
Jasmonic Acid-Induced β-Cyclocitral Confers Resistance to Bacterial Blight and Negatively Affects Abscisic Acid Biosynthesis in Rice


Abstract
:1. Introduction
2. Results
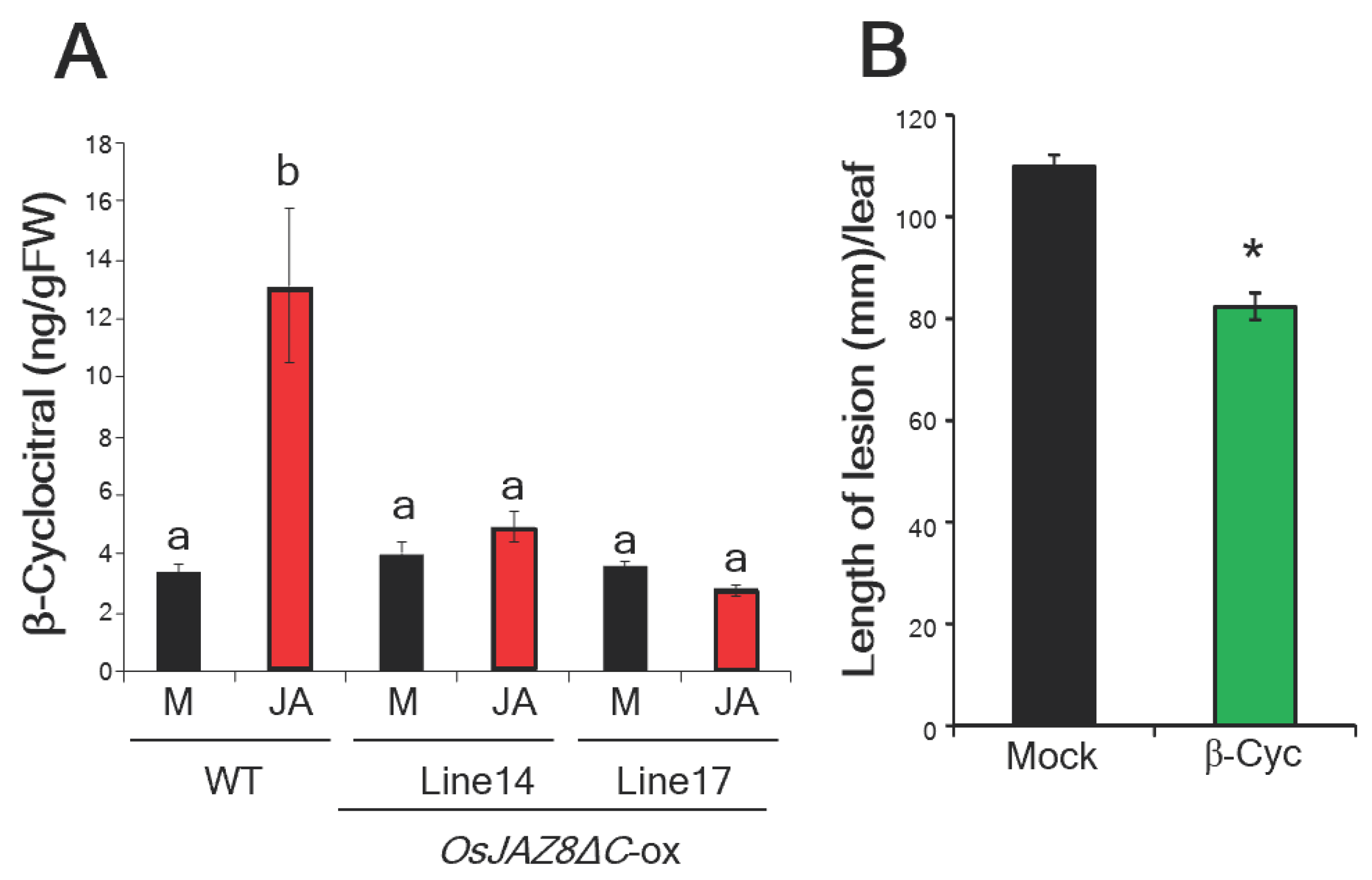
2.1. JA-Responsive β-Cyc Induces Resistance to Rice Bacterial Blight
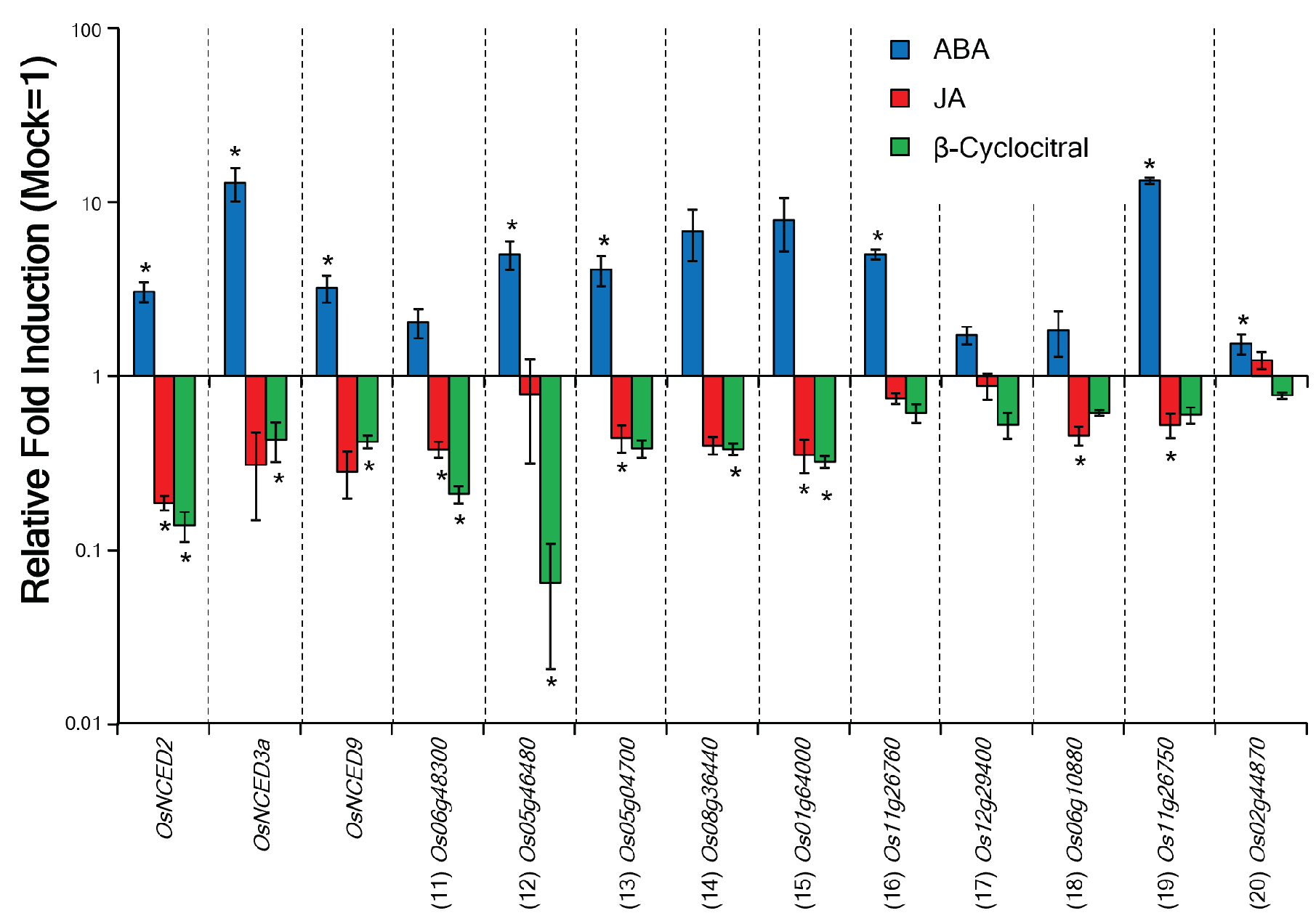
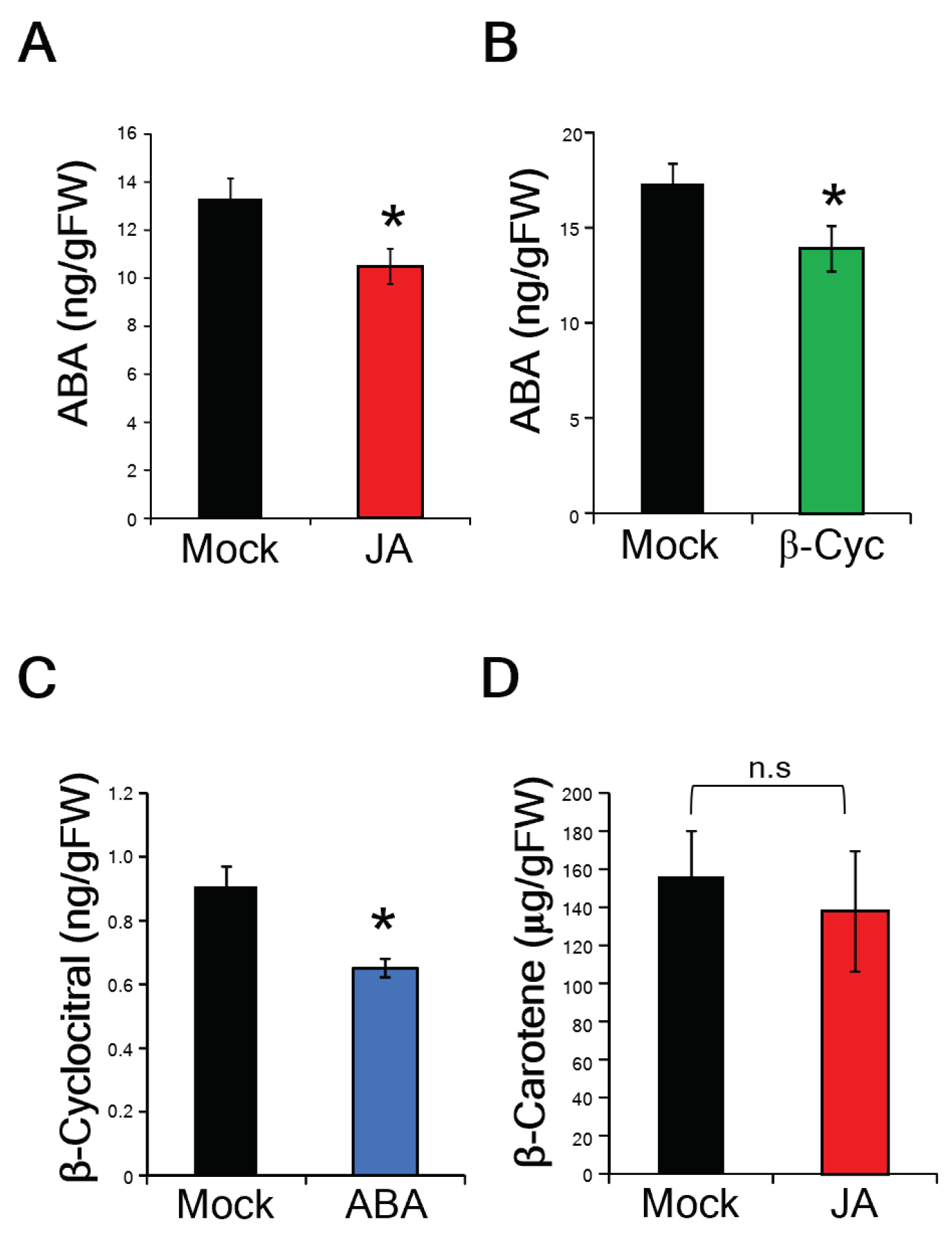
2.2. β-Cyc Negatively Affects the Expression of ABA-Responsive Genes and ABA Biosynthesis
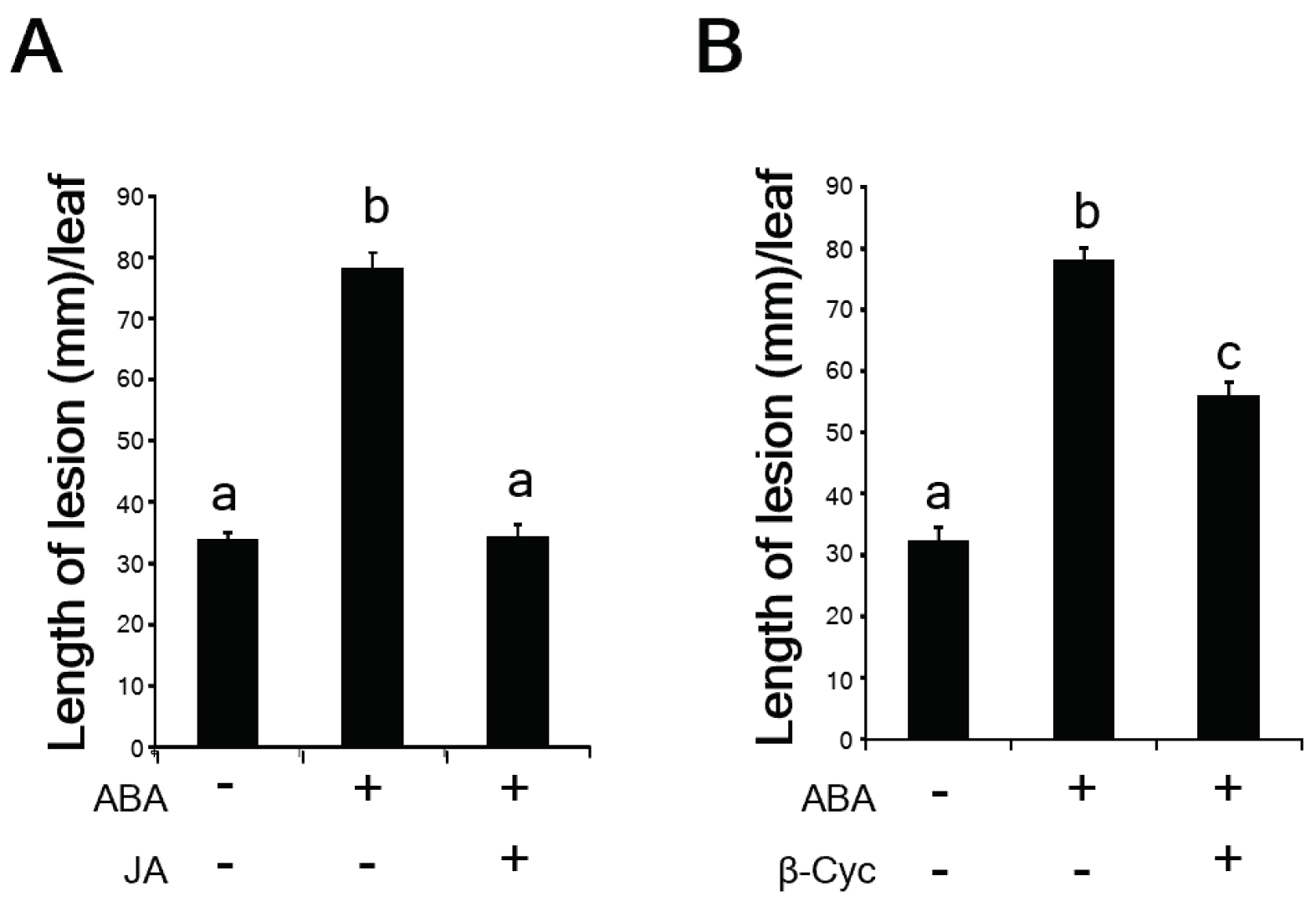
2.3. β-Cyc Negatively Affects ABA-Induced Susceptibility to Xoo
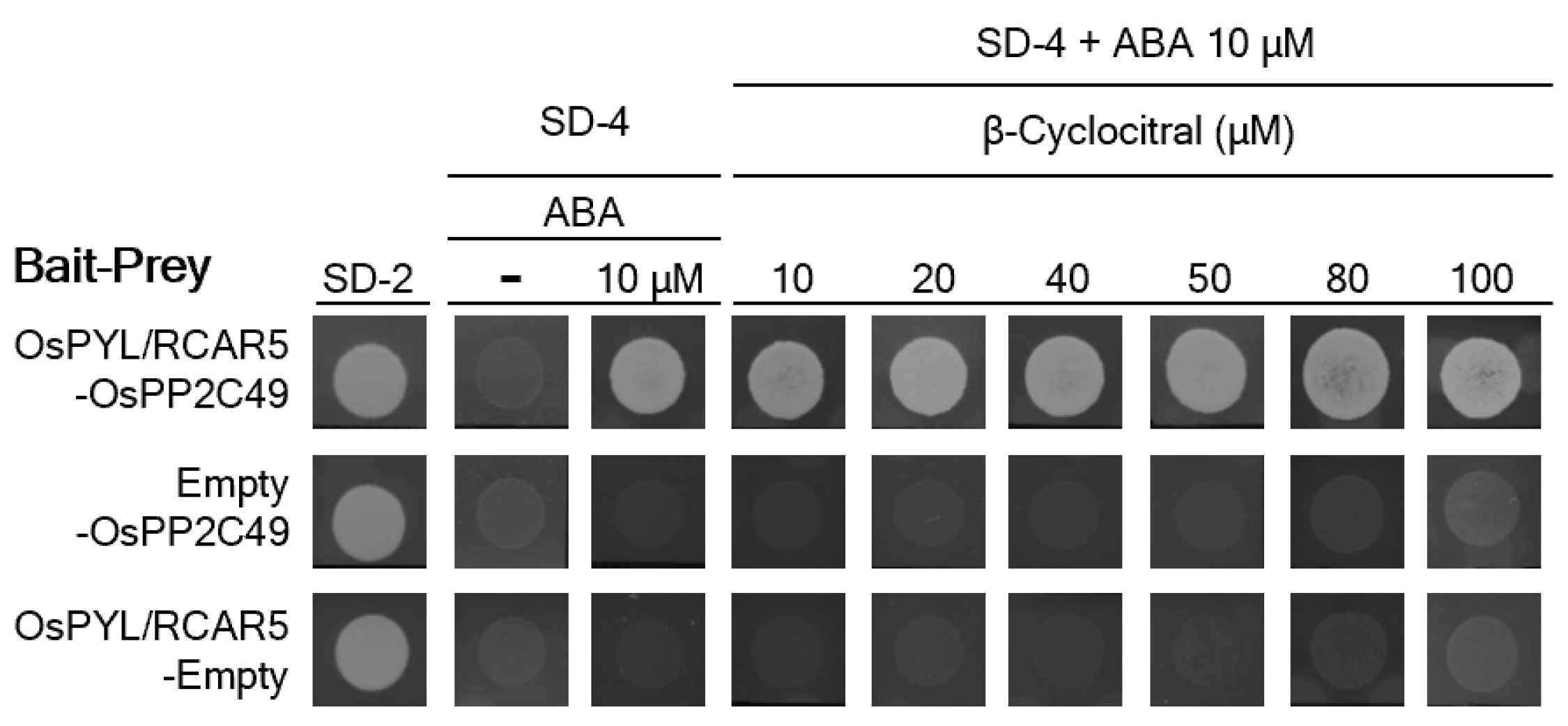
2.4. β-Cyc Has No Effect on ABA Perception on OsPYL/RCAR5
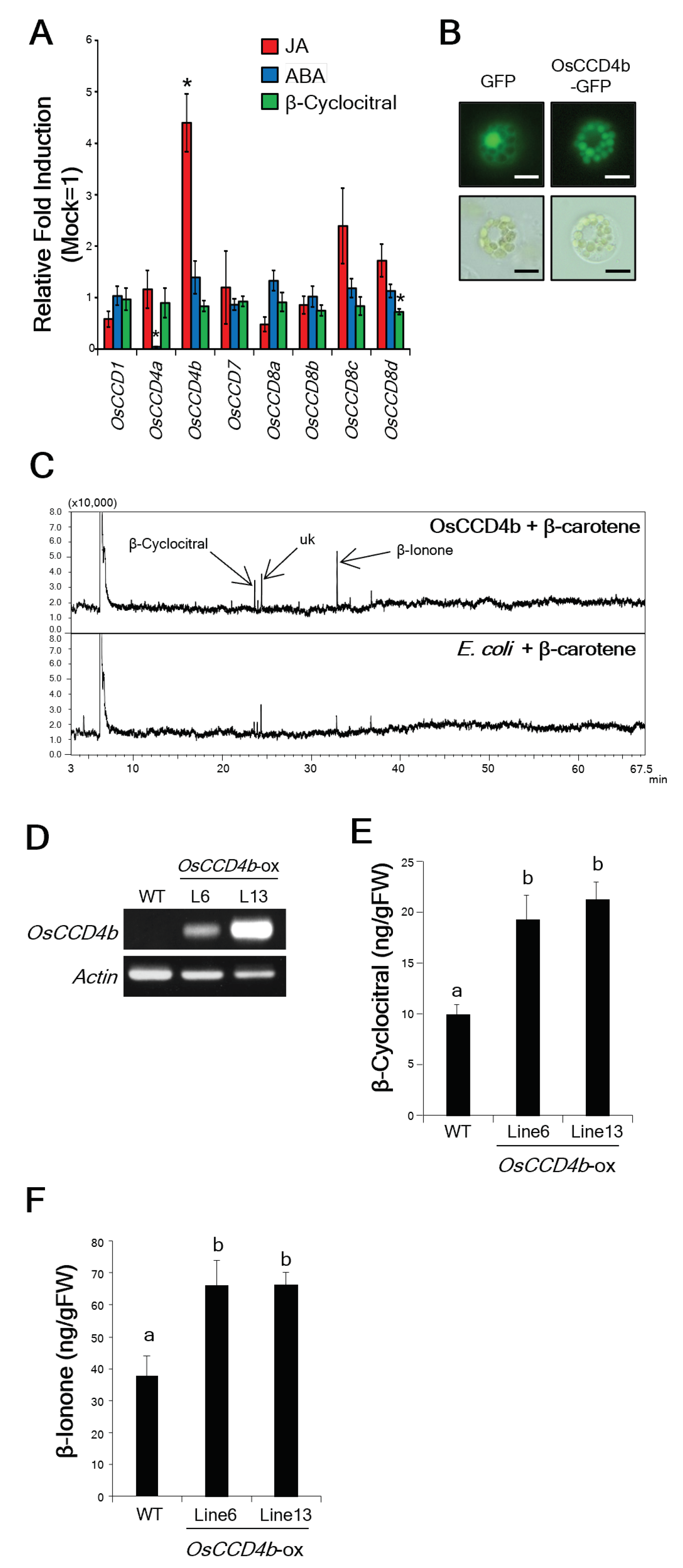
2.5. Identification of a Rice β-Cyc Biosynthesis Gene, OsCCD4b
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Chemical Treatments, and Bacterial Inoculation
4.2. Microarray Analysis
4.3. Quantification of Plant Volatiles, ABA, and β-Carotene
4.4. Reverse Transcription-Quantitative PCR (RT-qPCR)
4.5. Yeast Two-Hybrid (Y2H) System
4.6. Transient Localization Assay
4.7. Functional Expression of OsCCD4b in Escherichia coli
4.8. Enzyme Assay and Analysis of Volatiles
4.9. Production of OsCCD4b-Overexpressing Rice Plants
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trang Nguyen, H.; Thi Mai To, H.; Lebrun, M.; Bellafiore, S.; Champion, A. Jasmonates-the master regulator of rice development, adaptation, and defense. Plants 2019, 8, 339. [Google Scholar] [CrossRef] [Green Version]
- Vleesschauwer, D.D.; Gheysen, G.; Höfte, M. Hormone defense networking in rice: Tales from a different world. Trends Plant Sci. 2013, 18, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Tariq, R.; Ji, Z.; Wang, C.; Tang, Y.; Zou, L.; Sun, H.; Chen, G.; Zhao, K. RNA-Seq analysis of gene expression changes triggered by Xanthomonas oryzae pv. oryzae in a susceptible rice genotype. Rice 2019, 12, 44. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, A.; Vadassery, J.; Patel, H.K.; Pandey, A.; Palaparthi, R.; Mithöfer, A.; Sonti, R.V. Upregulation of jasmonate biosynthesis and jasmonate-responsive genes in rice leaves in response to a bacterial pathogen mimic. Funct. Integr. Genomics 2015, 15, 363–373. [Google Scholar] [CrossRef]
- Uji, Y.; Taniguchi, S.; Tamaoki, D.; Shishido, H.; Akimitsu, K.; Gomi, K. Overexpression of OsMYC2 results in the up-regulation of early JA-responsive genes and bacterial blight resistance in rice. Plant Cell Physiol. 2016, 57, 1814–1827. [Google Scholar] [CrossRef]
- Onohata, T.; Gomi, K. Overexpression of jasmonate-responsive OsbHLH034 in rice results in the induction of bacterial blight-resistance via an increase in lignin biosynthesis. Plant Cell Rep. 2020, 39, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Valea, I.; Motegi, A.; Kawamura, N.; Kawamoto, K.; Miyao, A.; Ozawa, R.; Takabayashi, J.; Gomi, K.; Nemoto, K.; Nozawa, A.; et al. The rice wound-inducible transcription factor RERJ1 sharing same signal transduction pathway with OsMYC2 is necessary for defense response to herbivory and bacterial blight. Plant Mol. Biol. 2022, 109, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Kashihara, K.; Onohata, T.; Okamoto, Y.; Uji, Y.; Mochizuki, S.; Akimitsu, K.; Gomi, K. Overexpression of OsNINJA1 negatively affects a part of OsMYC2-mediated abiotic and biotic responses in rice. J. Plant Physiol. 2019, 232, 180–187. [Google Scholar] [CrossRef]
- Pauwels, L.; Goossens, A. The JAZ proteins: A crucial interface in the jasmonate signaling cascade. Plant Cell 2011, 23, 3089–3100. [Google Scholar] [CrossRef] [Green Version]
- Yamada, S.; Kano, A.; Tamaoki, D.; Miyamoto, A.; Shishido, H.; Miyoshi, S.; Taniguchi, S.; Akimitsu, K.; Gomi, K. Involvement of OsJAZ8 in jasmonate-induced resistance to bacterial blight in rice. Plant Cell Physiol. 2012, 53, 2060–2072. [Google Scholar] [CrossRef]
- Kashihara, K.; Onohata, T.; Yariuchi, R.; Tanaka, S.; Akimitsu, K.; Gomi, K. The overexpression of OsSRO1a, which encodes an OsNINJA1- and OsMYC2-interacting protein, negatively affects OsMYC2-mediated jasmonate signaling in rice. Plant Cell Rep. 2020, 39, 489–500. [Google Scholar] [CrossRef]
- Kashihara, K.; Ojiri, T.; Gomi, K. Overexpression of OsNINJA1-interacting protein OsFHA1 affects jasmonate-mediated OsMYC2 signaling but not disease resistance in rice. J. Gen. Plant Pathol. 2022, 88, 225–231. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, Y.; Tang, L.; Tong, X.; Wang, L.; Liu, L.; Huang, S.; Zhang, J. SAPK10-mediated phosphorylation on WRKY72 releases its suppression on jasmonic acid biosynthesis and bacterial blight resistance. iScience 2019, 16, 499–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, Y.; Kang, Y.; Wu, M.; Liu, H.; Hui, S.; Zhang, Q.; Li, X.; Xiao, J.; Wang, S. Jasmonic acid-involved OsEDS1 signaling in rice-bacteria interactions. Rice 2019, 12, 25. [Google Scholar] [CrossRef] [Green Version]
- Tabei, H. Anatomical studies of rice plant affected with bacterial leaf blight, with special reference to stomatal infection at the coleoptile and the foliage leaf sheath of rice seedling. Ann. Phytopath. Sci. Jpn. 1967, 33, 12–16. [Google Scholar] [CrossRef]
- Hilaire, E.; Young, S.A.; Willard, L.H.; McGee, J.D.; Sweat, T.; Chittoor, J.M.; Guikema, J.A.; Leach, J.E. Vascular defense responses in rice: Peroxidase accumulation in xylem parenchyma cells and xylem wall thickening. Mol. Plant-Microbe Interact. 2001, 14, 1411–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bart, R.S.; Chern, M.; Vega-Sánchez, M.E.; Canlas, P.; Ronald, P.C. Rice Snl6, a cinnamoyl-CoA reductase-like gene family member, is required for NH1-mediated immunity to Xanthomonas oryzae pv. oryzae. PLoS Genet. 2010, 6, e1001123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, A.; Xue, G.; Cui, P.; Fan, F.; Liu, H.; Yin, C.; Sun, W.; Liang, Y. The role of silicon in enhancing resistance to bacterial blight of hydroponic- and soil-cultured rice. Sci. Rep. 2016, 6, 24640. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, G.; Fukuda, M.; Lucob-Agustin, N.; Inukai, Y.; Gomi, K. The Mutation of Rice MEDIATOR25, OsMED25, Induces Rice Bacterial Blight Resistance through Altering Jasmonate- and Auxin-Signaling. Plants 2022, 11, 1601. [Google Scholar] [CrossRef] [PubMed]
- Girija, A.M.; Kinathi, B.K.; Madhavi, M.B.; Ramesh, P.; Vungarala, S.; Patel, H.K.; Sonti, R.V. Rice leaf transcriptional profiling suggests a functional interplay between Xanthomonas oryzae pv. oryzae lipopolysaccharide and extracellular polysaccharide in modulation of defense responses during infection. Mol. Plant-Microbe Interact. 2017, 30, 16–27. [Google Scholar] [CrossRef]
- Taniguchi, S.; Hosokawa-Shinonaga, Y.; Tamaoki, D.; Yamada, S.; Akimitsu, K.; Gomi, K. Jasmonate induction of the monoterpene linalool confers resistance to rice bacterial blight and its biosynthesis is regulated by JAZ protein in rice. Plant Cell Environ. 2014, 37, 451–461. [Google Scholar] [CrossRef]
- Yoshitomi, K.; Taniguchi, S.; Tanaka, K.; Uji, Y.; Akimitsu, K.; Gomi, K. Rice terpene synthase 24 (OsTPS24) encodes a jasmonate-responsive monoterpene synthase that produces an antibacterial γ-terpinene against rice pathogen. J. Plant Physiol. 2016, 191, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Kiryu, M.; Hamanaka, M.; Yoshitomi, K.; Mochizuki, S.; Akimitsu, K.; Gomi, K. Rice terpene synthase 18 (OsTPS18) encodes a sesquiterpene synthase that produces an antibacterial (E)-nerolidol against a bacterial pathogen of rice. J. Gen. Plant Pathol. 2018, 84, 221–229. [Google Scholar] [CrossRef]
- Kiyama, H.; Matsunaga, A.; Suzuki, G.; Gomi, K. Monoterpene geraniol produced by Rice terpene synthase 21 suppresses the expression of cell-division related genes in the rice bacterial pathogen, Xanthomonas oryzae pv. oryzae. Physiol. Mol. Plant Pathol. 2021, 115, 101673. [Google Scholar] [CrossRef]
- Tanaka, K.; Taniguchi, S.; Tamaoki, D.; Yoshitomi, K.; Akimitsu, K.; Gomi, K. Multiple roles of plant volatiles in jasmonate-induced defense response in rice. Plant Signal. Behav. 2014, 9, e29247. [Google Scholar] [CrossRef] [Green Version]
- Gomi, K.; Satoh, M.; Ozawa, R.; Shinonaga, Y.; Sanada, S.; Sasaki, K.; Matsumura, M.; Ohashi, Y.; Kanno, H.; Akimitsu, K.; et al. Role of hydroperoxide lyase in white-backed planthopper (Sogatella furcifera Horváth)-induced resistance to bacterial blight in rice, Oryza sativa L. Plant J. 2010, 61, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Audenaert, K.; Hofte, M.; Vlesschauwer, D.D. Abscisic Acid Promotes Susceptibility to the Rice Leaf Blight Pathogen Xanthomonas oryzae pv oryzae by Suppressing Salicylic Acid-Mediated Defenses. PLoS ONE 2013, 8, e67413. [Google Scholar] [CrossRef] [Green Version]
- Rabbani, M.A.; Maruyama, K.; Abe, H.; Khan, M.A.; Katsura, K.; Ito, Y.; Yoshiwara, K.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Monitoring expression profiles of rice genes under cold, drought, and high-salinity stresses and abscisic acid application using cDNA microarray and RNA gel-blot analyses. Plant Physiol. 2003, 133, 1755–1767. [Google Scholar] [CrossRef] [Green Version]
- Vallabhaneni, R.; Wurtzel, E.T. From epoxycarotenoids to ABA: The role of ABA 8′-hydroxylases in drought-stressed maize roots. Arch. Biochem. Biophys. 2010, 504, 112–117. [Google Scholar] [CrossRef] [Green Version]
- Auldridge, M.E.; McCarty, D.R.; Klee, H.J. Plant carotenoid cleavage oxygenases and their apocarotenoid products. Curr. Opin. Plant Biol. 2006, 9, 315–321. [Google Scholar] [CrossRef]
- Kim, H.; Hwang, H.; Hong, J.W.; Lee, Y.N.; Ahn, I.P.; Yoon, I.S.; Yoo, S.D.; Lee, S.; Lee, S.C.; Kim, B.G. A rice orthologue of the ABA receptor, OsPYL/RCAR5, is a positive regulator of the ABA signal transduction pathway in seed germination and early seedling growth. J. Exp. Bot. 2012, 63, 1013–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio-Moraga, A.; Rambla, J.L.; Fernández-de-Carmen, A.; Trapero-Mozos, A.; Ahrazem, O.; Orzáez, D.; Granell, A.; Gómez-Gómez, L. New target carotenoids for CCD4 enzymes are revealed with the characterization of a novel stress-induced carotenoid cleavage dioxygenase gene from Crocus sativus. Plant Mol. Biol. 2014, 86, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Yuan, H.; Cao, H.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid Metabolism in Plants: The Role of Plastids. Mol. Plant. 2018, 11, 58–74. [Google Scholar] [CrossRef] [Green Version]
- Passardi, F.; Longet, D.; Penel, C.; Dunand, C. The class III peroxidase multigenic family in rice and its evolution in land plants. Phytochemistry 2004, 65, 1879–1893. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [Green Version]
- Aki, T.; Shigyo, M.; Nakano, R.; Yoneyama, T.; Yanagisawa, S. Nano scale proteomics revealed the presence of regulatory proteins including three FT-Like proteins in phloem and xylem saps from rice. Plant Cell Physiol. 2008, 49, 767–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Zhou, Z.; Yin, D.; Lv, Q.; Wang, L.; Xu, X.; Wang, J.; Li, X.; Zhao, X.; Jiang, G.; et al. Transgenic rice plants overexpressing BBTI4 confer partial but broad-spectrum bacterial blight resistance. J. Plant Biol. 2013, 56, 383–390. [Google Scholar] [CrossRef]
- Zhang, D.; Tian, C.; Yin, K.; Wang, W.; Qiu, J.L. Postinvasive Bacterial Resistance Conferred by Open Stomata in Rice. Mol. Plant-Micribe Interact. 2019, 32, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yan, S.; Huang, W.; Yang, J.; Dong, J.; Zhang, S.; Zhao, J.; Yang, T.; Mao, X.; Zhu, X.; et al. NAC transcription factor ONAC066 positively regulates disease resistance by suppressing the ABA signaling pathway in rice. Plant Mol. Biol. 2018, 98, 289–302. [Google Scholar] [CrossRef]
- Nahar, K.; Kyndt, T.; Nzogela, Y.B.; Gheysen, G. Abscisic acid interacts antagonistically with classical defense pathways in rice–migratory nematode interaction. New Phytol. 2012, 196, 901–913. [Google Scholar] [CrossRef]
- Xie, K.; Li, L.; Zhang, H.; Wang, R.; Tan, X.; He, Y.; Hong, G.; Li, J.; Ming, F.; Yao, X.; et al. Abscisic acid negatively modulates plant defence against rice black-streaked dwarf virus infection by suppressing the jasmonate pathway and regulating reactive oxygen species levels in rice. Plant Cell Environ. 2018, 41, 2504–2514. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Estrada-Tejedor, R.; Volke, D.C.; Phillips, M.A.; Gershenzon, J.; Wright, L.P. Negative regulation of plastidial isoprenoid pathway by herbivore-induced β-cyclocitral in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2021, 118, e2008747118. [Google Scholar] [CrossRef] [PubMed]
- Shimono, M.; Sugano, S.; Nakayama, A.; Jiang, C.J.; Ono, K.; Toki, S.; Takatsuji, H. Rice WRKY45 Plays a Crucial Role in Benzothiadiazole-Inducible Blast Resistance. Plant Cell 2007, 19, 2064–2076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uji, Y.; Kashihara, K.; Kiyama, H.; Mochizuki, S.; Akimitsu, K.; Gomi, K. Jasmonic acid-induced VQ-motif-containing protein OsVQ13 influences the OsWRKY45 signaling pathway and grain size by associating with OsMPK6 in rice. Int. J. Mol. Sci. 2019, 20, 2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Zhong, S.; Li, Q.; Zhu, Z.; Lou, Y.; Wang, L.; Wang, J.; Wang, M.; Li, Q.; Yang, D.; et al. Functional analysis of rice NPR1-like genes reveals that OsNPR1/NH1 is the rice orthologue conferring disease resistance with enhanced herbivore susceptibility. Plant Biotechnol. J. 2007, 5, 313–324. [Google Scholar] [CrossRef]
- Liu, Q.; Ning, Y.; Zhang, Y.; Yu, N.; Zhao, C.; Zhan, X.; Wu, W.; Chen, D.; Wei, X.; Wang, G.L.; et al. OsCUL3a Negatively Regulates Cell Death and Immunity by Degrading OsNPR1 in Rice. Plant Cell 2017, 29, 345–359. [Google Scholar] [CrossRef] [Green Version]
- Tamaoki, D.; Seo, S.; Yamada, S.; Kano, A.; Miyamoto, A.; Shishido, H.; Miyoshi, S.; Taniguchi, S.; Akimitsu, K.; Gomi, K. Jasmonic acid and salicylic acid activate a common defense system in rice. Plant Signaling Behav. 2013, 8, e24260. [Google Scholar] [CrossRef]
- Havaux, M. β-Cyclocitral and derivatives: Emerging molecular signals serving multiple biological functions. Plant Physiol. Biochem. 2020, 155, 35–41. [Google Scholar] [CrossRef]
- Nambara, E.; Marion-Poll, A. Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 2005, 56, 165–185. [Google Scholar] [CrossRef] [Green Version]
- Zwanenburg, B.; Blanco-Ania, D. Strigolactones: New plant hormones in the spotlight. J. Exp. Bot. 2018, 69, 2205–2218. [Google Scholar] [CrossRef]
- Ramel, F.; Mialoundama, A.S.; Havaux, M. Nonenzymic carotenoid oxidation and photooxidative stress signalling in plants. J. Exp. Bot. 2013, 64, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, M.J.; Alquézar, B.; Alós, E.; Medina, V.; Carmona, L.; Bruno, M.; Al-Babili, S.; Zacarías, L. A novel carotenoid cleavage activity involved in the biosynthesis of Citrus fruit-specific apocarotenoid pigments. J. Exp. Bot. 2013, 64, 4461–4478. [Google Scholar] [CrossRef] [PubMed]
- Kauffman, H.E.; Reddy, A.P.K.; Hsieh, S.P.Y.; Merca, S.D. An improved technique for evaluating resistance of rice varieties to Xanthomonas oryzae. Plant Dis. Rep. 1973, 57, 537–541. [Google Scholar]
- Zhou, K.; Ren, Y.; Lv, J.; Wang, Y.; Liu, F.; Zhou, F.; Zhao, S.; Chen, S.; Peng, C.; Zhang, X.; et al. Young Leaf Chlorosis 1, a chloroplast-localized gene required for chlorophyll and lutein accumulation during early leaf development in rice. Planta 2013, 237, 279–292. [Google Scholar] [CrossRef]
- Mitsuhara, I.; Ugaki, M.; Hirochika, H.; Ohshima, M.; Murakami, T.; Gotoh, Y.; Katayose, Y.; Nakamura, S.; Honkura, R.; Nishimiya, S.; et al. Efficient promoter cassettes for enhanced expression of foreign genes in dicotyledonous and monocotyledonous plants. Plant Cell Physiol. 1996, 37, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Bart, R.; Chern, M.; Park, C.J.; Bartley, L.; Ronald, P.C. A novel system for gene silencing using siRNAs in rice leaf and stem-derived protoplasts. Plant Methods 2006, 2, 13. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, Y.; Nishio, Z.; Nakazono, K.; Soma, M.; Nakajima, E.; Ugaki, M.; Hibi, T. Enhanced resistance to blast (Magnaporthe grisea) in transgenic Japonica rice by constitutive expression of rice chitinase. Theor. Appl. Genet. 1999, 99, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Wen-jun, S.; Forde, B.G. Efficient transformation of Agrobacterium spp. by high voltage electroporation. Nucleic Acids Res. 1989, 17, 8385. [Google Scholar] [CrossRef]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT + β-Cyc/WT | ||||
|---|---|---|---|---|
| Accession Number | Gene Name | Fold Change * | q-Value | Reference |
| Defense-related gene | ||||
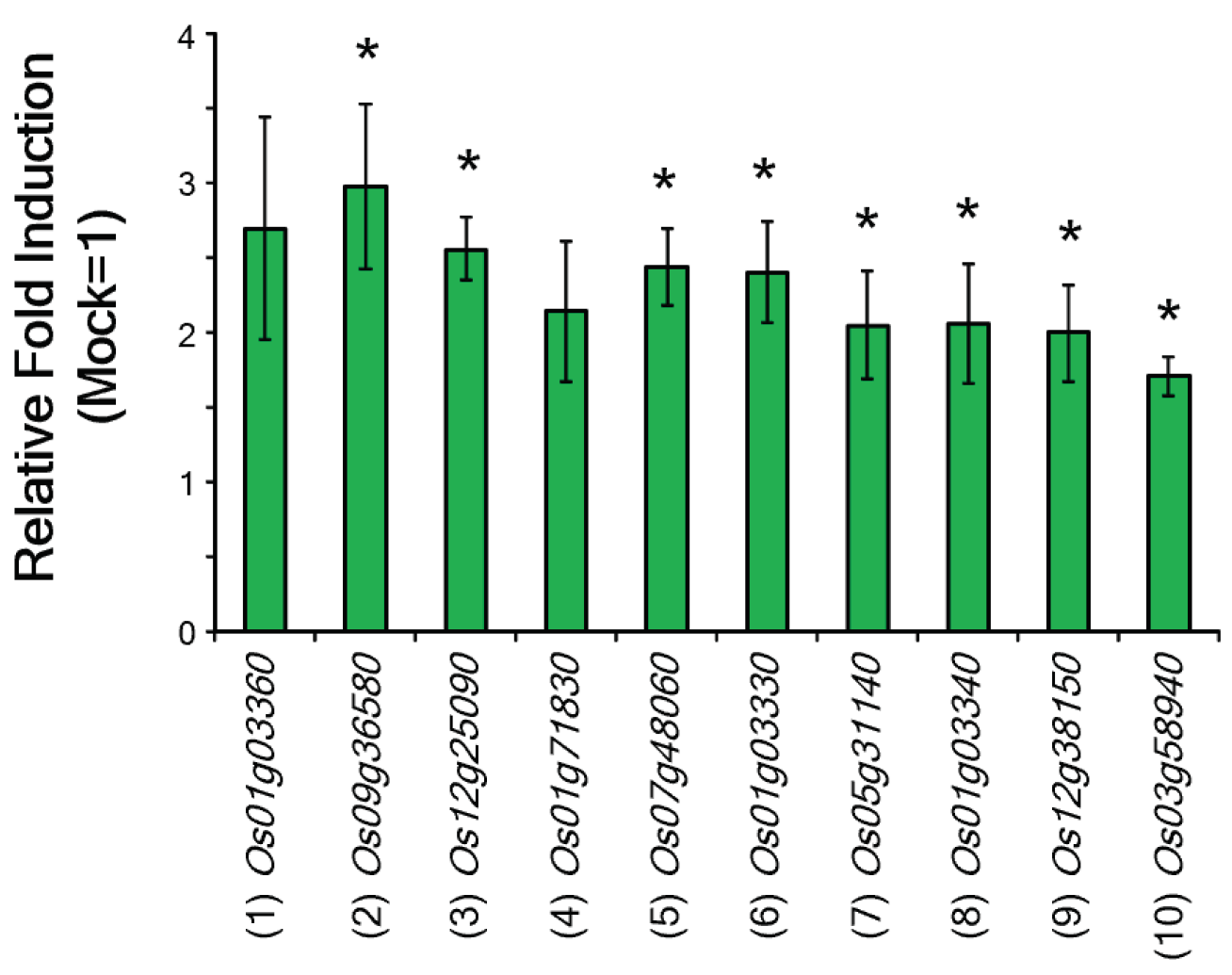
| (1) Os01g03360 | Bowman-Birk type bran trypsin inhibitor | 8.75 | <0.05 | This study |
| (2) Os09g36580 | Thaumatin-like protein 1 | 3.70 | <0.05 | This study |
| (3) Os12g25090 | Subtilisin/chymotrypsin inhibitor | 3.50 | <0.05 | This study |
| (4) Os01g71830 | Beta-1,3-glucanase | 3.44 | <0.05 | This study |
| (5) Os07g48060 | Peroxidase (OsPrx115) | 2.90 | <0.05 | This study |
| (6) Os01g03330 | Proteinase inhibitor I12 | 2.72 | <0.05 | This study |
| (7) Os05g31140 | Beta-glucanase | 2.55 | <0.05 | This study |
| (8) Os01g03340 | Bowman Birk trypsin inhibitor | 2.51 | <0.05 | This study |
| (9) Os12g38150 | Thaumatin | 2.25 | <0.05 | This study |
| (10) Os03g58940 | Lipid transfer protein | 2.01 | <0.05 | This study |
| ABA-responsive gene | ||||
| (11) Os06g48300 | Protein phosphatase 2C family protein | 0.20 | <0.05 | [28] |
| (12) Os05g46480 | LEA-like protein | 0.22 | >0.05 | [28] |
| (13) Os05g04700 | Similar to ICT protein | 0.48 | >0.05 | [28] |
| (14) Os08g36440 | Abscisic acid and stress inducible (A22) gene | 0.50 | >0.05 | This study |
| (15) Os01g64000 | Similar to ABA response element binding factor | 0.54 | >0.05 | This study |
| (16) Os11g26760 | Dehydrin RAB 16C | 0.60 | >0.05 | [28] |
| (17) Os12g29400 | GRAM domain containing protein | 0.71 | >0.05 | [28] |
| (18) Os06g10880 | Similar to ABA-responsive element binding protein 1 | 0.71 | >0.05 | This study |
| (19) Os11g26750 | Dehydrin RAB 16D | 0.81 | >0.05 | [28] |
| (20) Os02g44870 | Dehydrin family protein | 0.97 | >0.05 | [28] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taniguchi, S.; Takeda, A.; Kiryu, M.; Gomi, K. Jasmonic Acid-Induced β-Cyclocitral Confers Resistance to Bacterial Blight and Negatively Affects Abscisic Acid Biosynthesis in Rice. Int. J. Mol. Sci. 2023, 24, 1704. https://doi.org/10.3390/ijms24021704
Taniguchi S, Takeda A, Kiryu M, Gomi K. Jasmonic Acid-Induced β-Cyclocitral Confers Resistance to Bacterial Blight and Negatively Affects Abscisic Acid Biosynthesis in Rice. International Journal of Molecular Sciences. 2023; 24(2):1704. https://doi.org/10.3390/ijms24021704
Chicago/Turabian StyleTaniguchi, Shiduku, Aya Takeda, Masaki Kiryu, and Kenji Gomi. 2023. "Jasmonic Acid-Induced β-Cyclocitral Confers Resistance to Bacterial Blight and Negatively Affects Abscisic Acid Biosynthesis in Rice" International Journal of Molecular Sciences 24, no. 2: 1704. https://doi.org/10.3390/ijms24021704