

Glycoalkaloid Composition and Flavonoid Content as Driving Forces of Phytotoxicity in Diploid Potato
 , and
, and
Abstract
:1. Introduction
2. Results
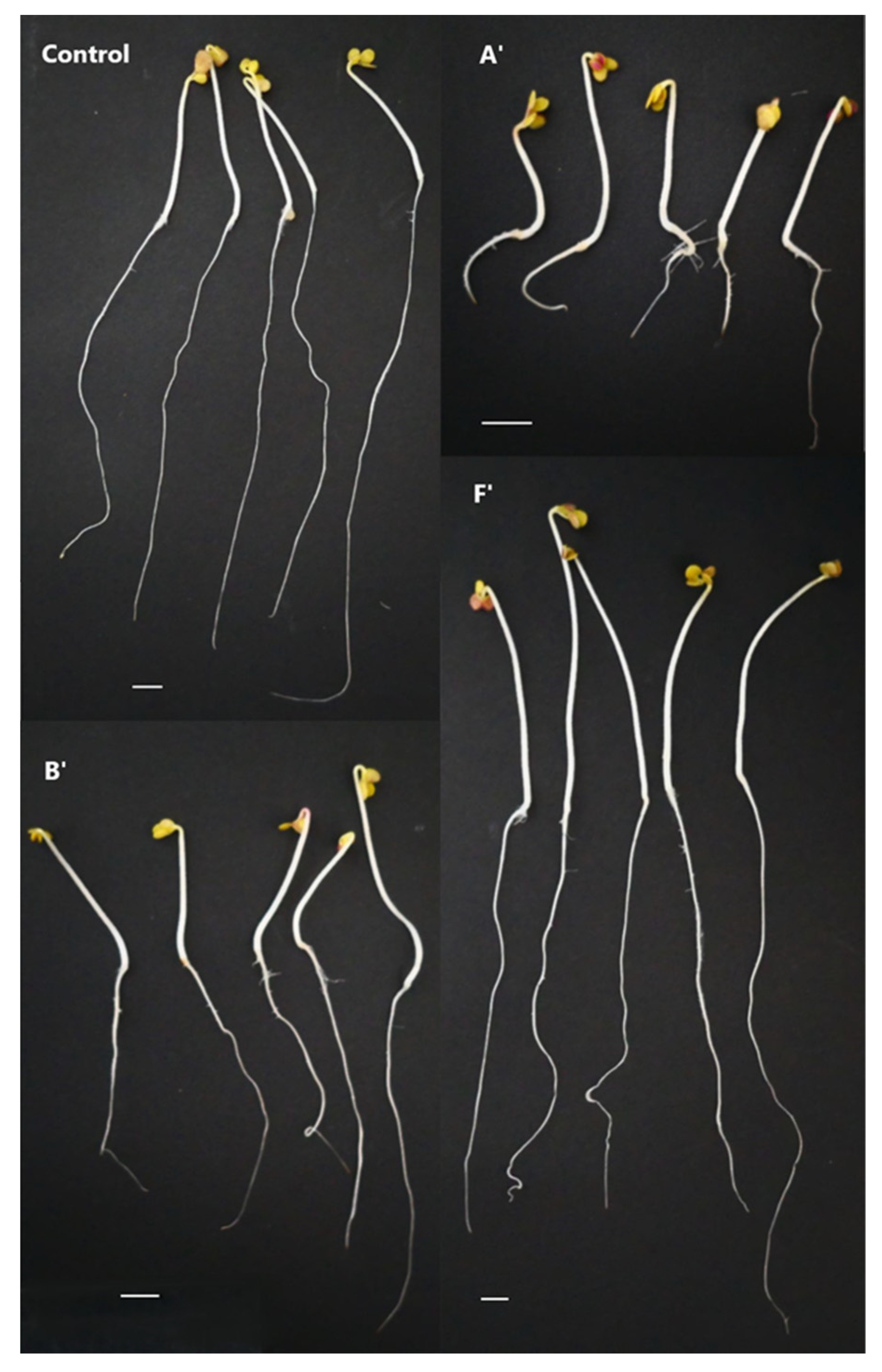
2.1. Evaluation of the Glycoalkaloid Content and the Phytotoxicity of Leaf Extracts from Individuals of the Potato Population
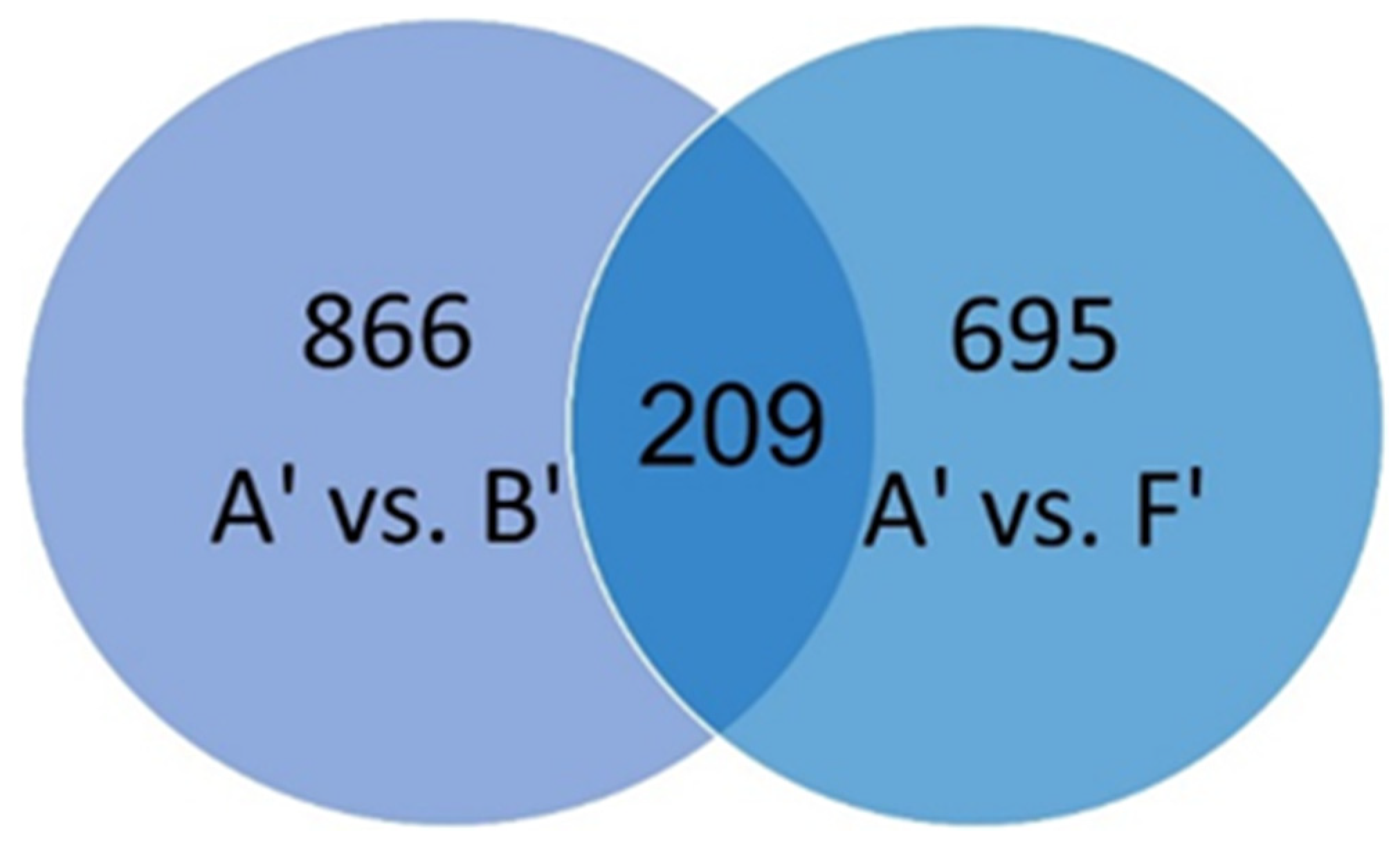
2.2. Profiles of Differentially Expressed Genes (DEGs)
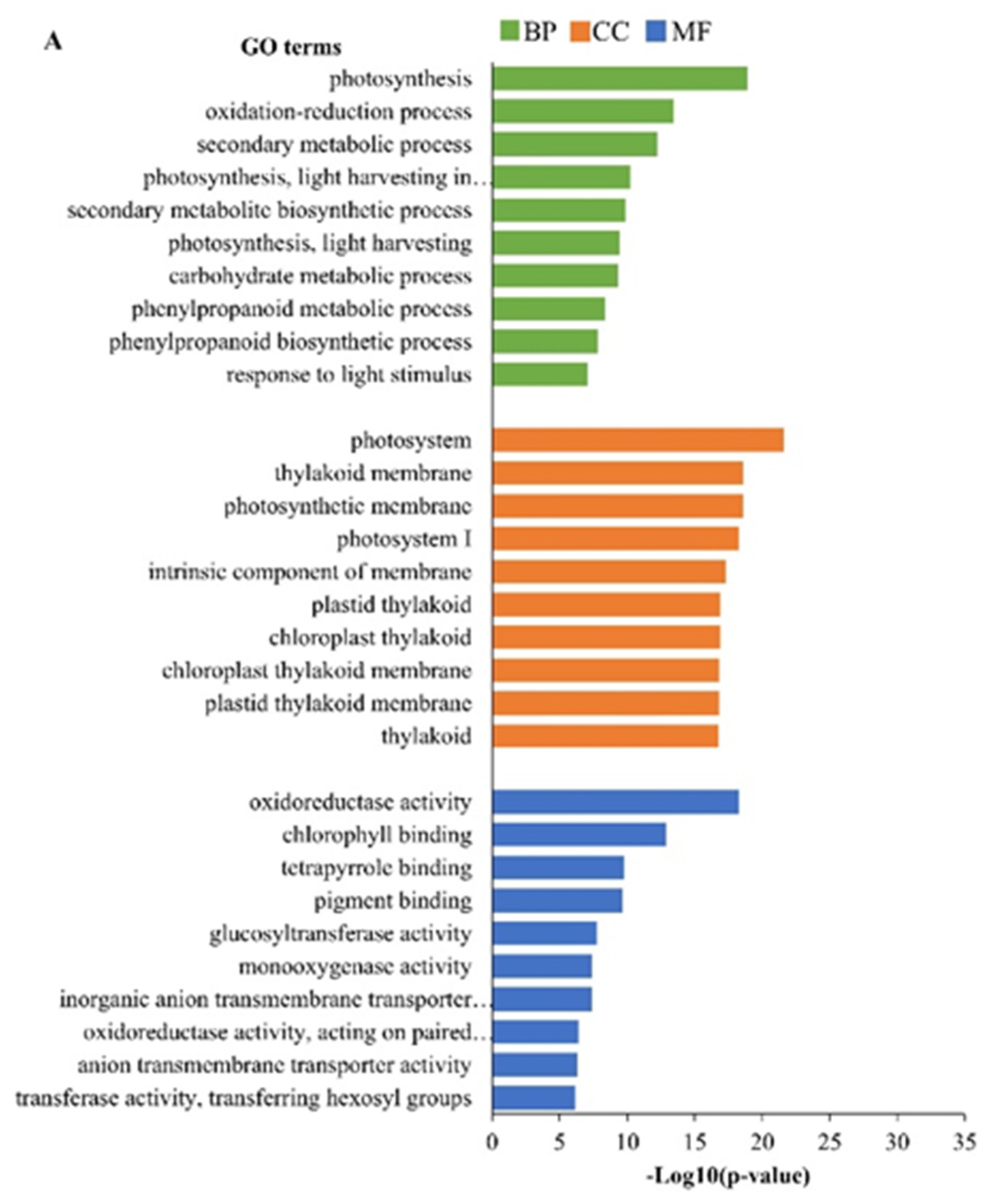
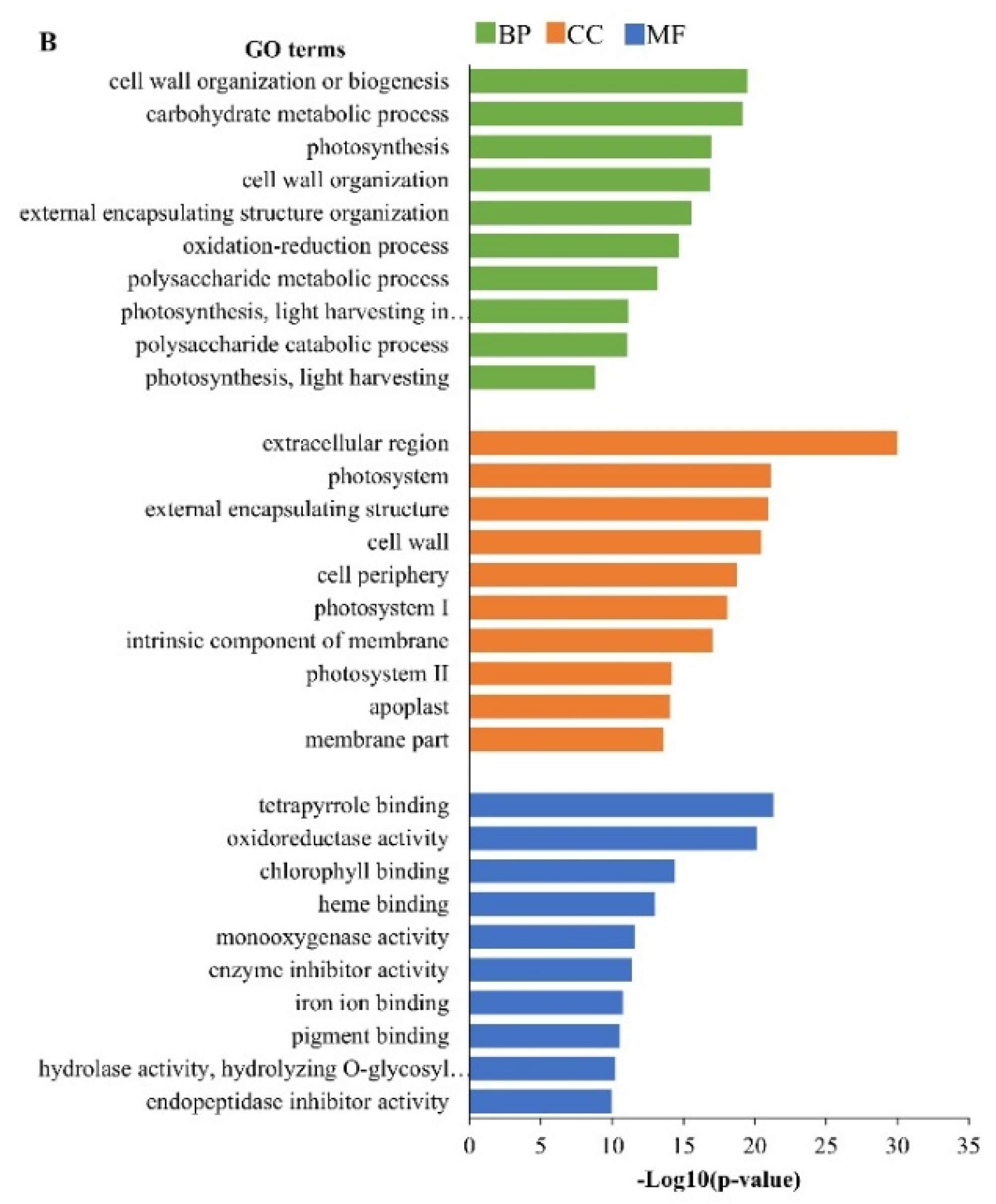
2.3. Gene Ontology (GO) Analysis of Differentially Expressed Genes (DEGs)
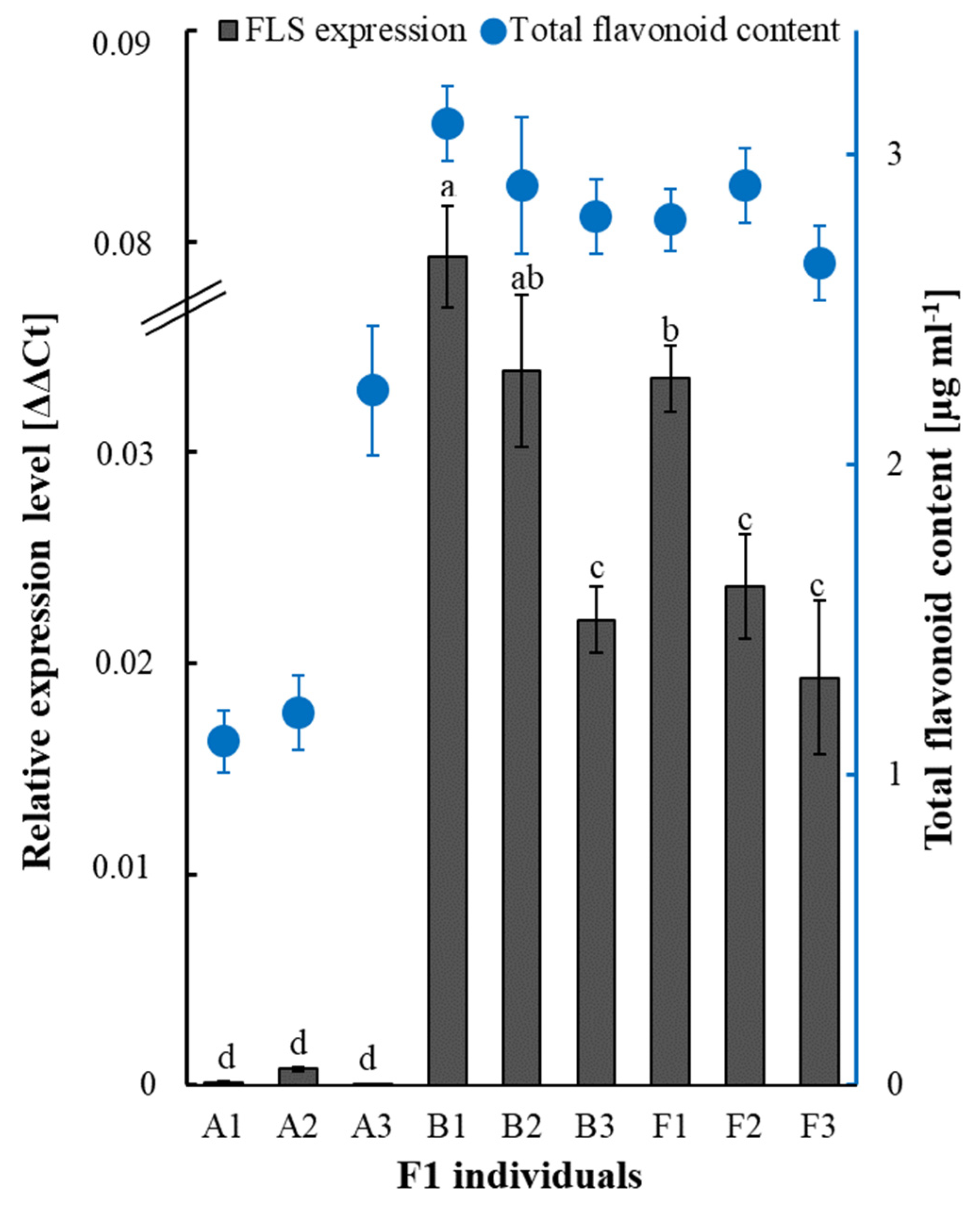
2.4. Flavonol Synthase/Flavanone 3-Hydroxylase-like (FLS) Expression and Total Flavonoid Content Analyzes
2.5. Glycoalkaloid Composition in Bulk Samples A’, B’, and F’
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Evaluation of The Phytotoxicity of Potato Leaf Extracts
4.3. Construction of Bulk Samples
4.4. RNA-Seq and GO Analyses of Bulk Samples
4.5. Expression Level of FLS and Total Flavonoid Content
4.6. Analysis of The Glycoalkaloid Composition in Potato Leaf Extracts
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inderjit; Nilsen, E.T. Bioassays and field studies for allelopathy in terrestrial plants: Progress and problems. Crit. Rev. Plant Sci. 2003, 22, 221–238. [Google Scholar] [CrossRef]
- Scavo, A.; Abbate, C.; Mauromicale, G. Plant allelochemicals: Agronomic, nutritional and ecological relevance in the soil system. Plant Soil 2019, 442, 23–48. [Google Scholar] [CrossRef]
- Trezzi, M.M.; Vidal, R.A.; Balbinot, A.A.; Von Hertwig Bittencourt, H.; Da Silva Souza Filho, A.P. Allelopathy: Driving mechanisms governing its activity in agriculture. J. Plant Interact. 2016, 11, 53–60. [Google Scholar] [CrossRef]
- Gniazdowska, A.; Bogatek, R. Allelopathic interactions between plants. Multi-site action of allelochemicals. Acta Physiol. Plant. 2005, 27, 395–407. [Google Scholar] [CrossRef]
- De Martino, L.; Mencherini, T.; Mancini, E.; Aquino, R.P.; De Almeida, L.F.; De Feo, V. In vitro phytotoxicity and antioxidant activity of selected flavonoids. Int. J. Mol. Sci. 2012, 13, 5406–5419. [Google Scholar] [CrossRef]
- De Albuquerque, M.B.; dos Santos, R.C.; Lima, L.M.; Melo Filho, D.A.; Nogueira, R.J.M.C.; da Cậmara, C.A.G.; de Rezende Ramos, A. Allelopathy, an alternative tool to improve cropping systems. A review. Agron. Sust. Developm. 2011, 31, 379–395. [Google Scholar] [CrossRef] [Green Version]
- Krasuska, U.; Andrzejczak, O.; Staszek, P.; Borucki, W.; Gniazdowska, A. Meta-Tyrosine induces modification of reactive nitrogen species level, protein nitration and nitrosoglutathione reductase in tomato roots. Nitric Oxide 2017, 68, 56–67. [Google Scholar] [CrossRef]
- Staszek, P.; Krasuska, U.; Otulak-Kozieł, K.; Fettke, J.; Gniazdowska, A. Canavanine-induced decrease in nitric oxide synthesis alters activity of antioxidant system but does not impact S-nitrosoglutathione catabolism in tomato roots. Front. Plant Sci. 2019, 10, 1077. [Google Scholar] [CrossRef] [Green Version]
- Staszek, P.; Krasuska, U.; Ciacka, K.; Gniazdowska, A. ROS metabolism perturbation as an element of mode of action of allelochemicals. Antioxidants 2021, 10, 1648. [Google Scholar] [CrossRef]
- Soltys, D.; Rudzińska-Langwald, A.; Gniazdowska, A.; Wiśniewska, A.; Bogatek, R. Inhibition of tomato (Solanum lycopersicum L.) root growth by cyanamide is due to altered cell division, phytohormone balance and expansin gene expression. Planta 2012, 236, 1629–1638. [Google Scholar] [CrossRef]
- Kaunda, J.S.; Zhang, Y.J. The genus Solanum: An ethnopharmacological, phytochemical and biological properties review. Nat. Prod. Bioprospect. 2019, 9, 77–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spooner, D.M.; Ghislain, M.; Simon, R.; Jansky, S.H.; Gavrilenko, T. Systematics, diversity, genetics, and evolution of wild and cultivated potatoes. Bot. Rev. 2014, 80, 283–383. [Google Scholar] [CrossRef]
- Mushtaq, W.; Siddiqui, M.B. Allelopathy in Solanaceae plants. J. Plant Prot. Res. 2018, 58, 1–7. [Google Scholar]
- Nelson, D.C.; Thoreson, M.C. Competition between Potatoes (Solanum tuberosum) and Weeds. Weed Sci. 1981, 29, 672–677. [Google Scholar] [CrossRef]
- Kumar, V.; Aulakh, C.S. Effect of planting geometry and potato seed tuber size on weeds and potato tuber yield. Ind. J. Weed Sci. 2022, 54, 291–295. [Google Scholar] [CrossRef]
- Jensen, P.H.; Harder, B.J.; Strobel, B.W.; Svensmark, B.; Hansen, H.C.B. Extraction and determination of the potato glycoalkaloid α-solanine in soil. Int. J. Environ. Anal. Chem. 2007, 87, 813–824. [Google Scholar] [CrossRef]
- Sołtys-Kalina, D.; Murawska, Z.; Strzelczyk-Żyta, D.; Wasilewicz-Flis, I.; Marczewski, W. Phytotoxic potential of cultivated and wild potato species (Solanum sp.): Role of glycoalkaloids, phenolics and flavonoids in phytotoxicity against mustard (Sinapis alba L.). Acta Physiol. Plant. 2019, 41, 55. [Google Scholar] [CrossRef] [Green Version]
- Szajko, K.; Ciekot, J.; Wasilewicz-Flis, I.; Marczewski, W.; Sołtys-Kalina, D. Transcriptional and proteomic insights into phytotoxic activity of interspecific potato hybrids with low glycoalkaloid contents. BMC Plant Biol. 2021, 21, 60. [Google Scholar] [CrossRef]
- Friedman, M.; McDonald, G.M.; Filadelfi-Keszi, M. Potato glycoalkaloids: Chemistry, analysis, safety, and plant physiology. Crit. Rev. Plant Sci. 1997, 16, 55–132. [Google Scholar] [CrossRef]
- Friedman, M. Potato glycoalkaloids and metabolites: Roles in the plant and in the diet. J. Agric. Food Chem. 2006, 54, 8655–8681. [Google Scholar] [CrossRef]
- Chung, I.M.; Ham, T.H.; Cho, G.W.; Kwon, S.W.; Lee, Y.; Seo, J.; An, Y.-J.; Kim, S.-Y.; Kim, S.-H.; Lee, J. Study of quantitative trait loci (QTLs) associated with the allelopathic trait in rice. Genes 2020, 11, 470. [Google Scholar] [CrossRef] [PubMed]
- Coleman, R.K.; Gill, G.S.; Rebetzke, G.J. Identification of quantitative trait loci for traits conferring weed competitiveness in wheat (Triticum aestivum L.). Aust. J. Agric. Res. 2001, 52, 1235–1246. [Google Scholar] [CrossRef]
- Zuo, S.; Liu, G.; Li, M. Genetic basis of allelopathic potential of winter wheat based on the perspective of quantitative trait locus. Field Crops Res. 2012, 135, 67–73. [Google Scholar] [CrossRef]
- Zhang, Q.; Lin, S.; Zhang, Q.; Wang, P.; Wang, H.; He, H. Comparative analysis on metabolites and gene expression difference of various allelopathic potential rice under weed stress. Weed Res. 2022, 62, 340–346. [Google Scholar] [CrossRef]
- Zhang, H.; Rutherford, S.; Qi, S.; Huang, P.; Dai, Z. Transcriptome profiling of Arabidopsis thaliana roots in response to allelopathic effects of Conyza canadensis. Ecotoxicology 2022, 31, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, Y.; Yuan, L.; Weber, E.; van Kleunen, M. Effect of allelopathy on plant performance: A meta-analysis. Ecol. Lett. 2021, 24, 348–362. [Google Scholar] [CrossRef]
- Jensen, P.H.; Strobel, B.W.; Hansen, H.C.B.; Jacobsen, O.S. Fate of toxic potato glycoalkaloids in a potato field. J. Agric. Food Chem. 2009, 57, 2862–2867. [Google Scholar] [CrossRef]
- Subrahmaniam, H.J.; Libourel, C.; Journet, E.P.; Morel, J.B.; Muños, S.; Niebel, A.; Raffaele, S.; Roux, F. The genetics underlying natural variation of plant-plant interactions, a beloved but forgotten member of the family of biotic interactions. Plant J. 2018, 93, 747–770. [Google Scholar] [CrossRef] [Green Version]
- Altamimi, A.S.; Souheila, N. Interspecific variations in the habitats of Rhus tripartitum L. populations in Saudi Arabia leading to changes in morphological traits and allelopathic activity. CERNE 2021, 27, e102993. [Google Scholar] [CrossRef]
- Mönchgesang, S.; Strehmel, N.; Schmidt, S.; Westphal, L.; Taruttis, F.; Müller, E.; Herklotz, S.; Neumann, S.; Scheel, D. Natural variation of root exudates in Arabidopsis thaliana-linking metabolomic and genomic data. Sci. Rep. 2016, 6, 29033. [Google Scholar]
- Peng, Z.; Wang, P.; Tang, D.; Shang, Y.; Li, C.-H.; Huang, S.-W.; Zhang, C.-Z. Inheritance of steroidal glycoalkaloids in potato tuber flesh. J. Integr. Agric. 2019, 18, 2255–2263. [Google Scholar] [CrossRef]
- Brooker, R.W.; Bennett, A.E.; Cong, W.F.; Daniell, T.J.; George, T.S.; Hallett, P.D.; Hawes, C.; Iannetta, P.P.M.; Jones, H.G.; Karley, A.J.; et al. Improving intercropping: A synthesis of research in agronomy, plant physiology and ecology. New Phytol. 2015, 206, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Wendling, M.; Büchi, L.; Amossé, C.; Jeangros, B.; Walter, A.; Charles, R. Specific interactions leading to transgressive overyielding in cover crop mixtures. Agric. Ecosyst. Environ. 2017, 241, 88–99. [Google Scholar] [CrossRef]
- Schilbert, H.M.; Schöne, M.; Baier, T.; Busche, M.; Viehöver, P.; Weisshaar, B.; Holtgräwe, D. Characterization of the Brassica napus flavonol synthase gene family reveals bifunctional flavonol synthases. Front. Plant Sci. 2021, 12, 733762. [Google Scholar] [CrossRef]
- Sun, F.; Li, S.; He, D.; Cao, G.; Ni, X.; Tai, G.; Zhou, Y.; Wang, D. Allelopathic effects of Solanum glycoalkaloids and their interactions. Phytochemistry 2010, 71, 1534–1538. [Google Scholar] [CrossRef]
- Bovy, A.; Schijlen, E.; Hall, R.D. Metabolic engineering of flavonoids in tomato (Solanum lycopersicum): The potential for metabolomics. Metabolomics 2007, 3, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Payyavula, R.S.; Navarre, D.A.; Kuhl, J.C.; Pantoja, A.; Pillai, S.S. Differential effects of environment on potato phenylpropanoid and carotenoid expression. BMC Plant Biol. 2012, 12, 39. [Google Scholar] [CrossRef] [Green Version]
- Alseekh, S.; Perez De Souza, L.; Benina, M.; Fernie, A.R. The style and substance of plant flavonoid decoration; towards defining both structure and function. Phytochemistry 2020, 174, 112347. [Google Scholar] [CrossRef]
- Gniazdowska, A.; Oracz, K.; Bogatek, R. Phytotoxic effect of sunflower (Helianthus annuus L.) to hormonal balance (ABA: Ethylene) in germinating mustard (Sinapis alba L.) seeds. Allelopath. J. 2007, 19, 215–226. [Google Scholar]
- Oracz, K.; Bailly, C.; Gniazdowska, A.; Come, D.; Corbineau, F.; Bogatek, R. Induction of oxidative stress by sunflower phytotoxins in germinating mustard seeds. J. Chem. Ecol. 2007, 33, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreu, A.; Oliva, C.; Distel, S.; Daleo, G. Production of phytoalexins, glycoalkaloids and phenolics in leaves and tubers of potato cultivars with different degrees of field resistance after infection with Phytophthora infestans. Am J Potato Res. 2001, 44, 1–9. [Google Scholar] [CrossRef]
- Zou, C.; Wang, P.; Xu, Y. Bulked sample analysis in genetics, genomics and crop improvement. Plant Biotechnol. J. 2016, 14, 1941–1955. [Google Scholar] [CrossRef] [PubMed]
- Chomczyński, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Nahar, N.; Westerberg, E.; Arif, U.; Huchelmann, A.; Olarte Guasca, A.; Beste, L.; Dalman, K.; Dutta, P.C.; Jonsson, L.; Sitbon, F. Transcript profiling of two potato cultivars during glycoalkaloid-inducing treatments shows differential expression of genes in sterol and glycoalkaloid metabolism. Sci. Rep. 2017, 7, 43268. [Google Scholar] [CrossRef]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Average | Minimum | Maximum | ±SD | Trait Distribution 1 | |
|---|---|---|---|---|---|---|
| W Value | p | |||||
| Glycoalkaloid content [μg mL−1] | 10.764 | 1.063 | 46.653 | 10.943 | 0.806 | <0.000 |
| Phytotoxicity * [%] | 86.743 | 41.618 | 140.227 | 28.283 | 0.934 | <0.000 |
| Group | Glycoalkaloid Content 1 | Phytotoxicity | Number of F1 Individuals (Frequency) |
|---|---|---|---|
| A | High | High | 5 (4%) |
| B | High | Low | 8 (7%) |
| C | Low | High | 50 (44%) |
| D | Low | Low | 10 (9%) |
| E | Low | Growth stimulation | 30 (27%) |
| F | High | Growth stimulation | 10 (9%) |
| Locus | Log2FC | p-Value 2 | Product Name |
|---|---|---|---|
| A’1 vs. B’ | |||
| upregulated DEGs | |||
| LOC102585933 | 8.30 | 9.63 × 10−36 | flavonol synthase/flavanone 3-hydroxylase-like |
| LOC102594045 | 7.96 | 1.27 × 10−10 | UDP-glycosyltransferase 71E1-like |
| LOC102584518 | 7.05 | 2.60 × 10−25 | metal transporter Nramp5-like |
| LOC107057710 | 6.93 | 1.12 × 10−7 | uncharacterized protein At1g18380-like |
| LOC102579990 | 6.87 | 1.04 × 10−7 | uncharacterized protein LOC102579990 |
| LOC107057684 | 6.18 | 5.55 × 10−5 | G-type lectin S-receptor-like serine/threonine-protein kinase At4g27290 |
| LOC102590804 | 6.16 | 2.38 × 10−6 | nodulation receptor kinase-like |
| LOC102590432 | 6.13 | 1.54 × 10−6 | probable L-type lectin-domain containing receptor kinase S.5 |
| LOC102596031 | 6.08 | 1.24 × 10−21 | uncharacterized LOC102601567. transcript variant X2 |
| LOC102601567 | 6.08 | 1.15 × 10−14 | uncharacterized protein At1g28695-like |
| downregulated DEGs | |||
| LOC107058371 | −10.00 | 4.61 × 10−15 | chitin-binding lectin 1-like |
| LOC102605914 | −8.96 | 7.09 × 10−12 | putative UPF0481 protein At3g02645 |
| LOC102579908 | −8.39 | 4.85 × 10−10 | uncharacterized protein LOC102579908 |
| LOC102603653 | −8.21 | 8.76 × 10−10 | WD repeat-containing protein 48-like. transcript variant X2 |
| LOC102594853 | −8.20 | 3.90 × 10−43 | Werner Syndrome-like exonuclease |
| LOC102596193 | −8.13 | 5.76 × 10−10 | uncharacterized protein LOC102596193 |
| LOC102606330 | −8.06 | 7.35 × 10−25 | - |
| LOC102588561 | −7.83 | 1.01 × 10−8 | - |
| LOC102599813 | −7.51 | 7.20 × 10−8 | 2-isopropylmalate synthase A-like |
| LOC102578236 | −7.50 | 3.94 × 10−8 | - |
| A’ vs. F’ | |||
| upregulated DEGs | |||
| LOC102605504 | 7.07 | 6.44 × 10−8 | probable trans−2-enoyl-CoA reductase. mitochondrial |
| LOC107061996 | 6.72 | 2.67 × 10−7 | zeatin O-glucosyltransferase-like |
| LOC102585933 | 6.40 | 1.27 × 10−20 | flavonol synthase/flavanone 3-hydroxylase-like |
| LOC102584318 | 6.38 | 1.69 × 10−6 | uncharacterized LOC102584318. transcript variant X3 |
| LOC102597028 | 6.31 | 4.30 × 10−6 | uncharacterized protein LOC102597028 |
| LOC107057684 | 6.18 | 8.19 × 10−6 | G-type lectin S-receptor-like serine/threonine-protein kinase At4g27290 |
| LOC102582531 | 6.08 | 5.30 × 10−21 | replication protein A 70 kDa DNA-binding subunit B-like. transcript variant X11 |
| LOC102579990 | 6.06 | 1.02 × 10−5 | uncharacterized protein LOC102579990 |
| LOC102591096 | 5.97 | 4.58 × 10−6 | probable S-adenosylmethionine-dependent methyltransferase At5g38100 |
| LOC102584503 | 5.90 | 3.28 × 10−14 | small heat shock protein. chloroplastic |
| downregulated DEGs | |||
| LOC102602896 | −9.85 | 1.48 × 10−14 | cytochrome P450 72A15-like |
| LOC102606330 | −9.83 | 9.54 × 10−15 | - |
| LOC102578543 | −8.42 | 3.99 × 10−14 | salicylate carboxymethyltransferase-like |
| LOC102603573 | −7.84 | 1.12 × 10−8 | uncharacterized LOC102603573. transcript variant X5 |
| LOC102588561 | −7.70 | 1.87 × 10−8 | - |
| LOC107060242 | −7.15 | 5.69 × 10−7 | uncharacterized LOC107060242. transcript variant X2 |
| LOC102584808 | −7.05 | 9.08 × 10−10 | intracellular ribonuclease LX-like |
| LOC102577909 | −6.38 | 1.29 × 10−14 | cysteine protease inhibitor 8-like |
| LOC107058413 | −6.24 | 7.30 × 10−5 | uncharacterized LOC107058413. transcript variant X1 |
| LOC102595577 | −6.22 | 1.80 × 10−5 | cation/H(+) antiporter 18-like |
| Glycoalkaloids Frequency [%] | ||||||
|---|---|---|---|---|---|---|
| Bulk | Leptinine I | Leptine II | Solasonine | Solamargine | α-Solanine | α-Chaconine |
| A’ | 2.8 | 5.0 | 4.2 | 8.0 | 27.2 | 52.8 |
| B’ | 8.3 | - | - | - | 34.0 | 57.7 |
| F’ | 6.6 | - | - | - | 35.5 | 57.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szajko, K.; Smyda-Dajmund, P.; Ciekot, J.; Marczewski, W.; Sołtys-Kalina, D. Glycoalkaloid Composition and Flavonoid Content as Driving Forces of Phytotoxicity in Diploid Potato. Int. J. Mol. Sci. 2023, 24, 1657. https://doi.org/10.3390/ijms24021657
Szajko K, Smyda-Dajmund P, Ciekot J, Marczewski W, Sołtys-Kalina D. Glycoalkaloid Composition and Flavonoid Content as Driving Forces of Phytotoxicity in Diploid Potato. International Journal of Molecular Sciences. 2023; 24(2):1657. https://doi.org/10.3390/ijms24021657
Chicago/Turabian StyleSzajko, Katarzyna, Paulina Smyda-Dajmund, Jarosław Ciekot, Waldemar Marczewski, and Dorota Sołtys-Kalina. 2023. "Glycoalkaloid Composition and Flavonoid Content as Driving Forces of Phytotoxicity in Diploid Potato" International Journal of Molecular Sciences 24, no. 2: 1657. https://doi.org/10.3390/ijms24021657