Physiol. Genom. 2003, 13, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Donners, M.M.; Verluyten, M.J.; Bouwman, F.G.; Mariman, E.C.; Devreese, B.; Vanrobaeys, F.; van Beeumen, J.; van den Akker, L.H.; Daemen, M.J.; Heeneman, S. Proteomic analysis of differential protein expression in human atherosclerotic plaque progression. J. Pathol. 2005, 206, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.J.; Ryang, Y.S.; Jang, S.W.; Lee, C.W.; Han, K.H.; Ko, J. Proteomic analysis of differential protein expression in atherosclerosis. Biomark. Biochem. Indic. Expo. Response Susceptibil. Chem. 2006, 11, 279–290. [Google Scholar] [CrossRef]
- Bagnato, C.; Thumar, J.; Mayya, V.; Hwang, S.I.; Zebroski, H.; Claffey, K.P.; Haudenschild, C.; Eng, J.K.; Lundgren, D.H.; Han, D.K. Proteomics analysis of human coronary atherosclerotic plaque: A feasibility study of direct tissue proteomics by liquid chromatography and tandem mass spectrometry. Mol. Cell. Proteom. 2007, 6, 1088–1102. [Google Scholar] [CrossRef]
- Terzuoli, L.; Felici, C.; Ciari, I.; Guerranti, R.; Pagani, R.; Marinello, E.; Porcelli, B. Synthetic gel of carotid artery plaque. Int. J. Immunopathol. Pharmacol. 2007, 20, 637–642. [Google Scholar] [CrossRef]
- Olson, F.J.; Sihlbom, C.; Davidsson, P.; Hulthe, J.; Fagerberg, B.; Bergstrom, G. Consistent differences in protein distribution along the longitudinal axis in symptomatic carotid atherosclerotic plaques. Biochem. Biophys. Res. Commun. 2010, 401, 574–580. [Google Scholar] [CrossRef]
- De Kleijn, D.P.; Moll, F.L.; Hellings, W.E.; Ozsarlak-Sozer, G.; de Bruin, P.; Doevendans, P.A.; Vink, A.; Catanzariti, L.M.; Schoneveld, A.H.; Algra, A.; et al. Local atherosclerotic plaques are a source of prognostic biomarkers for adverse cardiovascular events. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 612–619. [Google Scholar] [CrossRef]
- De la Cuesta, F.; Alvarez-Llamas, G.; Maroto, A.S.; Donado, A.; Zubiri, I.; Posada, M.; Padial, L.R.; Pinto, A.G.; Barderas, M.G.; Vivanco, F. A proteomic focus on the alterations occurring at the human atherosclerotic coronary intima. Mol. Cell. Proteom. 2011, 10, M110 003517. [Google Scholar] [CrossRef]
- Malaud, E.; Piquer, D.; Merle, D.; Molina, L.; Guerrier, L.; Boschetti, E.; Saussine, M.; Marty-Ane, C.; Albat, B.; Fareh, J. Carotid atherosclerotic plaques: Proteomics study after a low-abundance protein enrichment step. Electrophoresis 2012, 33, 470–482. [Google Scholar] [CrossRef]
- Hao, P.; Ren, Y.; Pasterkamp, G.; Moll, F.L.; de Kleijn, D.P.; Sze, S.K. Deep proteomic profiling of human carotid atherosclerotic plaques using multidimensional lc-ms/ms. Proteomics. Clin. Appl. 2014, 8, 631–635. [Google Scholar] [CrossRef]
- Malaud, E.; Merle, D.; Piquer, D.; Molina, L.; Salvetat, N.; Rubrecht, L.; Dupaty, E.; Galea, P.; Cobo, S.; Blanc, A.; et al. Local carotid atherosclerotic plaque proteins for the identification of circulating biomarkers in coronary patients. Atherosclerosis 2014, 233, 551–558. [Google Scholar] [CrossRef]
- Alonso-Orgaz, S.; Moreno-Luna, R.; Lopez, J.A.; Gil-Dones, F.; Padial, L.R.; Moreu, J.; de la Cuesta, F.; Barderas, M.G. Proteomic characterization of human coronary thrombus in patients with st-segment elevation acute myocardial infarction. J. Proteom. 2014, 109, 368–381. [Google Scholar] [CrossRef]
- Liang, W.; Ward, L.J.; Karlsson, H.; Ljunggren, S.A.; Li, W.; Lindahl, M.; Yuan, X.M. Distinctive proteomic profiles among different regions of human carotid plaques in men and women. Sci. Rep. 2016, 6, 26231. [Google Scholar] [CrossRef]
- Lee, R.; Fischer, R.; Charles, P.D.; Adlam, D.; Valli, A.; Di Gleria, K.; Kharbanda, R.K.; Choudhury, R.P.; Antoniades, C.; Kessler, B.M.; et al. A novel workflow combining plaque imaging, plaque and plasma proteomics identifies biomarkers of human coronary atherosclerotic plaque disruption. Clin. Proteom. 2017, 14, 22. [Google Scholar] [CrossRef]
- Li, J.; Liu, X.; Xiang, Y.; Ding, X.; Wang, T.; Liu, Y.; Yin, M.; Tan, C.; Deng, F.; Chen, L. Alpha-2-macroglobulin and heparin cofactor ii and the vulnerability of carotid atherosclerotic plaques: An itraq-based analysis. Biochem. Biophys. Res. Commun. 2017, 483, 964–971. [Google Scholar] [CrossRef]
- Langley, S.R.; Willeit, K.; Didangelos, A.; Matic, L.P.; Skroblin, P.; Barallobre-Barreiro, J.; Lengquist, M.; Rungger, G.; Kapustin, A.; Kedenko, L.; et al. Extracellular matrix proteomics identifies molecular signature of symptomatic carotid plaques. J. Clin. Investig. 2017, 127, 1546–1560. [Google Scholar] [CrossRef]
- Ward, L.J.; Olausson, P.; Li, W.; Yuan, X.M. Proteomics and multivariate modelling reveal sex-specific alterations in distinct regions of human carotid atheroma. Biol. Sex Differ. 2018, 9, 54. [Google Scholar] [CrossRef]
- Stakhneva, E.M.; Meshcheryakova, I.A.; Demidov, E.A.; Starostin, K.V.; Sadovski, E.V.; Peltek, S.E.; Voevoda, M.I.; Chernyavskii, A.M.; Volkov, A.M.; Ragino, Y.I. A proteomic study of atherosclerotic plaques in men with coronary atherosclerosis. Diagnostics 2019, 9, 177. [Google Scholar] [CrossRef]
- Hwang, S.I.; Thumar, J.; Lundgren, D.H.; Rezaul, K.; Mayya, V.; Wu, L.; Eng, J.; Wright, M.E.; Han, D.K. Direct cancer tissue proteomics: A method to identify candidate cancer biomarkers from formalin-fixed paraffin-embedded archival tissues. Oncogene 2007, 26, 65–76. [Google Scholar] [CrossRef]
- Duran, M.C.; Mas, S.; Martin-Ventura, J.L.; Meilhac, O.; Michel, J.B.; Gallego-Delgado, J.; Lazaro, A.; Tunon, J.; Egido, J.; Vivanco, F. Proteomic analysis of human vessels: Application to atherosclerotic plaques. Proteomics 2003, 3, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Martin-Ventura, J.L.; Duran, M.C.; Blanco-Colio, L.M.; Meilhac, O.; Leclercq, A.; Michel, J.B.; Jensen, O.N.; Hernandez-Merida, S.; Tunon, J.; Vivanco, F.; et al. Identification by a differential proteomic approach of heat shock protein 27 as a potential marker of atherosclerosis. Circulation 2004, 110, 2216–2219. [Google Scholar] [CrossRef] [PubMed]
- Duran, M.C.; Martin-Ventura, J.L.; Mohammed, S.; Barderas, M.G.; Blanco-Colio, L.M.; Mas, S.; Moral, V.; Ortega, L.; Tunon, J.; Jensen, O.N.; et al. Atorvastatin modulates the profile of proteins released by human atherosclerotic plaques. Eur. J. Pharmacol. 2007, 562, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Colio, L.M.; Martin-Ventura, J.L.; Munoz-Garcia, B.; Orbe, J.; Paramo, J.A.; Michel, J.B.; Ortiz, A.; Meilhac, O.; Egido, J. Identification of soluble tumor necrosis factor-like weak inducer of apoptosis (stweak) as a possible biomarker of subclinical atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 916–922. [Google Scholar] [CrossRef]
- De la Cuesta, F.; Barderas, M.G.; Calvo, E.; Zubiri, I.; Maroto, A.S.; Darde, V.M.; Martin-Rojas, T.; Gil-Dones, F.; Posada-Ayala, M.; Tejerina, T.; et al. Secretome analysis of atherosclerotic and non-atherosclerotic arteries reveals dynamic extracellular remodeling during pathogenesis. J. Proteom. 2012, 75, 2960–2971. [Google Scholar] [CrossRef]
- Aragones, G.; Auguet, T.; Guiu-Jurado, E.; Berlanga, A.; Curriu, M.; Martinez, S.; Alibalic, A.; Aguilar, C.; Hernandez, E.; Camara, M.L.; et al. Proteomic profile of unstable atheroma plaque: Increased neutrophil defensin 1, clusterin, and apolipoprotein e levels in carotid secretome. J. Proteome Res. 2016, 15, 933–944. [Google Scholar] [CrossRef]
- Stocker, R.; Keaney, J.F., Jr. Role of oxidative modifications in atherosclerosis. Physiol. Rev. 2004, 84, 1381–1478. [Google Scholar] [CrossRef]
- Steinberg, D.; Witztum, J.L. Oxidized low-density lipoprotein and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2311–2316. [Google Scholar] [CrossRef]
- Almofti, M.R.; Huang, Z.; Yang, P.; Rui, Y.; Yang, P. Proteomic analysis of rat aorta during atherosclerosis induced by high cholesterol diet and injection of vitamin d3. Clin. Exp. Pharmacol. Physiol. 2006, 33, 305–309. [Google Scholar] [CrossRef]
- Lepedda, A.J.; Nieddu, G.; Piperigkou, Z.; Kyriakopoulou, K.; Karamanos, N.; Formato, M. Circulating heparan sulfate proteoglycans as biomarkers in health and disease. Semin. Thromb. Hemost. 2021, 47, 295–307. [Google Scholar] [CrossRef]
- Qi, Y.X.; Qu, M.J.; Long, D.K.; Liu, B.; Yao, Q.P.; Chien, S.; Jiang, Z.L. Rho-gdp dissociation inhibitor alpha downregulated by low shear stress promotes vascular smooth muscle cell migration and apoptosis: A proteomic analysis. Cardiovasc. Res. 2008, 80, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Rodger, E.J.; Suetani, R.J.; Jones, G.T.; Kleffmann, T.; Carne, A.; Legge, M.; McCormick, S.P. Proteomic analysis of aortae from human lipoprotein(a) transgenic mice shows an early metabolic response independent of atherosclerosis. PLoS ONE 2012, 7, e30383. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Barallobre-Barreiro, J.; Jahangiri, M.; Mayr, M. Vascular proteomics in metabolic and cardiovascular diseases. J. Intern. Med. 2016, 280, 325–338. [Google Scholar] [CrossRef]
- Vaisar, T.; Hu, J.H.; Airhart, N.; Fox, K.; Heinecke, J.; Nicosia, R.F.; Kohler, T.; Potter, Z.E.; Simon, G.M.; Dix, M.M.; et al. Parallel Murine and Human Plaque Proteomics Reveals Pathways of Plaque Rupture. Circ. Res. 2020, 127, 997–1022. [Google Scholar] [CrossRef] [PubMed]
- Husi, H.; Van Agtmael, T.; Mullen, W.; Bahlmann, F.H.; Schanstra, J.P.; Vlahou, A.; Delles, C.; Perco, P.; Mischak, H. Proteome-based systems biology analysis of the diabetic mouse aorta reveals major changes in fatty acid biosynthesis as potential hallmark in diabetes mellitus-associated vascular disease. Circulation. Cardiovasc. Genet. 2014, 7, 161–170. [Google Scholar] [CrossRef] [PubMed]



| Lipoprotein Fractions | Purification | Proteomics | Main Findings | References | |
|---|---|---|---|---|---|
| ⁕ | Group 1: HDL from 20 healthy volunteers; Group 2: HDL3 from 6 controls and 7 CAD patients; Group 3: HDL3 from 32 controls and 32 CAD; HDL from atherosclerotic tissue | Ultracentrifugation/immunoaffinity chromatography | LC-ESI-MS/MS analysis | 48 proteins identified in HDL involved in lipid metabolism, complement regulation, proteinase inhibition, acute-phase response. Apo C-IV, paraoxonase-1, Complement C3, apo A-IV, apo E (confirmed by WB on HDL3 fraction from 64 subjects) enriched in HDL3 from CAD. More than 100 proteins identified in HDL from atherosclerotic plaque | [51] |
| ⁕ | Group 1: HDL3 from 6 CAD, 6 CAD + Statin/Niacin, 6 healthy controls; Group 2: HDL3 from 18 CAD, 18 CAD + Statin/Niacin. 12 months therapy | Sequential density ultracentrifugation | LC-FTICR MS | After 12 months of therapy, ↓ apo E, ↑ apo F and phospholipid transfer protein; combination therapy may revert CAD-associated changes in HDL3 protein composition | [77] |
| ⁕ | HDL from 7 hypercholesterolemia subjects and 9 normolipidemic subjects | Salt Gradient Ultracentrifugation | LC-MS/MS analysis | The quantitative HDL protein pattern correlates with the corresponding concentration and distribution of cholesterol from serum lipid measurements; hypercholesterolemia in unrelated individuals is the result of different deficiencies | [78] |
| ⁕ | HDL2 from 18 CAD and 20 controls | Sequential density ultracentrifugation | MALDI-TOF-TOF MS and pattern recognition analysis; LC-MALDI-TOF/TOF MS | HDL2 from CAD contains methionine sulfoxide residues in apolipoprotein A-I and elevated levels of apolipoprotein C-III; specific proteomic signatures of HDL2 accurately classify CAD and control subjects | [79] |
| ⁕ | HDL from 10 healthy men with low HDL cholesterol and 10 men with high HDL cholesterol levels challenged with endotoxin | Immunoaffinity chromatography (anti-apo A-I antibodies); density gradient ultracentrifugation | PS20 protein-chips (anti-apo A-I antibodies) coupled with SELDI-TOF MS analysis/2DE analysis | Profound changes in 21 markers (including SAA-1/2 and paraoxonase-1) were observed in the proteome in both groups; no relationship between baseline HDL cholesterol levels and HDL protein dynamics after endotoxin challenge | [80] |
| ⁕ | HDL from 39 new-onset AMI-patients vs 60 healthy individuals | Salt Gradient Ultracentrifugation | 2DE coupled with MALDI-TOF MS analysis | Altered glycosylation pattern in AMI-patients within the first 6 h after the onset of the event | [81] |
| ⁕ | HDL from 10 healthy controls, 10 CAD and 10 ACS | Sequential ultracentrifugation | 1DE coupled with LC-ESI-MS/MS analysis | 67 HDL-associated proteins identified; ↓ apo A-IV, ↑ SAA, ↑ Complement C3 in ACS vs. both CAD and controls | [82] |
| ⁕ | HDL from CAD, ACS, and healthy controls | Sequential ultracentrifugation or gel filtration chromatography | LC-ESI-MS/MS analysis | ↓ clusterin, ↑ apolipoprotein C-III in both CAD and ACS; HDL-proteome remodeling plays an important role for these altered functional properties of HDL (stimulation of proapoptotic pathways) | [83] |
| ⁕ | VLDL, LDL and HDL from four pooled plasma samples from 79 patients undergoing carotid endarterectomy and 57 healthy normolipidemic volunteers | Density gradient ultracentrifugation | 2DE coupled with MALDI-TOF-MS | ↑ acute-phase serum amyloid A protein (AP SAA) in all lipoprotein fractions, especially in LDL from atherosclerotic patients | [84] |
| ⁕ | HDL3 from 3 low HDL-c and 3 high HDL-c subjects | Density gradient ultracentrifugation | Top-Down Differential Mass Spectrometry: ETD LC-ESI-MS/MS | Proof-of-concept study; higher abundances of three apo C-III glycoforms in HDL3 from donors with low HDL-c | [85] |
| ⁕ | HDL from 10 congenital heart disease (CHD) patients vs. 10 controls | Sequential ultracentrifugation | iTRAQ technology coupled with nanoLC-MS/MS | 196 proteins identified, 5 upregulated proteins (inflammatory reactions) and 7 downregulated (lipid metabolism) proteins; HDL proteome changes to a pro-atherogenic profile in CHD patients | [86] |
| ⁕ | HDL2 and HDL3 from 40 ACS vs. 40 controls | Two-step discontinuous density-gradient ultracentrifugation | 2DE and 2DIGE analysis coupled with MALDI-TOF/TOF MS | 17 differentially expressed HDL-associated proteins identified, shifting to a dysfunctional HDL subfractions | [87] |
| ⁕ | HDL from diabetes and CVD | Sequential ultracentrifugation | MAP/LC-MRM/MS | 69 and 32 HDL proteins quantified, respectively, with multi-analyte profiling (MAP) and multiple reaction monitoring (MRM); high-throughput approach to examine changes in HDL proteins in diabetes and CVD | [88] |
| ⁕ | HDL from 21 CAD pre- and post- percutaneous transluminal coronary angioplasty (PTCA) | Immunoaffinity chromatography (anti-apo A-I antibodies) | Quantitative 16O/18O analysis/iTRAQ technology coupled with LC–MS/MS/system biology analysis | 225 identified proteins; high protein variability in HDL composition between individuals; post-PTCA increase in two protein clusters that included several apolipoproteins, fibrinogen-like protein 1 and other intracellular proteins, and a decrease in antithrombin-III, annexin A1 and several immunoglobulins | [89] |
| ⁕ | HDL from 20 subjects at risk for CAD: 10 patients had CAD and 10 did not | Sequential ultracentrifugation | Shotgun proteomic, glycomic, and ganglioside analyses using LC-MS | Combined HDL proteomic and glycomic profiles distinguished at-risk subjects with atherosclerosis from those without; CAD patients had ↓ apo A-I, ↓ apo A-II, ↓ apo E, ↓ SAA2, ↓ SAA4 (p = 0.007), ↑ sialylated glycans | [90] |
| ⁕ | HDL from 33 hypercholesterolemic subjects included in a clinical trial evaluating effects of virgin olive oil (VOO) and phenol-enriched VOO | Sequential density ultracentrifugation | Nano LC-MALDI MS/MS and nano LC-ORBITRAP-ESI MS/MS | 127 HDL-associated proteins identified; 15 differentially expressed proteins involved in liver X receptor-retinoid X receptor activation, acute phase response, and atherosclerosis; consumption of VOO or phenol-enriched VOOs, has an impact on the HDL proteome in a cardioprotective mode by upregulating proteins related to cholesterol homeostasis, protection against oxidation and blood coagulation while downregulating proteins implicated in acute-phase response, lipid transport, and immune response | [91] |
| ⁕ | HDL from 54 systemic Lupus Erythematosus (SLE) patients (with and without atherosclerotic plaque), 25 controls, patients with type 1 diabetes with or without coronary artery calcification | Sequential density gradient ultracentrifugation | Targeted proteomics (18 proteins): LC-ESI-MS/MS analysis | ↓ PON3 in HDL of both SLE and T1DM patients with subclinical atherosclerosis | [92] |
| ⁕ | LDL and HDL from 10 healthy individuals with normal LDL-C treated with rosuvastatin for 28 days | Size-exclusion chromatography | LC-ESI-MS/MS analysis | Among the differentially expressed proteins, α-1-antirypsin increased dramatically in HDL; enhanced HDL anti-inflammatory activity, contributing to the non-lipid lowering beneficial effects of statins | [93] |
| ⁕ | HDL from 5 chronic heart failure (CHF) patients vs. 5 controls | Sequential ultracentrifugation | SCX/RP LC–MS/MS | 494 proteins identified; 23 newly identified HDL-associated proteins; 223 bacterial peptides were found in both CHF and controls | [94] |
| ⁕ | HDL from 14 abdominal aortic aneurism (AAA) patients vs. 7 controls | Sequential ultracentrifugation | iTRAQ technology coupled with nanoLC-MS/MS | ↑ peroxiredoxin-6, ↑ class I histocompatibility antigen, ↑ retinol-binding protein 4, ↑ paraoxonase/arylesterase 1, ↓ α-2 macroglobulin, ↓ C4b-binding protein | [95] |
| ⁕ | HDL from 10 CVD vs. 7 controls | Two-step discontinuous density-gradient ultracentrifugation | nLC-MS/MS analysis | 118 proteins identified; 10 proteins positively associated with the combined level of persistent organic pollutants (POPs) or with highly chlorinated polychlorinated biphenyls (PCB) congeners. Among these, cholesteryl ester transfer protein and phospholipid transfer protein, as well as the inflammatory marker serum amyloid A, were found. The serum paraoxonase/arylesterase 1 activity was inversely associated with POPs. Pathway analysis demonstrated that upregulated proteins were associated with biological processes involving lipoprotein metabolism, while downregulated proteins were associated with processes such as negative regulation of proteinases, acute phase response, platelet degranulation, and complement activation | [96] |
| ⁕ | HDL from 126 subjects with clinical indication for a coronary computed tomography angiography (CCTA) | High-resolution size exclusion chromatography followed by phospholipid-associated proteins capture (Calcium Silicate Hydrate) | LC-ESI-MS/MS | 72 HDL-associated proteins detected in at least 75% of subjects; 13 proteins significantly associated with calcified plaque burden including cathelicidin antimicrobial peptide, gelsolin (GELS), kininogen-1 (KNG1), and paraoxonase-1 (inverse relationships), apo A-IV, vitamin D binding protein, alpha-2- macroglobulin, and apo C-II (positive relationships); 15 proteins significantly associated with non-calcified plaque burden including apo A-I, apo F, antithrombin III, and apo C-I (inverse relationships), serum amyloid A1, immunoglobulin heavy constant alpha 1, complement factor B, complement C2, complement C3, complement bC1s subcomponent (positive relationships); among the evaluated risk factors, BMI has the greatest overall impact on the protein composition of HDL | [97] |
| ⁕ | HDL from heart failure patients: cardiovascular deaths vs. survivors (1:1) | Calcium silicate matrix | nLC-MS/MS analysis | 647 proteins identified; 49 HDL proteins were significantly different; a set of 12 selected proteins predicted death with 76% accuracy | [98] |
| ⁕ | HDL from 943 participants without prevalent myocardial infarction referred for coronary angiography in the CASABLANCA study | 15NHis6Apo A-I was added to human serum, incubated, diluted, and then purified using PhyTips (Phynexus, San Jose, CA, USA), packed with Ni-NTA HisBind Superflow stationary phase | Targeted proteomics (apolipoprotein A-I, C-1, C-2, C-3, and C-4): LC-ESI-MS/MS analysis | An HDL apolipoproteomic score is associated with the presence of CAD, independent of circulating apo A-I and apo B concentrations and other conventional cardiovascular risk factors. Among individuals with CAD, this score may be independently associated with cardiovascular death | [99] |
| ⁕ | Apolipoprotein AI-associated lipoproteins from 231 healthy individuals and patients with CAD | Metal chelate affinity chromatography | Targeted proteomics (21 proteins): LC-MS/MS analysis | A multiplexed proteomic assay useful for the estimation of cholesterol efflux and CAD risk in the clinical laboratory | [100] |
| ⁕ | HDL from 8 patients with complete deficiency of cholesteryl ester transfer protein (CETP) vs. 8 normolipidemic healthy subjects | Sequential ultracentrifugation | LC-MS/MS analysis | 79 HDL-associated proteins identified, involved in lipid metabolism, protease inhibition, complement regulation, and acute-phase response, including 5 potential newly identified HDL-associated proteins; ↑ apo E, ↑ Complement C3, C4a, C4b, and C9, ↑ apo C-III | [101] |
| ⁕ | HDL from patients after acute ischemic stroke at 2 time points (24 h, 35 patients; 96 h, 20 patients) and from 35 control subjects | Sequential density ultracentrifugation | Data-dependent acquisition (DDA) mass spectrometry and parallel reaction monitoring (PRM) mass spectrometry | Some proteins involved in acute phase response and platelet activation were significantly altered in stroke HDL at 24 and 96 h (p < 0.05). Accordingly, cholesterol efflux capacity was reduced by 32% (p < 0.001) at both time points | [102] |
| ⁕ | HDL from 46 incident new CVD and 46 one-to-one matched controls, at various stages of CKD | Sequential ultracentrifugation | Targeted proteomics (31 proteins): LC-MS/MS analysis | PON1, PON3, LCAT, and apolipoprotein A-IV levels inversely associated with incident CVD events in CKD patients | [103] |
| ⁕ | HDL and LDL from pooled plasma samples from 75 patients undergoing carotid endarterectomy and 50 healthy normolipidemic volunteers | Density gradient ultracentrifugation | nanoLC-MS/MS | Protein signatures specific for patients with “hard” or “soft” carotid plaques | [56] |
| # | HDL from 27 ESRD vs. 19 healthy subjects | Salt Gradient Ultracentrifugation | LC-MS/MS Analysis | 35 HDL-associated proteins identified; antitrypsin, retinol-binding protein 4 (RBP4), transthyretin, apo A-VI, and further minor proteins were exclusively detected in uremic HDL; ↑ SAA1, ↑albumin, ↑lipoprotein-associated phospholipase A2, ↑ apo C-III, ↓ apo A-I, ↓ apo A-II | [104] |
| # | HDL from 7 chronic hemodialysis patients vs. 7 healthy controls | Sequential density ultracentrifugation | iTRAQ technology coupled with 2D nano-LC-MALDI-TOF/TOF MS | 122 proteins identified, 40 proteins differentially expressed | [105] |
| # | HDL from 34 ESRD vs. 17 healthy controls | Sequential density ultracentrifugation | HPLC coupled with MALDI-TOF-TOF MS | SAA is enriched in HDL from ESRD patients correlating with its reduced anti-inflammatory capacity | [106] |
| # | HDL from 10 ESRD vs. 10 healthy controls | Sequential ultracentrifugation | 1DE coupled with LC-ESI-MS/MS analysis | 49 HDL-associated proteins identified; ↑ surfactant protein B (SP-B), ↑ apolipoprotein C-II, ↑ serum amyloid A (SAA), ↑ α-1-microglobulin/bikunin precursor; SAA promotes inflammatory cytokine production | [107] |
| # | HDL from 40 ESRD vs. 20 controls | Sequential ultracentrifugation | LC-ESI-MS/MS analysis | 63 identified proteins; ↑ 22 proteins, ↓ 6 proteins; HDL proteome is extensively remodeled in uremic subjects | [108] |
| # | HDL from 509 CKD; estimated glomerular filtration rate (eGFR) > 60 mL/min/1.73 m2 vs. eGFR < 15 mL/min/1.73 m2 | Two-step density gradient ultracentrifugation | nLC-MS/MS analysis (targeted proteomics: quantification of 38 HDL-proteins) | ↑ retinol binding protein 4, ↑ apo C-III, ↑ apo L1, ↓ vitronectin | [109] |
| # | HDL from 143 patients starting hemodialysis vs. 110 patients with advanced CKD | Two-step density gradient ultracentrifugation | nLC-MS/MS analysis (targeted proteomics: quantification of 38 HDL-proteins) | ↑ serum amyloid A1, A2, and A4, ↑ hemoglobin-b, ↑ haptoglobin-related protein, ↑ cholesteryl ester transfer protein, ↑ phospholipid transfer protein, ↑ apo E; hemodialysis initiation is associated with higher concentrations of HDL-proteins related to inflammation and lipid metabolism | [110] |
| # | HDL from 9 non-diabetic hemodialysis patients vs. 8 control patients | Sequential ultracentrifugation | nLC-Quadrupole-Orbitrap-MS | 326 proteins identified; ↑ 10 proteins (UDP-glucose: glycoprotein glucosyltransferase 1, Beta-2-microglobulin, Pulmonary surfactant-associated protein B, Protein AMBP, Insulin-like growth factor II, Immunoglobulin heavy constant alpha 2, Immunoglobulin lambda constant 2, HLA class I histocompatibility antigen B-58 alpha chain, Complement factor D, Inter-alpha-trypsin inhibitor heavy chain H1), ↓ 9 proteins (Guanylin, Calpain-1 catalytic subunit, Keratin, type I cytoskeletal, Ras-related protein Rab-6B, Ganglioside GM2 activator, Prostaglandin-H2 D-isomerase, Secretoglobin family 3A member 2, Thioredoxin-dependent peroxide reductase mitochondrial, Solute carrier family 2 facilitated glucose transporter member 2) involved in lipid metabolism, hemostasis, wound healing, oxidative stress, and apoptosis pathways | [111] |
| § | HDL from 30 T1DM vs. 30 controls | Single Vertical Spin Density Gradient Ultracentrifugation | LC-ESI-MS/MS analysis | Compromised HDL anti-oxidant functions due to higher abundance of irreversible PTMs of proteins in T1DM | [112] |
| § | HDL from 191 T1DM subjects | Sequential ultracentrifugation | Targeted proteomics (46 proteins): LC-ESI-MS/MS analysis | 8 proteins associated with albuminuria including AMBP (α1-microglobulin/bikunin precursor) and PTGDS (prostaglandin-H2 D-isomerase) that strongly and positively associated with the albumin excretion rate. PON1 and PON3 levels in HDL strongly and negatively associated with the presence of coronary artery calcium. Only PON1, associated with both albumin excretion rate and coronary artery calcification. The HDL proteome is remodeled in type 1 diabetes mellitus subjects with albuminuria | [113] |
| § | HDL from 26 patients with T1DM vs. 13 healthy controls | High-resolution size exclusion chromatography followed by phospholipid-associated proteins capture (Calcium Silicate Hydrate) | nLC-TripleTOF-MS | 78 HDL-bound proteins were measured; youths with T1DM show proteomic alterations of their HDL compared to healthy controls, despite similar concentration of HDL cholesterol. The influences of these compositional changes on HDL function are not yet known | [114] |
| § | HDL from two cohorts of T1DM patients (n = 47, n = 100). T1DM without complications vs. T1DM with complications | Sequential density ultracentrifugation | Targeted proteomics (35 proteins): LC-MS/MS analysis | Elevated concentrations of M-HDL particles and elevated levels of HDL-associated PON1 may contribute to long-term protection from the vascular complications of diabetes by pathways that are independent of total cholesterol and HDL cholesterol. ↑ apo C-I, apo E, complement C3, apo C-II, apo M, and PON1, ↓ apo A-IV and retinol-binding protein 4 (RBP4). In the female subgroup analysis: ↑ apo C-I and PON1, ↓ apo A-IV. Targeted proteomics in the replication cohort: ↑ PON1 | [115] |
| § | HDL from 9 newly diagnosed T2DM patients and 8 healthy controls | Immunoaffinity chromatography | LC-MS/MS | Plasma adiponectin levels were reduced in subjects with T2DM, which was directly associated with suppressed ABCA1-dependent cholesterol efflux capacity of HDL. The fractional catabolic rates of HDL cholesterol, apo A-II, apo J, apo A-IV, transthyretin, complement C3, and vitamin D-binding protein (all p < 0.05) were increased in subjects with T2D. Despite the increased HDL flux of acute-phase HDL proteins, there was no change in the proinflammatory index of HDL. Although lecithin-cholesterol acyl transferase (LCAT) and CETP activities were not affected in subjects with T2DM, LCAT was inversely associated with blood glucose and CETP was inversely associated with plasma adiponectin. The degradation rates of apo A-II and apo A-IV were correlated with hemoglobin A1c. In conclusion, there were in vivo impairments in HDL proteome dynamics and HDL metabolism in diet-controlled patients with T2DM | [116] |
| § | HDL from obese adolescents with T1DM (25 treated with metformin as adjunct therapy and 10 with placebo): double-blind, placebo-controlled clinical trial | Size-exclusion chromatography followed by lipophilic absorption | Data-independent acquisition (DIA) mass spectrometry | Two proteins out of eighty-four were significantly affected by metformin treatment, peptidoglycan recognition protein 2 and alpha-2-macroglobulin. Metformin did not significantly affect cholesterol efflux capacity (CEC). Changes in affected HDL proteins did not correlate with CEC | [117] |
| § | HDL2 and HDL3 from case–control study that enrolled 50 T1DM and 30 non-diabetes control individuals | Discontinuous density ultracentrifugation | 45 proteins quantified by parallel reaction monitoring | 13 proteins in HDL2 and 33 in HDL3 were differentially expressed in T1DM. Six proteins related to lipid metabolism, one to inflammatory acute phase, one to complement system and one to antioxidant response were more abundant in HDL2, while fourteen lipid metabolism, three acute-phase, three antioxidants and one transport were found in HDL3 of T1DM subjects. Three proteins (lipid metabolism, transport, and unknown function) were more abundant in HDL2, and ten (lipid metabolism, transport, protease inhibition) were more abundant in HDL3 of controls | [118] |
| ǂ | HDL from C57BL/6 mice challenged with intraperitoneal endotoxin vs. controls injected with saline | Sequential ultracentrifugation | Two-dimensional gel electrophoresis (2DE) coupled with LC-MS/MS analysis | ↑ SAA, apo E, apo A-IV, apo A-V; ↓ apo A-I, apo A-II | [119] |
| ǂ | HDL from C57BL/6j mice following 24 weeks on saturated fatty acids (SFA)- or monounsaturated fatty acids (MUFA)-enriched high-fat diets (HFDs) or low-fat diet | Fast protein liquid chromatography (FPLC) | nLC-ESI-MS/MS analysis | HDL particles were enriched with acute-phase proteins (serum amyloid A, haptoglobin, and hemopexin) and depleted in paraoxonase-1 after SFA-HFD in comparison with MUFA-HFD | [120] |
| ǂ | HDL from knock-out (for apo A-I, apo A-II or apo A-IV) and wild type C57BL/6J mice | High-resolution size exclusion chromatography followed by phospholipid-associated proteins capture (Calcium Silicate Hydrate) | LC-ESI-MS/MS analysis | The loss of apo A-I or apo A-II massively reduced HDL lipids and changed the plasma size pattern and/or abundance of several plasma proteins. Surprisingly though, many HDL proteins were not affected, suggesting they assemble on lipid particles in the absence of apo A-I or apo A-II. In contrast, apo A-IV ablation had minor effects on plasma lipids and proteins, suggesting that it forms particles that largely exclude other apolipoproteins. Overall, the data indicate that distinct HDL subpopulations exist, and that they do not contain, nor depend on, apo A-I, apo A-II or apo A-IV, and these contribute substantially to the proteomic diversity of HDL | [93] |
| ǂ | HDL from Scavenger receptor class B type I−/− mice vs. wildtype (WT) mice | Apo B containing lipoproteins precipitation followed by ultracentrifugation | LC–MS/MS analysis/2DE analysis | ↓ apo A-I, ↓ PON1, ↑ SAA, ↑ apo A-IV, ↑ α-1-antitrypsin; impaired cholesterol homeostasis in macrophages, and reduced anti-oxidative and anti-inflammatory effects | [121] |
| Samples | Proteomics | Main Findings | References |
|---|---|---|---|
| 10 coronary arteries from CAD vs. 7 coronary arteries from normal individuals | 2DE coupled with LC-MS/MS | Increased expression of the ferritin light chain in CAD (1.41 vs. 0.75; p = 0.01) | [156] |
| 5 stable carotid plaques vs. 6 lesions with a thrombus | 2DE coupled with MALDI-TOF/TOF MS | Expression of six isoforms of alpha1 anti-trypsin (AAT) in advanced plaques, one of which was uniquely expressed in thrombus-containing plaques | [157] |
| 7 atherosclerotic aortas vs. 7 normal aortas specimens from the same patients | 2DE coupled with MALDI TOF MS | 27 differentially expressed proteins identified involved in a number of biological processes, including calcium-mediated processes, migration of vascular smooth muscle cells, matrix metalloproteinase activation and regulation of pro-inflammatory cytokines | [158] |
| 35 human coronary atherosclerotic specimens (frozen, embedded, LCM specimens) | LC-MS/MS | 806 proteins identified; first large-scale proteomics map of human coronary atherosclerotic plaques | [159] |
| 10 carotid plaques | 2DE | Synthetic gel from 2DE | [160] |
| 29 unstable carotid plaques vs. 19 stable plaques | 2DE coupled with MALDI TOF MS | 57 spots identified ↓ SOD3, GST, HSP27, HSP20, annexin A10, and Rho GDI ↑ ferritin light subunit, SOD2 and fibrinogen fragment D | [6] |
| 10 complicated segments in the internal carotid artery vs. more stable segments in the common carotid artery of the same patients | 2-D DIGE coupled with nano-LC ESI MS/MS | ↑ Protein S100A10, Fibrinogen beta, Ferritin light chain, AAT, Protein S100-A9, Serum albumin ↓ Apolipoprotein E, Actin, cytoplasmic 1, L-Lactate dehydrogenase B, Myosin regulatory light polypeptide 9, Acetyl-CoA acetyltransferase, mitochondrial, Actin, aortic smooth muscle, Transgelin, Calponin-1, Tropomyosin beta, Annexin A5, Myosin light polypeptide 6 | [161] |
| 80 carotid plaques from patients who had a cardiovascular event vs. 80 carotid plaques from patients who remained stable during follow-up | Two-dimensional LC-MS/MS | Plaque osteopontin levels in single lesions are predictive for cardiovascular events in other vascular territories. Local atherosclerotic plaques are a source of prognostic biomarkers with a high predictive value for secondary manifestations of atherosclerotic disease | [162] |
| Intimal proteome from the human atherosclerotic coronary artery (LCM) vs. preatherosclerotic coronary vs. radial arteries | 2-D DIGE coupled with MALDI TOF/TOF MS | 13 proteins were differentially expressed (7 upregulated and 6 downregulated), and are implicated in the migrative capacity of vascular smooth muscle cells, extracellular matrix composition, coagulation, apoptosis, heat shock response, and intraplaque hemorrhage deposition | [163] |
| Noncomplicated and complicated plaques from carotid atherosclerotic lesions | Enrichment step with Proteominer. 1DE and 2DE coupled with nLC-MS/MS | Novel low-abundance proteins were associated very well with biological alterations related to atherosclerosis | [164] |
| 38 carotid atherosclerotic plaques pooled into two samples and analyzed in triplicate | Multidimensional LC-MS/MS | A total of 4702 proteins were identified from atherosclerotic plaques at a false discovery rate (FDR) of 1%, of which 3846 were identified with at least 2 unique peptides | [165] |
| Fibrotic and hemorrhagic carotid atherosclerotic plaques from coronary patients | Enrichment step with Proteominer. 2DE coupled with nLC-MS/MS | 118 proteins, differentially expressed in fibrotic and hemorrhagic plaques, allowed for the identification of three biological processes related to atherosclerosis (platelet degranulation, vascular autophagy and negative regulation of fibrinolysis). Combinations of such circulating biomarkers could be used to stratify coronary patients | [166] |
| Human coronary thrombus | 2-DE followed by MALDI MS/MS 1-DE followed by LC–MALDI-MS/MS 1-DE followed by LC–ESI-MS/MS) | 708 proteins were identified in the thrombus. A positive correlation of 5 proteins (fermitin homolog 3, thrombospondin-1, myosin-9, beta parvin and ras-related protein Rap-1b) with CD41 was found, pointing out the potential activation of a focal adhesion pathway within thrombus platelets | [167] |
| Different regions of human carotid plaques from men and women | 2-DE followed by MALDI-TOF MS/nLC-MS/MS | Expression levels of 18 proteins were significantly altered in plaque regions compared to the internal control region. In total, 9 proteins showed site-specific alterations, irrespective of gender, with clear associations to extracellular matrix remodeling; 5 proteins displayed gender-specific alterations with 2-DE, with two alterations validated by nLC-MS/MS. Gender differences in ferritin light chain and transthyretin were validated using both techniques. The validation of immunohistochemistry confirmed significantly higher levels of ferritin in plaques from male patients | [168] |
| 23 lipid-rich plaques and 13 non-lipid-rich plaques | LC MS/MS | Library of 423 proteins that are present in coronary plaque debris; MMP-9 and another 5 plaque-enriched proteins (lipopolysaccharide binding protein, Annexin A5, eukaryotic translocation initiation factor, syntaxin 11, cytochrome B5 reductase 3) significantly enriched in plaque and in plasma after plaque disruption | [169] |
| 28 vulnerable carotid plaques vs. 30 stable plaques | iTRAQ nanoLC MS/MS | 28 proteins were differentially expressed, including alpha-2-macroglobulin (a2M) and heparin cofactor II (HCII). The plasma level of a2M was found to be positively correlated, while HCII level was negatively correlated, with the higher vulnerability of carotid plaques. Both proteins were efficient in differentiating stable and vulnerable carotid plaques | [170] |
| Human carotid endarterectomy specimens from 6 symptomatic versus 6 asymptomatic patients | (Three steps extraction of carotid plaque ECM) nanoLC MS/MS | A 4-biomarker signature (matrix metalloproteinase 9, S100A8/S100A9, cathepsin D, and galectin-3-binding protein) improved risk prediction and was successfully replicated in an independent cohort in the SAPHIR study | [171] |
| Human coronary arteries (n = 100) and aortas (n = 100) | LC MS/MS | Identified hundreds of proteins (n = 1925) and numerous networks and pathways that are associated with early atherosclerosis | [11] |
| Carotid plaques from 10 men vs. 10 women | LC MS/MS | Sex-specific differences were identified both in general levels and from the lesion region-specific perspective. In this study, men were shown to have greater levels of inflammatory response proteins like lysozyme C and sPLA2, and women to have greater levels of serine protease inhibitors and afamin. These differences in the proteome may be suggestive of women developing plaques with a lower inflammatory profile, and greater stability, than men | [172] |
| Coronary arteries at different stages of development, from 15 patients | 2-DE followed by MALDI-TOF MS | The amounts of the following proteins were increased in stable atherosclerotic plaques at the stage of lipidosis and fibrosis: vimentin, tropomyosin β-chain, actin, keratin, tubulin β-chain, microfibril-associated glycoprotein 4, serum amyloid P-component, and annexin 5. In plaques at the stage of fibrosis and calcification, the amounts of mimecan and fibrinogen were increased. In the unstable atherosclerotic plaque of the necrotic–dystrophic type, the amounts of human serum albumin, mimecan, fibrinogen, serum amyloid P-component and annexin were increased | [173] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nieddu, G.; Formato, M.; Lepedda, A.J. Searching for Atherosclerosis Biomarkers by Proteomics: A Focus on Lesion Pathogenesis and Vulnerability. Int. J. Mol. Sci. 2023, 24, 15175. https://doi.org/10.3390/ijms242015175
Nieddu G, Formato M, Lepedda AJ. Searching for Atherosclerosis Biomarkers by Proteomics: A Focus on Lesion Pathogenesis and Vulnerability. International Journal of Molecular Sciences. 2023; 24(20):15175. https://doi.org/10.3390/ijms242015175
Chicago/Turabian StyleNieddu, Gabriele, Marilena Formato, and Antonio Junior Lepedda. 2023. "Searching for Atherosclerosis Biomarkers by Proteomics: A Focus on Lesion Pathogenesis and Vulnerability" International Journal of Molecular Sciences 24, no. 20: 15175. https://doi.org/10.3390/ijms242015175