

Noncoding RNAs: Master Regulator of Fibroblast to Myofibroblast Transition in Fibrosis

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Overview of Non-Coding RNA (ncRNA)
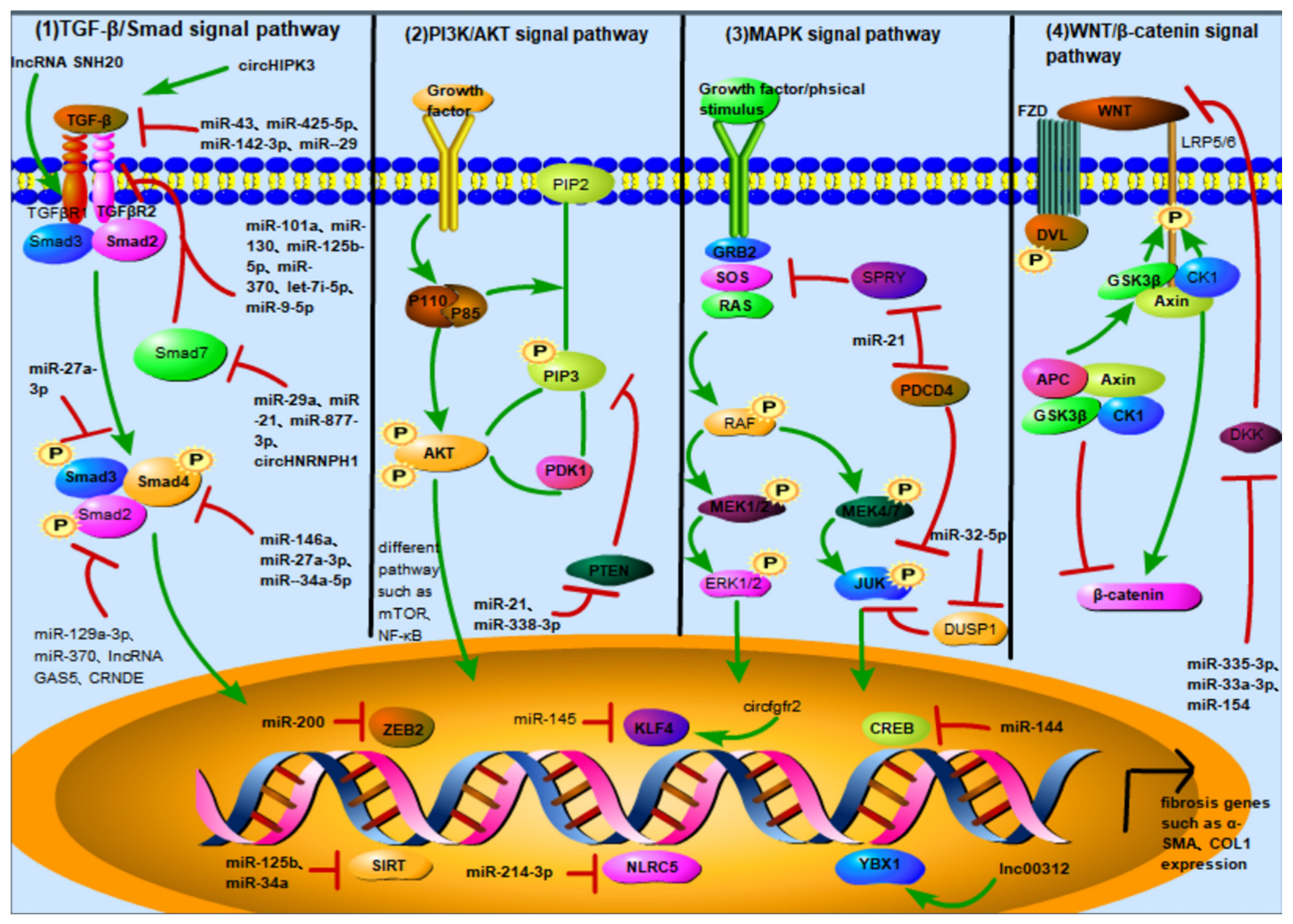
3. ncRNA Activates or Inhibits Different Signaling Pathways to Regulate FMT
3.1. TGF-β/Smad Signaling Pathway
ncRNA in the TGF-β/Smad Pathway
- TGF-β as the downstream target of ncRNA
- TGFβR as the downstream target of ncRNA
- Smads family as the downstream target of ncRNA
3.2. PI3K/AKT Signaling Pathway
ncRNA in the PI3K/AKT Pathway
3.3. MAPK Signaling Pathway
ncRNA in the MAPK Pathway
3.4. Wnt/β-Catenin Signaling Pathway
ncRNA in the Wnt/β-Catenin Pathway
3.5. Other Signaling Pathways
3.5.1. JAK/STAT Signaling Pathway
3.5.2. Notch Signaling
4. ncRNA Regulates FMT by Directly Affecting the Expression of Nuclear Transcription Factors
5. Conclusions and Prospective
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hinz, B.; Lagares, D. Evasion of apoptosis by myofibroblasts: A hallmark of fibrotic diseases. Nat. Rev. Rheumatol. 2020, 16, 11–31. [Google Scholar] [CrossRef] [PubMed]
- Rockey, D.C.; Bell, P.D.; Hill, J.A. Fibrosis—A Common Pathway to Organ Injury and Failure. N. Engl. J. Med. 2015, 372, 1138–1149. [Google Scholar] [CrossRef] [PubMed]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Guan, H.; Liu, J.-Q.; Zheng, Z.; Zhou, Q.; Zhang, H.; Su, L.-L.; Hu, D.-H. Hypoxia drives the transition of human dermal fibroblasts to a myofibroblast-like phenotype via the TGF-beta 1/Smad3 pathway. Int. J. Mol. Med. 2017, 39, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, J.; Zhou, Q.; Wang, H.; Xie, S.; Yang, X.; Ji, P.; Zhang, W.; He, T.; Liu, Y.; et al. Linagliptin inhibits high glucose-induced transdifferentiation of hypertrophic scar-derived fibroblasts to myofibroblasts via IGF/Akt/mTOR signalling pathway. Exp. Dermatol. 2019, 28, 19–27. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Y.; Wang, L.; Hou, C.; An, M. The effects of pressure intervention on wound healing and scar formation in a Bama minipig model. Burns 2019, 45, 413–422. [Google Scholar] [CrossRef]
- Gurtner, G.C.; Dauskardt, R.H.; Wong, V.W.; Bhatt, K.A.; Wu, K.; Vial, I.N.; Padois, K.; Korman, J.M.; Longaker, M.T. Improving Cutaneous Scar Formation by Controlling the Mechanical Environment Large Animal and Phase I Studies. Ann. Surg. 2011, 254, 217–225. [Google Scholar] [CrossRef]
- Plotkin, L.I.; Wallace, J.M. MicroRNAs and osteocytes. Bone 2021, 150, 115994. [Google Scholar] [CrossRef]
- Pu, M.; Chen, J.; Tao, Z.; Miao, L.; Qi, X.; Wang, Y.; Ren, J. Regulatory network of miRNA on its target: Coordination between transcriptional and post-transcriptional regulation of gene expression. Cell Mol. Life Sci. 2019, 76, 441–451. [Google Scholar] [CrossRef]
- Wu, M.; Fu, P.; Qu, L.; Liu, J.; Lin, A. Long Noncoding RNAs, New Critical Regulators in Cancer Immunity. Front. Oncol. 2020, 10, 550987. [Google Scholar] [CrossRef]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, S.; Tao, M.; Shen, X.; Ju, S. Translatable circRNAs and lncRNAs: Driving mechanisms and functions of their translation products. Cancer Lett. 2020, 483, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, B.; Bin, X.; Xie, C.; Li, B.; Liu, O.; Tang, Z. CircHIPK3: Key Player in Pathophysiology and Potential Diagnostic and Therapeutic Tool. Front. Med. 2021, 8, 615417. [Google Scholar] [CrossRef] [PubMed]
- Hanniford, D.; Ulloa-Morales, A.; Karz, A.; Berzoti-Coelho, M.G.; Moubarak, R.S.; Sanchez-Sendra, B.; Kloetgen, A.; Davalos, V.; Imig, J.; Wu, P.; et al. Epigenetic Silencing of CDR1as Drives IGF2BP3-Mediated Melanoma Invasion and Metastasis. Cancer Cell 2020, 37, 55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhao, K.; Xu, X.; Yang, Y.; Yan, S.; Wei, P.; Liu, H.; Xu, J.; Xiao, F.; Zhou, H.; et al. A peptide encoded by circular form of LINC-PINT suppresses oncogenic transcriptional elongation in glioblastoma. Nat. Commun. 2018, 9, 4475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Yang, L.; Chen, L.-L. The Biogenesis, Functions, and Challenges of Circular RNAs. Mol. Cell. 2018, 71, 428–442. [Google Scholar] [CrossRef] [Green Version]
- Salzman, J.; Chen, R.E.; Olsen, M.N.; Wang, P.L.; Brown, P.O. Cell-Type Specific Features of Circular RNA Expression. PLos Genet. 2013, 9, E1003777. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Yu, F.; Li, X.; Gao, H.; Li, P. CircHIPK3 Plays Vital Roles in Cardiovascular Disease. Front. Cardiovasc. Med. 2021, 8, 733248. [Google Scholar] [CrossRef]
- Xie, Y.; Yuan, X.; Zhou, W.; Kosiba, A.A.; Shi, H.; Gu, J.; Qin, Z. The circular RNA HIPK3 (circHIPK3) and its regulation in cancer progression: Review. Life Sci. 2020, 254, 117252. [Google Scholar] [CrossRef]
- Wilson, S.E. TGF beta-1,-2 and-3 in the modulation of fibrosis in the cornea and other organs. Exp. Eye Res. 2021, 207, 108594. [Google Scholar] [CrossRef]
- Saadat, S.; Noureddini, M.; Mahjoubin-Tehran, M.; Nazemi, S.; Shojaie, L.; Aschner, M.; Maleki, B.; Abbasi-kolli, M.; Rajabi Moghadam, H.; Alani, B.; et al. Pivotal Role of TGF-beta/Smad Signaling in Cardiac Fibrosis: Non-coding RNAs as Effectual Players. Front. Cardiovasc. Med. 2021, 7, 588347. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, Y.; Qiu, Z.; Zhao, R.; Liu, Z.; Chen, W.; Ge, J.; Shi, B. CircHIPK3 regulates cardiac fibroblast proliferation, migration and phenotypic switching through the miR-152-3p/TGF-beta 2 axis under hypoxia. PeerJ 2020, 8, 9796. [Google Scholar] [CrossRef] [PubMed]
- Tho, L.; Bei, Y.; Chen, P.; Lei, Z.; Fu, S.; Zhang, H.; Xu, J.; Che, L.; Chen, X.; Sluijter, J.P.G.; et al. Crucial Role of miR-433 in Regulating Cardiac Fibrosis. Theranostics 2016, 6, 2068–2083. [Google Scholar] [CrossRef] [Green Version]
- Duan, M.; Zhang, Y.; Zhang, H.; Meng, Y.; Qian, M.; Zhang, G. Epidermal stem cell-derived exosomes promote skin regeneration by downregulating transforming growth factor-beta 1 in wound healing. Stem. Cell Res. Ther. 2020, 11, 452. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Chao, G.; Li, W.; Zhu, J.; Li, F.; Qi, B.; Wei, Y.; Chen, S.; Zhou, G.; Lu, X.; et al. Activated CD4(+) T cells-derived exosomal miR-142-3p boosts post-ischemic ventricular remodeling by activating myofibrobiast. Aging 2020, 12, 7380–7396. [Google Scholar] [CrossRef]
- Liang, C.; Bu, S.; Fan, X. Suppressive effect of microRNA-29b on hepatic stellate cell activation and its crosstalk with TGF-beta 1/Smad3. Cell Biochem. Funct. 2016, 34, 326–333. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Bao, C.; Guo, W.; Li, S.; Chen, J.; Chen, B.; Luo, Y.; Lyu, D.; Li, Y.; Shi, G.; et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat. Commun. 2016, 7, 11215. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.M.; Wilusz, J.E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev. 2014, 28, 2233–2247. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Sun, H. Biogenesis, cellular effects, and biomarker value of circHIPK3. Cancer Cell Int. 2021, 21, 256. [Google Scholar] [CrossRef]
- Xu, Q.; Cheng, D.; Li, G.; Liu, Y.; Li, P.; Sun, W.; Ma, D.; Ni, C. CircHIPK3 regulates pulmonary fibrosis by facilitating glycolysis in miR-30a-3p/FOXK2-dependent manner. Int. J. Biol. Sci. 2021, 17, 2294–2307. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-X.; Lu, J.; Xie, H.; Wang, D.-p.; Ni, H.-E.; Zhu, Y.; Ren, L.-H.; Meng, X.-X.; Wang, R.-L. circHIPK3 regulates lung fibroblast-to-myofibroblast transition by functioning as a competing endogenous RNA. Cell Death Dis. 2019, 10, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, J.-Y.; Ma, L.-M.; Guo, Y.-H.; Zhang, Y.-C.; Zhou, H.; Shao, P.; Chen, Y.-Q.; Qu, L.-H. Deep Sequencing of Human Nuclear and Cytoplasmic Small RNAs Reveals an Unexpectedly Complex Subcellular Distribution of miRNAs and tRNA 3’ Trailers. PLoS ONE 2010, 5, e0010563. [Google Scholar] [CrossRef] [Green Version]
- Dohner, K.; Brown, J.; Hehmann, U.; Hetzel, C.; Stewart, J.; Lowther, G.; Scholl, C.; Frohling, S.; Cuneo, A.; Tsui, L.C.; et al. Molecular cytogenetic characterization of a critical region in bands 7q35-q36 commonly deleted in malignant myeloid disorders. Blood 1998, 92, 4031–4035. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.J.; Factora, T.D.; Dey, S.; Kota, J. 1 A Systematic Review of miR-29 in Cancer. Mol. Ther.-Oncolytics 2019, 12, 173–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, R.; Dai, X.; Li, Y.; Li, C.; Liu, L. Exosomes from miR-29a-modified adipose-derived mesenchymal stem cells reduce excessive scar formation by inhibiting TGF-beta 2/Smad3 signaling. Mol. Med. Rep. 2021, 24, 12398. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-C.; Liao, Y.-W.; Hsieh, P.-L.; Chang, Y.-C. Targeting lncRNA H19/miR-29b/COL1A1 Axis Impedes Myofibroblast Activities of Precancerous Oral Submucous Fibrosis. Int. J. Mol. Sci. 2021, 22, 2216. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y.; Huang, L.; Lan, Y.J.; Zhang, Q. The inhibition effect of miR-29c on lung fibroblasts transdifferentiation induced by SiO(2). Chin. J. Ind. Hyg. Occup. Dis. 2019, 37, 321–326. [Google Scholar] [CrossRef]
- Zhu, Y.; Feng, Z.; Jian, Z.; Xiao, Y. Long noncoding RNA TUG1 promotes cardiac fibroblast transformation to myofibroblasts via miR-29c in chronic hypoxia. Mol. Med. Rep. 2018, 18, 3451–3460. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Wang, K.; Liao, Y.; Zeng, Q.; Li, Y.; Hu, F.; Liu, Y.; Meng, K.; Qian, C.; Zhang, Q.; et al. MicroRNA-101a Inhibits Cardiac Fibrosis Induced by Hypoxia via Targeting TGF beta RI on Cardiac Fibroblasts. Cell Physiol. Biochem. 2015, 35, 213–226. [Google Scholar] [CrossRef]
- Feng, Y.; Bao, Y.; Ding, J.; Li, H.; Liu, W.; Wang, X.; Guan, H.; Chen, Z. MicroRNA-130a attenuates cardiac fibrosis after myocardial infarction through TGF-beta/Smad signaling by directly targeting TGF-beta receptor 1. Bioengineered 2022, 13, 5779–5791. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Hou, L.; Zhao, J.; Wang, Y.; Jiang, F.; Jiang, Q.; Zhu, Z.; Tian, L. Exosomal let-7i-5p from three-dimensional cultured human umbilical cord mesenchymal stem cells inhibits fibroblast activation in silicosis through targeting TGFBR1. Ecotoxicol. Environ. Saf. 2022, 233, 113302. [Google Scholar] [CrossRef] [PubMed]
- Ukey, S.; Jain, A.; Dwivedi, S.; Choudhury, C.; Vishnoi, J.R.; Chugh, A.; Purohit, P.; Pareek, P.; Elhence, P.; Misra, S.; et al. Study of MicroRNA (miR-221-3p, miR-133a-3p, and miR-9-5p) Expressions in Oral Submucous Fibrosis and Squamous Cell Carcinoma. Indian J. Clin. Biochem. 2022. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, Y.; Liu, Y.; Li, X.; Tang, L.; Duan, M.; Li, J.; Zhang, G. Exosomes derived from human umbilical cord blood mesenchymal stem cells stimulate regenerative wound healing via transforming growth factor-beta receptor inhibition. Stem Cell Res. Ther. 2021, 12, 434. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Xu, Q.; Liu, Y.; Li, G.; Sun, W.; Ma, D.; Ni, C. Long noncoding RNA-SNHG20 promotes silica-induced pulmonary fibrosis by miR-490-3p/TGFBR1 axis. Toxicology 2021, 451, 152683. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wu, H.; Chen, S.; Qi, B.; Zhou, G.; Cai, L.; Zhao, L.; Wei, Y.; Liu, S. MicroRNA-30c suppresses the pro-fibrogenic effects of cardiac fibroblasts induced by TGF-beta 1 and prevents atrial fibrosis by targeting TGF beta RII. J. Cell Mol. Med. 2018, 22, 3045–3057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, P.; Sassi, Y.; Troncone, L.; Benard, L.; Ishikawa, K.; Gordon, R.E.; Lamas, S.; Laborda, J.; Hajjar, R.J.; Lebeche, D. Deletion of delta-like 1 homologue accelerates fibroblast-myofibroblast differentiation and induces myocardial fibrosis. Eur. Heart J. 2019, 40, 967–978. [Google Scholar] [CrossRef]
- Miguel, V.; Busnadiego, O.; Fierro-Fernandez, M.; Lamas, S. Protective role for miR-9-5p in the fibrogenic transformation of human dermal fibroblasts. Fibrogenesis Tissue Repair 2016, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Xiao, Z.; Guo, H.; Fang, X.; Liang, J.; Zhu, J.; Yang, J.; Li, H.; Pan, R.; Yuan, S.; et al. Novel role of the clustered miR-23b-3p and miR-27b-3p in enhanced expression of fibrosis-associated genes by targeting TGFBR3 in atrial fibroblasts. J. Cell Mol. Med. 2019, 23, 3246–3256. [Google Scholar] [CrossRef]
- Liang, H.; Zhang, C.; Ban, T.; Liu, Y.; Mei, L.; Piao, X.; Zhao, D.; Lu, Y.; Chu, W.; Yang, B. A novel reciprocal loop between microRNA-21 and TGF beta RIII is involved in cardiac fibrosis. Int. J. Biochem. Cell Biol. 2012, 44, 2152–2160. [Google Scholar] [CrossRef]
- Su, J.-L.; Chen, P.-S.; Johansson, G.; Kuo, M.-L. Function and regulation of let-7 family microRNAs. MicroRNA 2012, 1, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Banerjee, S.; Xie, N.; Ge, J.; Liu, R.-M.; Matalon, S.; Thannickal, V.J.; Liu, G. MicroRNA-27a-3p Is a Negative Regulator of Lung Fibrosis by Targeting Myofibroblast Differentiation. Am. J. Respir. Cell Mol. Biol. 2016, 54, 843–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, P.; Qiao, Q.; Ji, G.; Zhang, Z. CircAMD1 regulates proliferation and collagen synthesis via sponging miR-27a-3p in P63-mutant human dermal fibroblasts. Differentiation 2021, 119, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Wang, Y.-C.; Mei, X.; Shi, N.; Sun, C.; Ran, R.; Zhang, G.; Li, W.; Staveley-O’Carroll, K.F.; Li, G.; et al. LncRNA GAS5 attenuates fibroblast activation through inhibiting Smad3 signaling. Am. J. Physiol.-Cell Physiol. 2020, 319, C105–C115. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Zhang, Y.; Hu, Y.; Guan, J.; Xu, L.; Xiao, W.; Zhong, Q.; Ren, C.; Lu, J.; Liang, J.; et al. Long noncoding RNA Crnde attenuates cardiac fibrosis via Smad3-Crnde negative feedback in diabetic cardiomyopathy. FEBS J. 2019, 286, 1645–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Yang, K.; Chen, D.; Song, Y.; Qiao, W.; Sun, X.; Meng, L.; Bian, Z. Antifibrotic Potential of MiR-335-3p in Hereditary Gingival Fibromatosis. J. Dent. Res. 2019, 98, 1140–1149. [Google Scholar] [CrossRef]
- Schneider, C.; King, R.M.; Philipson, L. Genes specifically expressed at growth arrest of mammalian cells. Cell 1988, 54, 787–793. [Google Scholar] [CrossRef]
- Xiang, Z.; Liqing, Y.; Qingqing, Y.; Qiang, H.; Hongbo, C. Retard or exacerbate: Role of long non-coding RNA growth arrest-specific 5 in the fibrosis. Cytokine Growth Factor Rev. 2022, 67, 89–104. [Google Scholar] [CrossRef]
- She, Q.; Shi, P.; Xu, S.-S.; Xuan, H.-Y.; Tao, H.; Shi, K.-H.; Yang, Y. DNMT1 Methylation of LncRNA GAS5 Leads to Cardiac Fibroblast Pyroptosis via Affecting NLRP3 Axis. Inflammation 2020, 43, 1065–1076. [Google Scholar] [CrossRef]
- Wang, W.; Jia, Y.-J.; Yang, Y.-L.; Xue, M.; Zheng, Z.-J.; Wang, L.; Xue, Y.-M. LncRNA GAS5 exacerbates renal tubular epithelial fibrosis by acting as a competing endogenous RNA of miR-96-5p. Biomed. Pharmacother. 2020, 121, 109411. [Google Scholar] [CrossRef]
- Dong, Z.-G.; Yang, Y.-P.; Jiang, Z.-J. Knockdown CRNDE alleviates LPS-induced inflammation injury via FOXM1 in WI-38 cells. Biomed. Pharmacother. 2018, 103, 1678–1687. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, P.; Wang, C.; Tian, R. LncRNA CRNDE hinders the progression of osteoarthritis by epigenetic regulation of DACT1. Cell Mol. Life Sci. 2022, 79, 405. [Google Scholar] [CrossRef]
- Qi, Y.; Cui, S.; Liu, L.; Liu, B.; Wang, T.; Yan, S.; Tian, H.; Huang, X. Expression and role of miR-146a and SMAD4 in placental tissue of pregnant women with preeclampsia. J. Obstet. Gynaecol. Res. 2022, 48, 2151–2161. [Google Scholar] [CrossRef]
- Dong, M.; Wang, X.; Tong, L.; Wang, J.; Yang, Y.; Liu, Y.; Jing, Y.; Zhao, H.; Chen, J. Mir-27a-3p attenuates bronchiolitis obliterans in vivo via the regulation of dendritic cells’ maturation and the suppression of myofibroblasts’ differentiation. Clin. Transl. Med. 2020, 10, 140. [Google Scholar] [CrossRef] [PubMed]
- Nakasa, T.; Miyaki, S.; Okubo, A.; Hashimoto, M.; Nishida, K.; Ochi, M.; Asahara, H. Expression of MicroRNA-146 in rheumatoid arthritis synovial tissue. Arthritis Rheum. 2008, 58, 1284–1292. [Google Scholar] [CrossRef] [Green Version]
- Yogeswaran, A.; Troidl, C.; McNamara, J.W.; Wilhelm, J.; Truschel, T.; Widmann, L.; Aslam, M.; Hamm, C.W.; Sadayappan, S.; Lipps, C. The C0-C1f Region of Cardiac Myosin Binding Protein-C Induces Pro-Inflammatory Responses in Fibroblasts via TLR4 Signaling. Cells 2021, 10, 1326. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Morelli, M.B.; Matarese, A.; Sardu, C.; Santulli, G. Cardiomyocyte-derived exosomal microRNA-92a mediates post-ischemic myofibroblast activation both in vitro and ex vivo. ESC Heart Fail. 2020, 7, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Zhang, M.; Ding, X.; Yang, X.; Zheng, D.; Mou, H.; Ruan, X.; Feng, Y.; Han, H.; Wang, S. MicroRNA-21a-5p regulates TGF-beta 1-induced myofibroblast transformation of rabbit keratocytes by targeting Smad7. Scienceasia 2022, 48, 57–61. [Google Scholar] [CrossRef]
- Wang, C.; Gu, S.; Cao, H.; Li, Z.; Xiang, Z.; Hu, K.; Han, X. miR-877-3p targets Smad7 and is associated with myofibroblast differentiation and bleomycin-induced lung fibrosis. Sci. Rep. 2016, 6, 30122. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wang, Y.; Deng, Y.; Ni, H.; Shen, G.; Liu, X.; Li, J.; Wang, F. Epigenetic Control of circHNRNPH1 in Postischemic Myocardial Fibrosis through Targeting of TGF-beta Receptor Type I. Mol. Ther.-Nucleic Acids 2021, 25, 93–104. [Google Scholar] [CrossRef]
- Wang, J.; Hu, K.; Cai, X.; Yang, B.; He, Q.; Wang, J.; Weng, Q. Targeting PI3K/AKT signaling for treatment of idiopathic pulmonary fibrosis. Acta Pharm. Sin. B 2022, 12, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; You, J.; Gong, D.; Xu, Y.; Yang, B.; Jiang, C. PDGF-BB induces conversion, proliferation, migration, and collagen synthesis of oral mucosal fibroblasts through PDGFR-beta/PI3K/ AKT signaling pathway. Cancer Biomark. 2021, 30, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Kalantari, P.; Harandi, O.F.; Agarwal, S.; Rus, F.; Kurt-Jones, E.A.; Fitzgerald, K.A.; Caffrey, D.R.; Golenbock, D.T. miR-718 represses proinflammatory cytokine production through targeting phosphatase and tensin homolog (PTEN). J. Biol. Chem. 2017, 292, 5634–5644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Liang, Y.; Lin, Q.; Liu, J.; Luo, F.; Li, X.; Zhou, H.; Zhuang, S.; Zhang, H. MiR-29 mediates TGF1-induced extracellular matrix synthesis through activation of PI3K-AKT pathway in human lung fibroblasts. J. Cell Biochem. 2013, 114, 1336–1342. [Google Scholar] [CrossRef]
- Li, H.; Tian, X.; Ruan, Y.; Xing, J.; Meng, Z. Asiatic acid alleviates Ang-II induced cardiac hypertrophy and fibrosis via miR-126/PIK3R2 signaling. Nutr. Metab. 2021, 18, 71. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Li, W.; Yu, W.; Rao, T.; Li, H.; Ruan, Y.; Yuan, R.; Li, C.; Ning, J.; Li, S.; et al. Exosomal miR-21 from tubular cells contributes to renal fibrosis by activating fibroblasts via targeting PTEN in obstructed kidneys. Theranostics 2021, 11, 8660–8673. [Google Scholar] [CrossRef]
- Rackow, A.R.; Judge, J.L.; Woeller, C.F.; Sime, P.J.; Kottmann, R.M. miR-338-3p blocks TGF8-induced myofibroblast differentiation through the induction of PTEN. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2022, 322, L385–L400. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Han, L.; Gui, W.; Wang, F.; Yan, W.; Jiang, H. MiR-503 suppresses fibroblast activation and myofibroblast differentiation by targeting VEGFA and FGFR1 in silica-induced pulmonary fibrosis. J. Cell Mol. Med. 2020, 24, 14339–14348. [Google Scholar] [CrossRef]
- Huang, Y.; Xie, Y.; Abel, P.W.; Wei, P.; Plowman, J.; Toews, M.L.; Strah, H.; Siddique, A.; Bailey, K.L.; Tu, Y. TGF-beta 1-induced miR-424 promotes pulmonary myofibroblast differentiation by targeting Slit2 protein expression. Biochem. Pharmacol. 2020, 180, 114172. [Google Scholar] [CrossRef]
- Huang, C.; Yang, Y.; Liu, L. Interaction of long noncoding RNAs and microRNAs in the pathogenesis of idiopathic pulmonary fibrosis. Physiol. Genom. 2015, 47, 463–469. [Google Scholar] [CrossRef]
- Li, X.; Yu, T.; Shan, H.; Jiang, H.; Sun, J.; Zhao, X.; Su, W.; Yang, L.; Shan, H.; Liang, H. lncRNA PFAL promotes lung fibrosis through CTGF by competitively binding miR-18a. FASEB J. 2018, 32, 5285–5297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.; Xie, Y.; Yuan, Y.; Hu, K. LncRNA GAS5 as miR-26a-5p Sponge Regulates the PTEN/PI3K/Akt Axis and Affects Extracellular Matrix Synthesis in Degenerative Nucleus Pulposus Cells in vitro. Front. Neurol. 2021, 12, 653341. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-Y.; Li, C.; Bai, W.-D.; Su, L.-L.; Liu, J.-Q.; Li, Y.; Shi, J.-H.; Cai, W.-X.; Bai, X.-Z.; Jia, Y.-H.; et al. MicroRNA-21 Regulates hTERT via PTEN in Hypertrophic Scar Fibroblasts. PLoS ONE 2014, 9, E0097114. [Google Scholar] [CrossRef]
- Fan, Y.; Zhao, X.; Ma, J.; Yang, L. LncRNA GAS5 Competitively Combined With miR-21 Regulates PTEN and Influences EMT of Peritoneal Mesothelial Cells via Wnt/beta-Catenin Signaling Pathway. Front. Physiol. 2021, 12, 654951. [Google Scholar] [CrossRef]
- Song, M.; Zhao, G.; Sun, H.; Yao, S.; Zhou, Z.; Jiang, P.; Wu, Q.; Zhu, H.; Wang, H.; Dai, C.; et al. circPTPN12/miR-21-5 p/Delta Np63 alpha pathway contributes to human endometrial fibrosis. Elife 2021, 10, 65735. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Guo, H.; Qian, J.; Liu, W.; Li, Y.; Yuan, L.; Zhou, Y.; Lin, R.; Xie, X.; Yang, Q.; et al. Effects of bone morphogenetic protein 4 on TGF-beta 1-induced cell proliferation, apoptosis, activation and differentiation in mouse lung fibroblasts via ERK/p38 MAPK signaling pathway. PeerJ 2022, 10, 13775. [Google Scholar] [CrossRef]
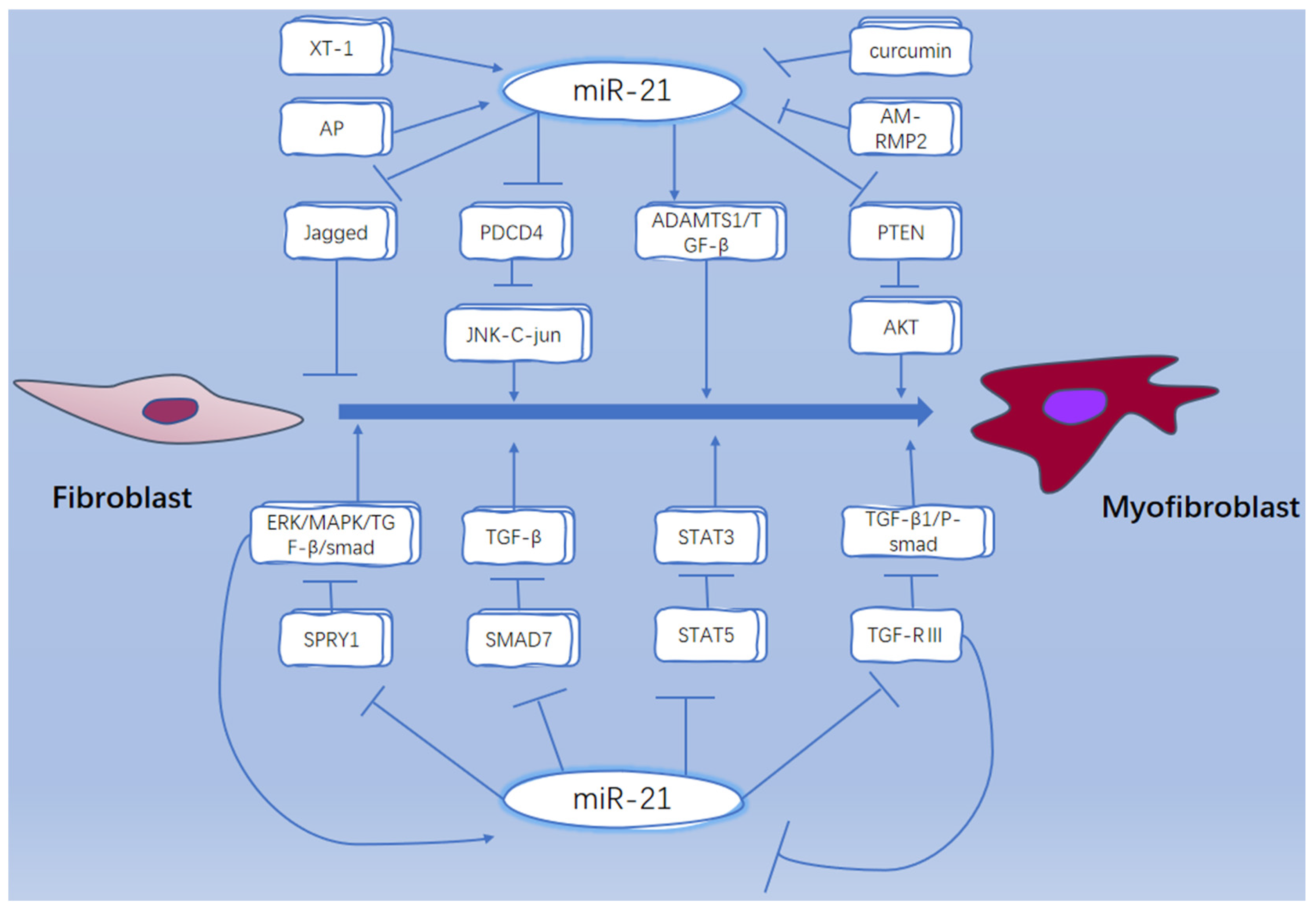
- Sun, Q.; Miao, J.; Luo, J.; Yuan, Q.; Cao, H.; Su, W.; Zhou, Y.; Jiang, L.; Fang, L.; Dai, C.; et al. The feedback loop between miR-21, PDCD4 and AP-1 functions as a driving force for renal fibrogenesis. J. Cell Sci. 2018, 131, jcs202317. [Google Scholar] [CrossRef] [Green Version]
- Auler, M.; Bergmeier, V.; Georgieva, V.S.; Pitzler, L.; Frie, C.; Nuechel, J.; Eckes, B.; Hinz, B.; Brachvogel, B. miR-127-3p Is an Epigenetic Activator of Myofibroblast Senescence Situated within the MicroRNA-Enriched Dlk1-Dio3-Imprinted Domain on Mouse Chromosome 12. J. Investig. Dermatol. 2021, 141, 1076–1086. [Google Scholar] [CrossRef]
- Shen, J.; Xing, W.; Liu, R.; Zhang, Y.; Xie, C.; Gong, F. MiR-32-5p influences high glucose-induced cardiac fibroblast proliferation and phenotypic alteration by inhibiting DUSP1. BMC Mol. Biol. 2019, 20, 21. [Google Scholar] [CrossRef]
- Huang, C.; Wang, R.; Lu, J.; He, Y.; Wu, Y.; Ma, W.; Xu, J.; Wu, Z.; Feng, Z.; Wu, M. MicroRNA-338-3p as a therapeutic target in cardiac fibrosis through FGFR2 suppression. J. Clin. Lab. Anal. 2022, 36, e24584. [Google Scholar] [CrossRef]
- Zheng, L.; Xu, C.-C.; Chen, W.-D.; Shen, W.-L.; Ruan, C.-C.; Zhu, L.-M.; Zhu, D.-L.; Gao, P.-J. MicroRNA-155 regulates angiotensin II type 1 receptor expression and phenotypic differentiation in vascular adventitial fibroblasts. Biochem. Biophys. Res. Commun. 2010, 400, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Kuse, N.; Kamio, K.; Azuma, A.; Matsuda, K.; Inomata, M.; Usuki, J.; Morinaga, A.; Tanaka, T.; Kashiwada, T.; Atsumi, K.; et al. Exosome-Derived microRNA-22 Ameliorates Pulmonary Fibrosis by Regulating Fibroblast-to-Myofibroblast Differentiation in Vitro and in Vivo. J. Nippon. Med. Sch. 2020, 87, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, G.; Jin, X.; Zhang, L. LncRNA FENDRR Upregulation Promotes Hepatic Carcinoma Cells Apoptosis By Targeting miR-362-5p Via NPR3 and p38-MAPK Pathway. Cancer Biother. Radiopharm. 2020, 35, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Sun, B.; Wang, Z.; Yang, M.; Cui, Z.; Lin, S.; Jin, M.; Yi, C. Exosomes From M2 Macrophage Promote Peritendinous Fibrosis Posterior Tendon Injury via the MiR-15b-5p/FGF-1/7/9 Pathway by Delivery of circRNA-Ep400. Front. Cell Dev. Biol. 2021, 9, 1557. [Google Scholar] [CrossRef]
- Liao, Y.-W.; Tsai, L.-L.; Lee, Y.-H.; Hsieh, P.-L.; Yu, C.-C.; Lu, M.-Y. miR-21 promotes the fibrotic properties in oral mucosa through targeting PDCD4. J. Dent. Sci. 2022, 17, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Mao, C.; Zhou, E.; You, J.; Gao, E.; Han, Z.; Fan, Y.; He, Q.; Wang, C. MicroRNA-21 Mediates a Positive Feedback on Angiotensin II-Induced Myofibroblast Transformation. J. Inflamm. Res. 2020, 13, 1007–1020. [Google Scholar] [CrossRef]
- Yan, J.; Yang, Y.; Fan, X.; Liang, G.; Wang, Z.; Li, J.; Wang, L.; Chen, Y.; Adetula, A.A.; Tang, Y.; et al. circRNAome profiling reveals circFgfr2 regulates myogenesis and muscle regeneration via a feedback loop. J. Cachexia Sarcopenia Muscle 2022, 13, 696–712. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.; Deng, R.; Gao, S.; Jiang, Q.; Liu, R.; Li, H.; Miao, Y.; Zhai, Y.; Zhang, S.; et al. Betulinic acid attenuated bleomycin-induced pulmonary fibrosis by effectively intervening Wnt/beta-catenin signaling. Phytomedicine 2021, 81, 153428. [Google Scholar] [CrossRef]
- Ortiz-Quintero, B.; Buendia-Roldan, I.; Gustavo Ramirez-Salazar, E.; Balderas-Martinez, Y.I.; Lizbeth Ramirez-Rodriguez, S.; Martinez-Espinosa, K.; Selman, M. Circulating microRNA Signature Associated to Interstitial Lung Abnormalities in Respiratory Asymptomatic Subjects. Cells 2020, 9, 1556. [Google Scholar] [CrossRef]
- Henderson, J.; Pryzborski, S.; Stratton, R.; O’Reilly, S. Wnt antagonist DKK-1 levels in systemic sclerosis are lower in skin but not in blood and are regulated by microRNA33a-3p. Exp. Dermatol. 2021, 30, 162–168. [Google Scholar] [CrossRef]
- Sun, L.-Y.; Bie, Z.-D.; Zhang, C.-H.; Li, H.; Li, L.-D.; Yang, J. MiR-154 directly suppresses DKK2 to activate Wnt signaling pathway and enhance activation of cardiac fibroblasts. Cell Biol. Int. 2016, 40, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Huang, X.; Luo, S.; Zhang, H.; Hu, F.; Chen, R.; Huang, C.; Su, Z. The MicroRNA MiR-29c Alleviates Renal Fibrosis via TPM1-Mediated Suppression of the Wnt/beta-Catenin Pathway. Front. Physiol. 2020, 11, 331. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.-j.; Qian, H.; Hu, K.; Zhang, Z.-z.; He, J.-b. miR-27a-3p inhibited synthesis of Col I and Col III in pulmonary fibroblasts through Wnt3a/beta-eatenin signaling pathway. Chin. Pharmacol. Bull. 2019, 35, 229–234. [Google Scholar] [CrossRef]
- Liu, L.; Qian, H.; Hu, K.; Wang, L.; Zhang, Z.; Yin, H.; He, J. miR-27a-3p inhibits pulmonary fibrosis by blocking Wnt3a/beta-catenin pathway in rats. Chin. J. Cell. Mol. Immunol. 2018, 34, 1015–1020. [Google Scholar]
- Hao, K.; Lei, W.; Wu, H.; Wu, J.; Yang, Z.; Yan, S.; Lu, X.-A.; Li, J.; Xia, X.; Han, X.; et al. LncRNA-Safe contributes to cardiac fibrosis through Safe-Sfrp2-HuR complex in mouse myocardial infarction. Theranostics 2019, 9, 7282–7297. [Google Scholar] [CrossRef]
- Papaioannou, I.; Xu, S.; Denton, C.P.; Abraham, D.J.; Ponticos, M. STAT3 controls COL1A2 enhancer activation cooperatively with JunB, regulates type I collagen synthesis posttranscriptionally, and is essential for lung myofibroblast differentiation. Mol. Biol. Cell. 2018, 29, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Qiu, Y. MiR-210-5p regulates STAT3 activation by targeting STAT5A in the differentiation of dermal fibroblasts. 3 Biotech 2021, 11, 243. [Google Scholar] [CrossRef]
- Yue, Z.; Jiang, Z.; Ruan, B.; Duan, J.; Song, P.; Liu, J.; Han, H.; Wang, L. Disruption of myofibroblastic Notch signaling attenuates liver fibrosis by modulating fibrosis progression and regression. Int. J. Biol. Sci. 2021, 17, 2135–2146. [Google Scholar] [CrossRef]
- Yao, Q.; Xing, Y.; Wang, Z.; Liang, J.; Lin, Q.; Huang, M.; Chen, Y.; Lin, B.; Xu, X.; Chen, W. MiR-16-5p suppresses myofibroblast activation in systemic sclerosis by inhibiting NOTCH signaling. Aging 2021, 13, 2640–2654. [Google Scholar] [CrossRef]
- Zhao, S.; Xiao, X.; Sun, S.; Li, D.; Wang, W.; Fu, Y.; Fan, F. MicroRNA-30d/JAG1 axis modulates pulmonary fibrosis through Notch signaling pathway. Pathol. Res. Pract. 2018, 214, 1315–1323. [Google Scholar] [CrossRef]
- Zhou, X.-L.; Xu, H.; Liu, Z.-B.; Wu, Q.-C.; Zhu, R.-R.; Liu, J.-C. miR-21 promotes cardiac fibroblast-to-myofibroblast transformation and myocardial fibrosis by targeting Jagged1. J. Cell Mol. Med. 2018, 22, 3816–3824. [Google Scholar] [CrossRef]
- Wasson, C.W.; Ross, R.L.; Wells, R.; Corinaldesi, C.; Georgiou, I.C.; Riobo-Del Galdo, N.A.; Del Galdo, F. Long non-coding RNA HOTAIR induces GLI2 expression through Notch signalling in systemic sclerosis dermal fibroblasts. Arthritis Res. Ther. 2020, 22, 286. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.-W.; Yu, C.-C.; Hsieh, P.-L.; Chang, Y.-C. miR-200b ameliorates myofibroblast transdifferentiation in precancerous oral submucous fibrosis through targeting ZEB2. J. Cell Mol. Med. 2018, 22, 4130–4138. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.-Y.; Yu, C.-C.; Chen, P.-Y.; Hsieh, P.-L.; Peng, C.-Y.; Liao, Y.-W.; Yu, C.-H.; Lin, K.-H. miR-200c inhibits the arecoline-associated myofibroblastic transdifferentiation in buccal mucosal fibroblasts. J. Formos. Med. Assoc. 2018, 117, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Feng, L.; Li, Z.; Xu, G.; Fan, X. MicroRNA-21 activates hepatic stellate cells via PTEN/Akt signaling. Biomed. Pharmacother. 2013, 67, 387–392. [Google Scholar] [CrossRef]
- Gallant-Behm, C.L.; Piper, J.; Lynch, J.M.; Seto, A.G.; Hong, S.J.; Mustoe, T.A.; Maari, C.; Pestano, L.A.; Dalby, C.M.; Jackson, A.L.; et al. A MicroRNA-29 Mimic (Remlarsen) Represses Extracellular Matrix Expression and Fibroplasia in the Skin. J. Investig. Dermatol. 2019, 139, 1073–1081. [Google Scholar] [CrossRef]
- Dai, C.; Tao, H.; Shi, K.-H.; Xu, S.-S. Effect of miR-369-5p on cardiac fibroblast activation and proliferation in SD rats. Chin. Pharmacol. Bull. 2018, 34, 1693–1696. [Google Scholar] [CrossRef]
- Schnittert, J.; Kuninty, P.R.; Bystry, T.F.; Brock, R.; Storm, G.; Prakash, J. Anti-microRNA targeting using peptide-based nanocomplexes to inhibit differentiation of human pancreatic stellate cells. Nanomedicine 2017, 12, 1369–1384. [Google Scholar] [CrossRef]
- Szczerba, E.; Zajkowska, A.; Bochowicz, A.; Pankiewicz, K.; Szewczyk, G.; Opolski, G.; Maciejewski, T.; Malecki, M.; Fijalkowska, A. Downregulated expression of microRNAs associated with cardiac hypertrophy and fibrosis in physiological pregnancy and the association with echocardiographically-evaluated myocardial function. Biomed. Rep. 2020, 13, 41. [Google Scholar] [CrossRef]
- Zanotti, S.; Gibertini, S.; Blasevich, F.; Bragato, C.; Ruggieri, A.; Saredi, S.; Fabbri, M.; Bernasconi, P.; Maggi, L.; Mantegazza, R.; et al. Exosomes and exosomal miRNAs from muscle-derived fibroblasts promote skeletal muscle fibrosis. Matrix Biol. 2018, 74, 77–100. [Google Scholar] [CrossRef]
- Yi, M.; Liu, B.; Tang, Y.; Li, F.; Qin, W.; Yuan, X. Irradiated Human Umbilical Vein Endothelial Cells Undergo Endothelial-Mesenchymal Transition via the Snail/miR-199a-5p Axis to Promote the Differentiation of Fibroblasts into Myofibroblasts. Biomed. Res. Int. 2018, 2018, 4135806. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, C.L.L.; Henaoui, I.S.; Courcot, E.; Roderburg, C.; Cauffiez, C.; Aubert, S.; Copin, M.-C.; Wallaert, B.; Glowacki, F.; Dewaeles, E.; et al. miR-199a-5p Is Upregulated during Fibrogenic Response to Tissue Injury and Mediates TGFbeta-Induced Lung Fibroblast Activation by Targeting Caveolin-1. PLos Genet. 2013, 9, 1003291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, P.-Y.; Ho, D.C.-Y.; Chen, S.-H.; Hsieh, P.-L.; Liao, Y.-W.; Tsai, L.-L.; Yu, C.-C.; Fang, C.-Y. Down-regulation of miR-29c promotes the progression of oral submucous fibrosis through targeting tropomyosin-1. J. Formos. Med. Assoc. 2022, 121, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Chen, X.; Sun, J.; An, G.; Li, X.; Wang, Y.; Niu, P.; Zhu, Z.; Tian, L. MicroRNA-29b inhibits supernatants from silica-treated macrophages from inducing extracellular matrix synthesis in lung fibroblasts. Toxicol. Res. 2017, 6, 878–888. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Kong, M. Tetrahydroxy stilbene glucoside alleviates palmitic acid-induced inflammation and apoptosis in cardiomyocytes by regulating miR-129-3p/Smad3 signaling. Cell Mol. Biol. Lett. 2019, 24, 5. [Google Scholar] [CrossRef]
- Wang, F.; Zhao, X.-Q.; Liu, J.-N.; Wang, Z.-H.; Wang, X.-L.; Hou, X.-Y.; Liu, R.; Gao, F.; Zhang, M.-X.; Zhang, Y.; et al. Antagonist of microRNA-21 improves balloon injury-induced rat iliac artery remodeling by regulating proliferation and apoptosis of adventitial fibroblasts and myofibroblasts. J. Cell Biochem. 2012, 113, 2989–3001. [Google Scholar] [CrossRef]
- Yang, H.-W.; Yu, C.-C.; Hsieh, P.-L.; Liao, Y.-W.; Chu, P.-M.; Yu, C.-H.; Fang, C.-Y. Arecoline enhances miR-21 to promote buccal mucosal fibroblasts activation. J. Formos. Med. Assoc. 2021, 120, 1108–1113. [Google Scholar] [CrossRef]
- Singh, P.; Srivastava, A.N.; Sharma, R.; Mateen, S.; Shukla, B.; Singh, A.; Chandel, S. Circulating MicroRNA-21 Expression as a Novel Serum Biomarker for Oral Sub-Mucous Fibrosis and Oral Squamous Cell Carcinoma. Asian Pac. J. Cancer Prev. 2018, 19, 1053–1057. [Google Scholar]
- Wang, J.; Guo, R.; Ma, X.; Wang, Y.; Zhang, Q.; Zheng, N.; Zhang, J.; Li, C. Liraglutide inhibits AngII-induced cardiac fibroblast proliferation and ECM deposition through regulating miR-21/PTEN/PI3K pathway. Cell Tissue Bank. 2022. [Google Scholar] [CrossRef]
- Schipper, J.; Westerhuis, J.J.; Beddows, I.; Madaj, Z.; Monsma, D.; Hostetter, G.; Kiupel, M.; Conejo-Garcia, J.R.; Sempere, L.F. Loss of microRNA-21 leads to profound stromal remodeling and short survival in K-Ras-driven mouse models of pancreatic cancer. Int. J. Cancer 2020, 147, 2265–2278. [Google Scholar] [CrossRef]
- Li, N.; Wang, Z.; Gao, F.; Lei, Y.; Li, Z. Melatonin ameliorates renal fibroblast-myofibroblast transdifferentiation and renal fibrosis through miR-21-5p regulation. J. Cell Mol. Med. 2020, 24, 5615–5628. [Google Scholar] [CrossRef] [Green Version]
- Yao, Q.; Cao, S.; Li, C.; Mengesha, A.; Kong, B.; Wei, M. Micro-RNA-21 regulates TGF-beta-induced myofibroblast differentiation by targeting PDCD4 in tumor-stroma interaction. Int. J. Cancer 2011, 128, 1783–1792. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Nie, Y.; Qu, S.; Liao, J.-Y.; Cui, X.; Yao, H.; Zeng, Y.; Su, F.; Song, E.; Liu, Q. miR-21 Induces Myofibroblast Differentiation and Promotes the Malignant Progression of Breast Phyllodes Tumors. Cancer Res. 2014, 74, 4341–4352. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Friggeri, A.; Yang, Y.; Milosevic, J.; Ding, Q.; Thannickal, V.J.; Kaminski, N.; Abraham, E. miR-21 mediates fibrogenic activation of pulmonary fibroblasts and lung fibrosis. J. Exp. Med. 2010, 207, 1589–1597. [Google Scholar] [CrossRef]
- Jiang, C.; Guo, Y.; Yu, H.; Lu, S.; Meng, L. Pleiotropic microRNA-21 in pulmonary remodeling: Novel insights for molecular mechanism and present advancements. Allergy Asthma Clin. Immunol. 2019, 15, 33. [Google Scholar] [CrossRef] [Green Version]
- Kilari, S.; Cai, C.; Zhao, C.; Sharma, A.; Chernogubova, E.; Simeon, M.; Wu, C.-C.; Song, H.-L.; Maegdefessel, L.; Misra, S. The Role of MicroRNA-21 in Venous Neointimal Hyperplasia: Implications for Targeting miR-21 for VNH Treatment. Mol. Ther. 2019, 27, 1681–1693. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-C.; Tseng, Y.-K.; Chi, C.-C.; Chen, Y.-H.; Yang, C.-M.; Huang, S.-J.; Lee, Y.-C.; Liou, H.-H.; Tsai, K.-W.; Ger, L.-P. Genetic variants in microRNA-146a (C > G) and microRNA-1269b (G > C) are associated with the decreased risk of oral premalignant lesions, oral cancer, and pharyngeal cancer. Arch. Oral Biol. 2016, 72, 21–32. [Google Scholar] [CrossRef]
- Yang, J.; Yu, X.; Xue, F.; Li, Y.; Liu, W.; Zhang, S. Exosomes derived from cardiomyocytes promote cardiac fibrosis via myocyte-fibroblast cross-talk. Am. J. Transl. Res. 2018, 10, 4350–4366. [Google Scholar]
- Al-Harbi, B.; Hendrayani, S.-F.; Silva, G.; Aboussekhra, A. Let-7b inhibits cancer-promoting effects of breast cancer-associated fibroblasts through IL-8 repression. Oncotarget. 2018, 9, 17825–17838. [Google Scholar] [CrossRef]
- Huleihel, L.; Ben-Yehudah, A.; Milosevic, J.; Yu, G.; Pandit, K.; Sakamoto, K.; Yousef, H.; LeJeune, M.; Coon, T.A.; Redinger, C.J.; et al. Let-7d microRNA affects mesenchymal phenotypic properties of lung fibroblasts. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2014, 306, L534–L542. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Jia, X.; Zhang, Q.; Zhang, L.; Yang, J.; Hu, C.; Shi, J.; Jiang, X.; Lu, J.; Shen, H. Neutrophil extracellular traps activate lung fibroblast to induce polymyositis-related interstitial lung diseases via TLR9-miR-7-Smad2 pathway. J. Cell Mol. Med. 2020, 24, 1658–1669. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Shi, J.; Hu, Z.; Hu, X. The deficiency of miR-214-3p exacerbates cardiac fibrosis via miR-214-3p/NLRC5 axis. Clin. Sci. 2019, 133, 1845–1856. [Google Scholar] [CrossRef]
- Dong, H.; Dong, S.; Zhang, L.; Gao, X.; Lv, G.; Chen, W.; Shao, S. MicroRNA-214 exerts a Cardio-protective effect by inhibition of fibrosis. Anat. Rec.-Adv. Integr. Anat. Evol. Biol. 2016, 299, 1348–1357. [Google Scholar] [CrossRef]
- Sun, M.; Yu, H.; Zhang, Y.; Li, Z.; Gao, W. MicroRNA-214 Mediates Isoproterenol-induced Proliferation and Collagen Synthesis in Cardiac Fibroblasts. Sci. Rep. 2015, 5, 18351. [Google Scholar] [CrossRef] [Green Version]
- Conde, E.; Gimenez-Moyano, S.; Martin-Gomez, L.; Rodriguez, M.; Edurne Ramos, M.; Aguado-Fraile, E.; Blanco-Sanchez, I.; Saiz, A.; Laura Garcia-Bermejo, M. HIF-1 alpha induction during reperfusion avoids maladaptive repair after renal ischemia/reperfusion involving miR127-3p. Sci. Rep. 2017, 7, 41099. [Google Scholar] [CrossRef] [Green Version]
- Hogan, T.B.; Tiwari, N.; Nagaraja, M.R.; Shetty, S.K.; Fan, L.; Shetty, R.S.; Bhandary, Y.P.; Shetty, S. Caveolin-1 peptide regulates p53-microRNA-34a feedback in fibrotic lung fibroblasts. Iscience 2022, 25, 104022. [Google Scholar] [CrossRef]
- Bulvik, R.; Biton, M.; Berkman, N.; Breuer, R.; Wallach-Dayan, S.B. Forefront: MiR-34a-Knockout Mice with Wild Type Hematopoietic Cells, Retain Persistent Fibrosis Following Lung Injury. Int. J. Mol. Sci. 2020, 21, 2228. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhang, Y.; Zhu, H.; Hu, J.; Xie, Z. MiR-34a/miR-93 target c-Ski to modulate the proliferaton of rat cardiac fibroblasts and extracellular matrix deposition in vivo and in vitro. Cell Signalling. 2018, 46, 145–153. [Google Scholar] [CrossRef]
- Kanno, Y.; Shu, E.; Niwa, H.; Seishima, M.; Ozaki, K.-i. MicroRNA-30c attenuates fibrosis progression and vascular dysfunction in systemic sclerosis model mice. Mol. Biol. Rep. 2021, 48, 3431–3437. [Google Scholar] [CrossRef]
- Bajor, A.; Ratuszny, D.; Gras, C.; Pielen, A.; Boergel, M.; Framme, C.; Blasczyk, R.; Figueiredo, C. Inhibition of miR-145 as a strategy to prevent corneal scarring. Investig. Ophthalmol. Visual Sci. 2015, 56, 4917. [Google Scholar]
- Song, H.-F.; He, S.; Li, S.-H.; Wu, J.; Yin, W.; Shao, Z.; Du, G.-q.; Wu, J.; Li, J.; Weisel, R.D.; et al. Knock-out of MicroRNA 145 impairs cardiac fibroblast function and wound healing post-myocardial infarction. J. Cell Mol. Med. 2020, 24, 9409–9419. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Yasuno, K.; Tagawa, H.; Sakamoto, N.; Oue, N.; Yashiro, M.; Sentani, K.; Goto, K.; Shinmei, S.; Oo, H.Z.; et al. MicroRNA-145 is a potential prognostic factor of scirrhous type gastric cancer. Oncol. Rep. 2014, 32, 1720–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, T.-D.; Riedel, L.; Fischer, B.; Schmidt, V.; Hendig, D.; Distler, J.; Kuhn, J.; Knabbe, C.; Faust, I. microRNA-145 mediates xylosyltransferase-I induction in myofibroblasts via suppression of transcription factor KLF4. Biochem. Biophys. Res. Commun. 2020, 523, 1001–1006. [Google Scholar] [CrossRef] [PubMed]
- Gras, C.; Ratuszny, D.; Hadamitzky, C.; Zhang, H.; Blasczyk, R.; Figueiredo, C. miR-145 Contributes to Hypertrophic Scarring of the Skin by Inducing Myofibroblast Activity. Mol. Med. 2015, 21, 296–304. [Google Scholar] [CrossRef]
- Ratuszny, D.; Gras, C.; Bajor, A.; Boerger, A.-K.; Pielen, A.; Boergel, M.; Framme, C.; Blasczyk, R.; Figueiredo, C. miR-145 Is a Promising Therapeutic Target to Prevent Cornea Scarring. Hum. Gene Ther. 2015, 26, 698–707. [Google Scholar] [CrossRef]
- Yang, S.; Cui, H.; Xie, N.; Icyuz, M.; Banerjee, S.; Antony, V.B.; Abraham, E.; Thannickal, V.J.; Liu, G. miR-145 regulates myofibroblast differentiation and lung fibrosis. FASEB J. 2013, 27, 2382–2391. [Google Scholar] [CrossRef] [Green Version]
- Melling, G.E.; Flannery, S.E.; Abidin, S.A.; Clemmens, H.; Prajapati, P.; Hinsley, E.E.; Hunt, S.; Catto, J.W.F.; Della Coletta, R.; Mellone, M.; et al. A miRNA-145/TGF-beta 1 negative feedback loop regulates the cancer-associated fibroblast phenotype. Carcinogenesis 2018, 39, 798–807. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-S.; Li, S.-H.; Guo, J.; Mihic, A.; Wu, J.; Sun, L.; Davis, K.; Weisel, R.D.; Li, R.-K. Role of miR-145 in cardiac myofibroblast differentiation. J. Mol. Cell. Cardiol. 2014, 66, 94–105. [Google Scholar] [CrossRef]
- Liakouli, V.; Cipriani, P.; Di Benedetto, P.; Panzera, N.; Ruscitti, P.; Pantano, I.; Berardicurti, O.; Carubbi, F.; Esteves, F.; Mavria, G.; et al. Epidermal Growth Factor Like-domain 7 and miR-126 are abnormally expressed in diffuse Systemic Sclerosis fibroblasts. Sci. Rep. 2019, 9, 4589. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yin, H.; Jiao, L.; Liu, T.; Gao, Y.; Shao, Y.; Zhang, Y.; Shan, H.; Zhang, Y.; Yang, B. Abnormal Downregulation of Caveolin-3 Mediates the Pro-Fibrotic Action of MicroRNA-22 in a Model of Myocardial Infarction. Cell Physiol. Biochem. 2018, 45, 1641–1653. [Google Scholar] [CrossRef]
- Li, J.; Yue, S.; Fang, J.; Zeng, J.; Chen, S.; Tian, J.; Nie, S.; Liu, X.; Ding, H. MicroRNA-10a/b inhibit TGF-beta/Smad-induced renal fibrosis by targeting TGF-beta receptor 1 in diabetic kidney disease. Mol. Ther.-Nucleic Acids 2022, 28, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.-Y.; Yu, C.-C.; Liao, Y.-W.; Hsieh, P.L.; Ohiro, Y.; Chu, P.M.; Huang, Y.-C.; Yu, C.-H.; Tsai, L.-L. miR-10b regulated by Twist maintains myofibroblasts activities in oral submucous fibrosis. J. Formos. Med. Assoc. 2020, 119, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Chugh, A.; Purohit, P.; Vishnoi, J.R.; Kaur, A.; Modi, A.; Mishra, S.; Sharma, P.; Rodha, M.S.; Pareek, P.; Bhattacharya, S.; et al. Correlation of hsa miR-101-5p and hsa miR-155-3p Expression With c-Fos in Patients of Oral Submucous Fibrosis (OSMF) and Oral Squamous Cell Carcinoma (OSCC). J. Maxillofac. Oral Surg. 2021. [Google Scholar] [CrossRef]
- Chou, M.-Y.; Fang, C.-Y.; Hsieh, P.-L.; Liao, Y.-W.; Yu, C.-C.; Lee, S.-S. Depletion of miR-155 hinders the myofibroblast activities and reactive oxygen species generation in oral submucous fibrosis. J. Formos. Med. Assoc. 2022, 121, 467–472. [Google Scholar] [CrossRef]
- Artlett, C.M.; Sassi-Gaha, S.; Hope, J.L.; Feghali-Bostwick, C.A.; Katsikis, P.D. Mir-155 is overexpressed in systemic sclerosis fibroblasts and is required for NLRP3 inflammasome-mediated collagen synthesis during fibrosis. Arthritis Res. Ther. 2017, 19, 144. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Huang, H.; Xie, Q.; Wang, Z.; Fan, Y.; Kong, B.; Huang, D.; Xiao, Y. MiR-155 Knockout in Fibroblasts Improves Cardiac Remodeling by Targeting Tumor Protein p53-Inducible Nuclear Protein 1. J. Cardiovasc. Pharmacol. Ther. 2016, 21, 423–435. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Abak, A.; Talebi, S.F.; Shoorei, H.; Branicki, W.; Taheri, M.; Dilmaghani, N.A. Role of miRNA and lncRNAs in organ fibrosis and aging. Biomed. Pharmacother. 2021, 143, 112132. [Google Scholar] [CrossRef]
- Shen, S.; Wang, X.; Xu, D.; Tao, L.; Wu, X.; Xiao, J.; Li, X. Mir-195-3p/-5p Decrease Cardiac Fibroblast Proliferation and the Transdifferentiation into Myofibroblasts. Circ. Res. 2015, 117, 1580982. [Google Scholar] [CrossRef]
- You, K.; Li, S.-Y.; Gong, J.; Fang, J.-H.; Zhang, C.; Zhang, M.; Yuan, Y.; Yang, J.; Zhuang, S.-M. MicroRNA-125b Promotes Hepatic Stellate Cell Activation and Liver Fibrosis by Activating RhoA Signaling. Mol. Ther.-Nucleic Acids 2018, 12, 57–66. [Google Scholar] [CrossRef]
- Nagpal, V.; Rai, R.; Place, A.T.; Murphy, S.B.; Verma, S.K.; Ghosh, A.K.; Vaughan, D.E. MiR-125b Is Critical for Fibroblast-to-Myofibroblast Transition and Cardiac Fibrosis. Circulation 2016, 133, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Bie, Z.-D.; Sun, L.-Y.; Geng, C.-L.; Meng, Q.-G.; Lin, X.-J.; Wang, Y.-F.; Wang, X.-F.; Yang, J. MiR-125b regulates SFRP5 expression to promote growth and activation of cardiac fibroblasts. Cell Biol. Int. 2016, 40, 1224–1234. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Li, M.; Jiang, X.; Huang, X.; Gu, S.; Ye, J.; Zhu, L.; Hou, M.; Zan, T. Young fibroblast-derived exosomal microRNA-125b transfers beneficial effects on aged cutaneous wound healing. J. Nanobiotechnol. 2022, 20, 144. [Google Scholar] [CrossRef] [PubMed]
- Duru, N.; Zhang, Y.; Gernapudi, R.; Wolfson, B.; Lo, P.-K.; Yao, Y.; Zhou, Q. Loss of miR-140 is a key risk factor for radiation-induced lung fibrosis through reprogramming fibroblasts and macrophages. Sci. Rep. 2016, 6, 39572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Chen, Q.; Song, H.; Jiang, W.; Xie, S.; Huang, J.; Kang, G. MicroRNA-375 prevents TGF-beta-dependent transdifferentiation of lung fibroblasts via the MAP2K6/P38 pathway. Mol. Med. Rep. 2020, 22, 1803–1810. [Google Scholar] [CrossRef]
- Ji, Y.; Qiu, M.; Shen, Y.; Gao, L.; Wang, Y.; Sun, W.; Li, X.; Lu, Y.; Kong, X. MicroRNA-327 regulates cardiac hypertrophy and fibrosis induced by pressure overload. Int. J. Mol. Med. 2018, 41, 1909–1916. [Google Scholar] [CrossRef]
- Akerman, A.W.; Collins, E.N.; Peterson, A.R.; Collins, L.B.; Harrison, J.K.; DeVaughn, A.; Townsend, J.M.; Vanbuskirk, R.L.; Riopedre-Maqueira, J.; Reyes, A.; et al. miR-133a Replacement Attenuates Thoracic Aortic Aneurysm in Mice. J. Am. Heart Assoc. 2021, 10, 19862. [Google Scholar] [CrossRef]
- Wei, P.; Xie, Y.; Abel, P.W.; Huang, Y.; Ma, Q.; Li, L.; Hao, J.; Wolff, D.W.; Wei, T.; Tu, Y. Transforming growth factor (TGF)-beta 1-induced miR-133a inhibits myofibroblast differentiation and pulmonary fibrosis. Cell Death Dis. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.-Y.; Lu, M.; Liu, J.; Wang, Z.-N.; Wang, W.-W.; Li, Y.; Song, Z.-J.; Xu, L.; Liu, Q.; Li, F.-H. MicroRNA-144 regulates angiotensin II-induced cardiac fibroblast activation by targeting CREB. Exp. Ther. Med. 2020, 20, 2113–2121. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Zhang, W.; Liu, Y.; Li, Y.; Wang, K.; Zhang, Y.; Yang, C.; Li, X.; Shi, J.; et al. MicroRNA-192 regulates hypertrophic scar fibrosis by targeting SIP1. J. Mol. Histol. 2017, 48, 357–366. [Google Scholar] [CrossRef]
- Cui, J.; Qi, S.; Liao, R.; Su, D.; Wang, Y.; Xue, S. MiR-574-5p promotes the differentiation of human cardiac fibroblasts via regulating ARID3A. Biochem. Biophys. Res. Commun. 2020, 521, 427–433. [Google Scholar] [CrossRef]
- Xu, S.-S.; Ding, J.-F.; Shi, P.; Shi, K.-H.; Tao, H. DNMT1-Induced miR-152-3p Suppression Facilitates Cardiac Fibroblast Activation in Cardiac Fibrosis. Cardiovasc. Toxicol. 2021, 21, 984–999. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Bounds, K.R.; Chatterjee, P.; Gupta, S. MicroRNA-130a, a Potential Antifibrotic Target in Cardiac Fibrosis. J. Am. Heart Assoc. 2017, 6, e006763. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Carter, D.E.; Leask, A. miR-218 regulates focal adhesion kinase-dependent TGF beta signaling in fibroblasts. Mol. Biol. Cell. 2014, 25, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Xian, S.; Li, J.; Zhang, Z. miR-26b inhibits isoproterenol-induced cardiac fibrosis via the Keap1/Nrf2 signaling pathway. Exp. Ther. Med. 2020, 19, 2067–2074. [Google Scholar] [CrossRef]
- Levy, M.Z.; Rabinowicz, N.; Kohon, M.Y.; Shalom, A.; Berl, A.; Hornik-Lurie, T.; Drucker, L.; Matalon, S.T.; Levy, Y. MiRNAs in Systemic Sclerosis Patients with Pulmonary Arterial Hypertension: Markers and Effectors. Biomedicines 2022, 10, 629. [Google Scholar] [CrossRef]
- Heindryckx, F.; Binet, F.; Ponticos, M.; Rombouts, K.; Lau, J.; Kreuger, J.; Gerwins, P. Endoplasmic reticulum stress enhances fibrosis through IRE1 alpha-mediated degradation of miR-150 and XBP-1 splicing. EMBO Mol. Med. 2016, 8, 729–744. [Google Scholar] [CrossRef]
- Deng, P.; Chen, L.; Liu, Z.; Ye, P.; Wang, S.; Wu, J.; Yao, Y.; Sun, Y.; Huang, X.; Ren, L.; et al. MicroRNA-150 Inhibits the Activation of Cardiac Fibroblasts by Regulating c-Myb. Cell Physiol. Biochem. 2016, 38, 2103–2122. [Google Scholar] [CrossRef] [Green Version]
- Ren, L.; Yang, C.; Dou, Y.; Zhan, R.; Sun, Y.; Yu, Y. MiR-541-5p regulates lung fibrosis by targeting cyclic nucleotide phosphodiesterase 1A. Exp. Lung Res. 2017, 43, 249–258. [Google Scholar] [CrossRef]
- Liu, C.-M.; Liao, Y.-W.; Hsieh, P.-L.; Yu, C.-H.; Chueh, P.J.; Lin, T.; Yang, P.-Y.; Yu, C.-C.; Chou, M.-Y. miR-1246 as a therapeutic target in oral submucosa fibrosis pathogenesis. J. Formos. Med. Assoc. 2019, 118, 1093–1098. [Google Scholar] [CrossRef]
- Qu, C.; Liu, X.; Ye, T.; Wang, L.; Liu, S.; Zhou, X.; Wu, G.; Lin, J.; Shi, S.; Yang, B. miR-216a exacerbates TGF-beta-induced myofibroblast transdifferentiation via PTEN/AKT signaling. Mol. Med. Rep. 2019, 19, 5345–5352. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, H.; Fan, X.; Guo, J. Inhibition of miR-497 Attenuates Oral Submucous Fibrosis by Inhibiting Myofibroblast Transdifferentiation in Buccal Mucosal Fibroblasts. Oral Health Prev. Dent. 2022, 20, 339–348. [Google Scholar] [CrossRef]
- Chai, C.Y.; Tai, I.C.; Zhou, R.; Song, J.L.; Zhang, C.Y.; Sun, S.R. MicroRNA-9-5p inhibits proliferation and induces apoptosis of human hypertrophic scar fibroblasts through targeting peroxisome proliferator-activated receptor beta. Biol. Open. 2020, 9, bio051904. [Google Scholar] [CrossRef]
- Han, B.; Zhang, Y.; Xiao, Y.; Shi, B.; Wu, H.; Liu, D. Adipose-Derived Stem Cell-Derived Extracellular Vesicles Inhibit the Fibrosis of Fibrotic Buccal Mucosal Fibroblasts via the MicroRNA-375/FOXF1 Axis. Stem Cells Int. 2021, 2021, 9964159 . [Google Scholar] [CrossRef]
- Zeng, C.; Wang, Y.-L.; Xie, C.; Sang, Y.; Li, T.-J.; Zhang, M.; Wang, R.; Zhang, Q.; Zheng, L.; Zhuang, S.-M. Identification of a novel TGF-beta-miR-122-fibronectin 1/serum response factor signaling cascade and its implication in hepatic fibrogenesis. Oncotarget 2015, 6, 12224–12233. [Google Scholar] [CrossRef]
- Xu, M.; Fang, S.; Xie, A. Posttranscriptional control of PLOD1 in adipose-derived stem cells regulates scar formation through altering macrophage polarization. Ann. Transl. Med. 2021, 9, 4978. [Google Scholar] [CrossRef]
- Wang, Q.-G.; Cheng, B.C.-Y.; He, Y.-Z.; Li, L.-J.; Ling, Y.; Luo, G.; Wang, L.; Liang, S.; Zhang, Y. miR-320a in serum exosomes promotes myocardial fibroblast proliferation via regulating the PIK3CA/Akt/mTOR signaling pathway in HEH2 cells. Exp. Ther. Med. 2021, 22, 10305. [Google Scholar] [CrossRef]
- Huang, Y.; Xie, Y.; Bailey, K.; Toews, M.L.; Abel, P.W.; Tu, Y. TGF-beta 1-induced miR-424 Mediates Pulmonary Myofibrolast Differentiation via Targeting Slit2. FASEB J. 2019, 33, 114172. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.; Shi, J.; Liu, K.; Wang, X.; Jia, Y.; He, T.; Shen, K.; Wang, Y.; Liu, J.; et al. Exosomes derived from human adipose mesenchymal stem cells attenuate hypertrophic scar fibrosis by miR-192-5p/IL-17RA/Smad axis. Stem. Cell Res. Ther. 2021, 12, 221. [Google Scholar] [CrossRef]
- Wu, Y.; Luan, J.; Jiao, C.; Zhang, S.; Ma, C.; Zhang, Y.; Fu, J.; Lai, E.Y.; Kopp, J.B.; Pi, J.; et al. circHIPK3 Exacerbates Folic Acid-Induced Renal Tubulointerstitial Fibrosis by Sponging miR-30a. Front. Physiol. 2022, 12, 715567. [Google Scholar] [CrossRef]
- Ding, F.; Lu, L.; Wu, C.; Pan, X.; Liu, B.; Zhang, Y.; Wang, Y.; Wu, W.; Yan, B.; Zhang, Y.; et al. circHIPK3 prevents cardiac senescence by acting as a scaffold to recruit ubiquitin ligase to degrade HuR. Theranostics 2022, 12, 7550–7566. [Google Scholar] [CrossRef]
- He, X.; Ou, C. CircRNA circHIPK3: A novel therapeutic target for angiotensin II-induced cardiac fibrosis. Int. J. Cardiol. 2020, 312, 98. [Google Scholar] [CrossRef]
- Wu, Y.; Wu, M.; Yang, J.; Li, Y.; Peng, W.; Wu, M.; Yu, C.; Fang, M. Silencing CircHIPK3 Sponges miR-93-5p to Inhibit the Activation of Rac1/PI3K/AKT Pathway and Improves Myocardial Infarction-Induced Cardiac Dysfunction. Front. Cardiovasc. Med. 2021, 8, 645378. [Google Scholar] [CrossRef]
- Wu, N.; Li, C.; Xu, B.; Xiang, Y.; Jia, X.; Yuan, Z.; Wu, L.; Zhong, L.; Li, Y. Circular RNA mmu_circ_0005019 inhibits fibrosis of cardiac fibroblasts and reverses electrical remodeling of cardiomyocytes. BMC Cardiovasc. Disord. 2021, 21, 308. [Google Scholar] [CrossRef]
- Du, W.W.; Xu, J.; Yang, W.; Wu, N.; Li, F.; Zhou, L.; Wang, S.; Li, X.; He, A.T.; Du, K.Y.; et al. A Neuroligin Isoform Translated by circNlgn Contributes to Cardiac Remodeling. Circ. Res. 2021, 129, 568–582. [Google Scholar] [CrossRef]
- Zhang, L.; Chi, X.; Luo, W.; Yu, S.; Zhang, J.; Guo, Y.; Ren, Q.; Zhang, W. Lung myofibroblast transition and fibrosis is regulated by circ0044226. Int. J. Biochem. Cell Biol. 2020, 118, 105660. [Google Scholar] [CrossRef]
- Zheng, S.; Hu, C.; Lin, H.; Li, G.; Xia, R.; Zhang, X.; Su, D.; Li, Z.; Zhou, Q.; Chen, R. circCUL2 induces an inflammatory CAF phenotype in pancreatic ductal adenocarcinoma via the activation of the MyD88-dependent NF-kappa B signaling pathway. J. Exp. Clin. Cancer Res. 2022, 41, 71. [Google Scholar] [CrossRef]
- Zhou, B.; Yu, J.-W. A novel identified circular RNA, circRNA_010567, promotes myocardial fibrosis via suppressing miR-141 by targeting TGF-beta 1. Biochem. Biophys. Res. Commun. 2017, 487, 769–775. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, J.; Deng, H.; Ma, R.; Liao, J.-Y.; Liang, H.; Hu, J.; Li, J.; Guo, Z.; Cai, J.; et al. Targeting Mitochondria-Located circRNA SCAR Alleviates NASH via Reducing mROS Output. Cell 2020, 183, 76. [Google Scholar] [CrossRef]
- Ge, X.; Sun, Y.; Tang, Y.; Lin, J.; Zhou, F.; Yao, G.; Su, X. Circular RNA HECTD1 knockdown inhibits transforming growth factor-beta/small mothers against decapentaplegic (TGF-beta/Smad) signaling to reduce hypertrophic scar fibrosis. Bioengineered 2022, 13, 7303–7315. [Google Scholar] [CrossRef]
- Ding, K.; Li, D.; Zhang, R.; Zuo, M. Circ_0047339 promotes the activation of fibroblasts and affects the development of urethral stricture by targeting the miR-4691-5p/TSP-1 axis. Sci. Rep. 2022, 12, 14746. [Google Scholar] [CrossRef]
- Cheng, X.; Ai, K.; Yi, L.; Liu, W.; Li, Y.; Wang, Y.; Zhang, D. The mmu_circRNA_37492/hsa_circ_0012138 function as potential ceRNA to attenuate obstructive renal fibrosis. Cell Death Dis. 2022, 13, 207. [Google Scholar] [CrossRef] [PubMed]
- Savary, G.; Dewaeles, E.; Diazzi, S.; Buscot, M.; Nottet, N.; Fassy, J.; Courcot, E.; Henaoui, I.-S.; Lemaire, J.; Martis, N.; et al. The Long Noncoding RNA DNM3OS Is a Reservoir of FibromiRs with Major Functions in Lung Fibroblast Response to TGF-beta and Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2019, 200, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Piccoli, M.-T.; Gupta, S.K.; Viereck, J.; Foinquinos, A.; Samolovac, S.; Kramer, F.L.; Garg, A.; Remke, J.; Zimmer, K.; Batkai, S.; et al. Inhibition of the Cardiac Fibroblast- Enriched lncRNA Meg3 Prevents Cardiac Fibrosis and Diastolic Dysfunction. Circ. Res. 2017, 121, 575. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, X.; Wang, T.; Li, J.; Su, Q.; Zhong, C.; Chen, Z.; Liang, Y. The MIR155 host gene/microRNA-627/HMGB1/NF-kappa B loop modulates fibroblast proliferation and extracellular matrix deposition. Life Sci. 2021, 269. [Google Scholar] [CrossRef]
- Su, H.; Xie, J.; Wen, L.; Wang, S.; Chen, S.; Li, J.; Qi, C.; Zhang, Q.; He, X.; Zheng, L.; et al. LncRNA Gas5 regulates Fn1 deposition via Creb5 in renal fibrosis. Epigenomics 2021, 13, 699–713. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Liao, Y.-W.; Hsieh, P.-L.; Lu, M.-Y.; Peng, C.-Y.; Chu, P.-M.; Yang, H.-W.; Huang, Y.-F.; Yu, C.-C.; Yu, C.-H. LncRNA GAS5-AS1 inhibits myofibroblasts activities in oral submucous fibrosis. J. Formos. Med. Assoc. 2018, 117, 727–733. [Google Scholar] [CrossRef]
- Zhou, X.-H.; Chai, H.-X.; Bai, M.; Zhang, Z. LncRNA-GAS5 regulates PDCD4 expression and mediates myocardial infarction-induced cardiomyocytes apoptosis via targeting MiR-21. Cell Cycle 2020, 19, 1363–1377. [Google Scholar] [CrossRef]
- Tao, H.; Shi, P.; Zhao, X.-D.; Xuan, H.-Y.; Ding, X.-S. MeCP2 inactivation of LncRNA GAS5 triggers cardiac fibroblasts activation in cardiac fibrosis. Cell Signalling 2020, 74. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, R.; Liang, Y.; Fu, X.; Wang, D.; Wang, C. Blockade of lncRNA-ASLNCS5088-enriched exosome generation in M2 macrophages by GW4869 dampens the effect of M2 macrophages on orchestrating fibroblast activation. FASEB J. 2019, 33, 12200–12212. [Google Scholar] [CrossRef] [Green Version]
- Wasson, C.W.; Abignano, G.; Hermes, H.; Malaab, M.; Ross, R.L.; Jimenez, S.A.; Chang, H.Y.; Feghali-Bostwick, C.A.; del Galdo, F. Long non-coding RNA HOTAIR drives EZH2-dependent myofibroblast activation in systemic sclerosis through miRNA 34a-dependent activation of NOTCH. Ann. Rheum. Dis. 2020, 79, 507–517. [Google Scholar] [CrossRef]
- Chen, T.; Guo, Y.; Wang, J.; Ai, L.; Ma, L.; He, W.; Li, Z.; Yu, X.; Li, J.; Fan, X.; et al. LncRNA CTD-2528L19.6 prevents the progression of IPF by alleviating fibroblast activation. Cell Death Dis. 2021, 12, 600. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Tai, W.; Lu, N.; Li, T.; Liu, Y.; Wu, W.; Li, Z.; Pu, L.; Zhao, X.; Zhang, T.; et al. lncRNA ZFAS1 promotes lung fibroblast-to-myofibroblast transition and ferroptosis via functioning as a ceRNA through miR-150-5p/SLC38A1 axis. Aging 2020, 12, 9085–9102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Fu, X.; Kataoka, M.; Liu, N.; Wang, Y.; Gao, F.; Liang, T.; Dong, X.; Pei, J.; Hu, X.; et al. Long noncoding RNA Cfast regulates cardiac fibrosis. Mol. Ther.-Nucleic Acids 2021, 23, 377–392. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Pan, Z.; Zhao, X.; Liu, L.; Sun, J.; Su, X.; Xu, C.; Zhou, Y.; Zhao, D.; Xu, B.; et al. LncRNA PFL contributes to cardiac fibrosis by acting as a competing endogenous RNA of let-7d. Theranostics 2018, 8, 1180–1194. [Google Scholar] [CrossRef]
- Bijkerk, R.; Au, Y.W.; Stam, W.; Duijs, J.M.G.J.; Koudijs, A.; Lievers, E.; Rabelink, T.J.; van Zonneveld, A.J. Long Non-coding RNAs Rian and Miat Mediate Myofibroblast Formation in Kidney Fibrosis. Front. Pharmacol. 2019, 10, 215. [Google Scholar] [CrossRef] [Green Version]
- Xiao, T.; Zou, Z.; Xue, J.; Syed, B.M.; Sun, J.; Dai, X.; Shi, M.; Li, J.; Wei, S.; Tang, H.; et al. LncRNA H19-mediated M2 polarization of macrophages promotes myofibroblast differentiation in pulmonary fibrosis induced by arsenic exposure. Environ. Pollut. 2021, 268, 115810. [Google Scholar] [CrossRef]
- Li, W.; Cheng, B. Knockdown of LncRNA NEAT1 inhibits myofibroblast activity in oral submucous fibrosis through miR-760/TPM1 axis. J. Dent. Sci. 2022, 17, 707–717. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, H.; Chen, H.; Li, H.; Xu, P.; Liu, B.; Zhang, Q.; Lv, C.; Song, X. ATF3-activated accelerating effect of LINC00941/lncIAPF on fibroblast-to-myofibroblast differentiation by blocking autophagy depending on ELAVL1/HuR in pulmonary fibrosis. Autophagy 2022, 18, 2636–2655. [Google Scholar] [CrossRef]
- Cai, W.; Xu, H.; Zhang, B.; Gao, X.; Li, S.; Wei, Z.; Li, S.; Mao, N.; Jin, F.; Li, Y.; et al. Differential expression of lncRNAs during silicosis and the role of LOC103691771 in myofibroblast differentiation induced by TGF-beta 1. Biomed. Pharmacother. 2020, 125, 109980. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, S.; Li, X.; Gong, M. LncRNA SNHG7 promotes cardiac remodeling by upregulating ROCK1 via sponging miR-34-5p. Aging 2020, 12, 10441–10456. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Yu, C.-C.; Hsieh, P.-L.; Liao, Y.-W.; Yu, C.-H.; Su, T.-R. Inhibition of lncRNA HOTTIP ameliorated myofibroblast activities and inflammatory cytokines in oral submucous fibrosis. J. Formos. Med. Assoc. 2021, 120, 1188–1193. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Jiao, B.; Gui, W.; Zhang, Q.; Wang, F.; Han, L. Long non-coding RNA SNHG1 promotes fibroblast-to-myofibroblast transition during the development of pulmonary fibrosis induced by silica particles exposure. Ecotoxicol. Environ. Saf. 2021, 228, 112938. [Google Scholar] [CrossRef] [PubMed]
- Kong, N.; Bao, Y.; Zhao, H.; Kang, X.; Tai, X.; Chen, X.; Guo, W.; Shen, Y. Long Noncoding RNA LINC01518 Modulates Proliferation and Migration in TGF-beta 1-Treated Human Tenon Capsule Fibroblast Cells Through the Regulation of hsa-miR-216b-5p. Neuromolecular Med. 2022, 24, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-H.; Fang, C.-Y.; Yu, C.-C.; Hsieh, P.-L.; Liao, Y.-W.; Tsai, L.-L.; Chu, P.-M. LINC00312/YBX1 Axis Regulates Myofibroblast Activities in Oral Submucous Fibrosis. Int. J. Mol. Sci. 2020, 21, 2979. [Google Scholar] [CrossRef]
- Zhang, L.; Gao, J.; Gong, A.; Dong, Y.; Hao, X.; Wang, X.; Zheng, J.; Ma, W.; Song, Y.; Zhang, J.; et al. The Long Noncoding RNA LINC00963 Inhibits Corneal Fibrosis Scar Formation by Targeting miR-143-3p. DNA Cell Biol. 2021, 41, 1034. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Zhou, Y.; Wen, D.; Wang, J. Noncoding RNAs: Master Regulator of Fibroblast to Myofibroblast Transition in Fibrosis. Int. J. Mol. Sci. 2023, 24, 1801. https://doi.org/10.3390/ijms24021801
Zhang H, Zhou Y, Wen D, Wang J. Noncoding RNAs: Master Regulator of Fibroblast to Myofibroblast Transition in Fibrosis. International Journal of Molecular Sciences. 2023; 24(2):1801. https://doi.org/10.3390/ijms24021801
Chicago/Turabian StyleZhang, Huamin, Yutong Zhou, Dada Wen, and Jie Wang. 2023. "Noncoding RNAs: Master Regulator of Fibroblast to Myofibroblast Transition in Fibrosis" International Journal of Molecular Sciences 24, no. 2: 1801. https://doi.org/10.3390/ijms24021801