

Estrogenic and Non-Estrogenic Disruptor Effect of Zearalenone on Male Reproduction: A Review
 ,
,
Abstract
:1. Introduction
2. Occurrence of Zearalenone in Food and Feed
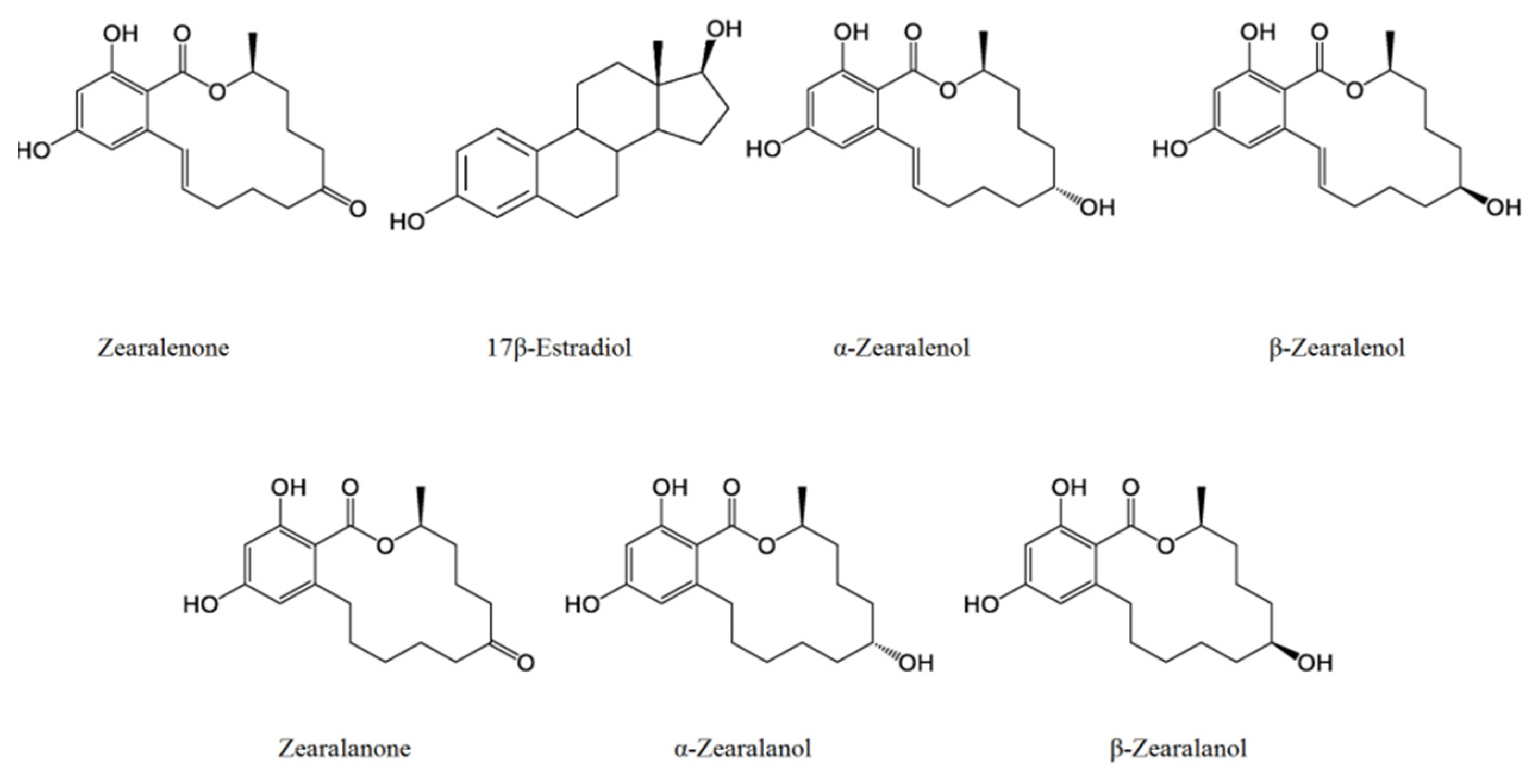
3. Chemical Properties
4. Overview of the Pathogenesis of Zearalenone
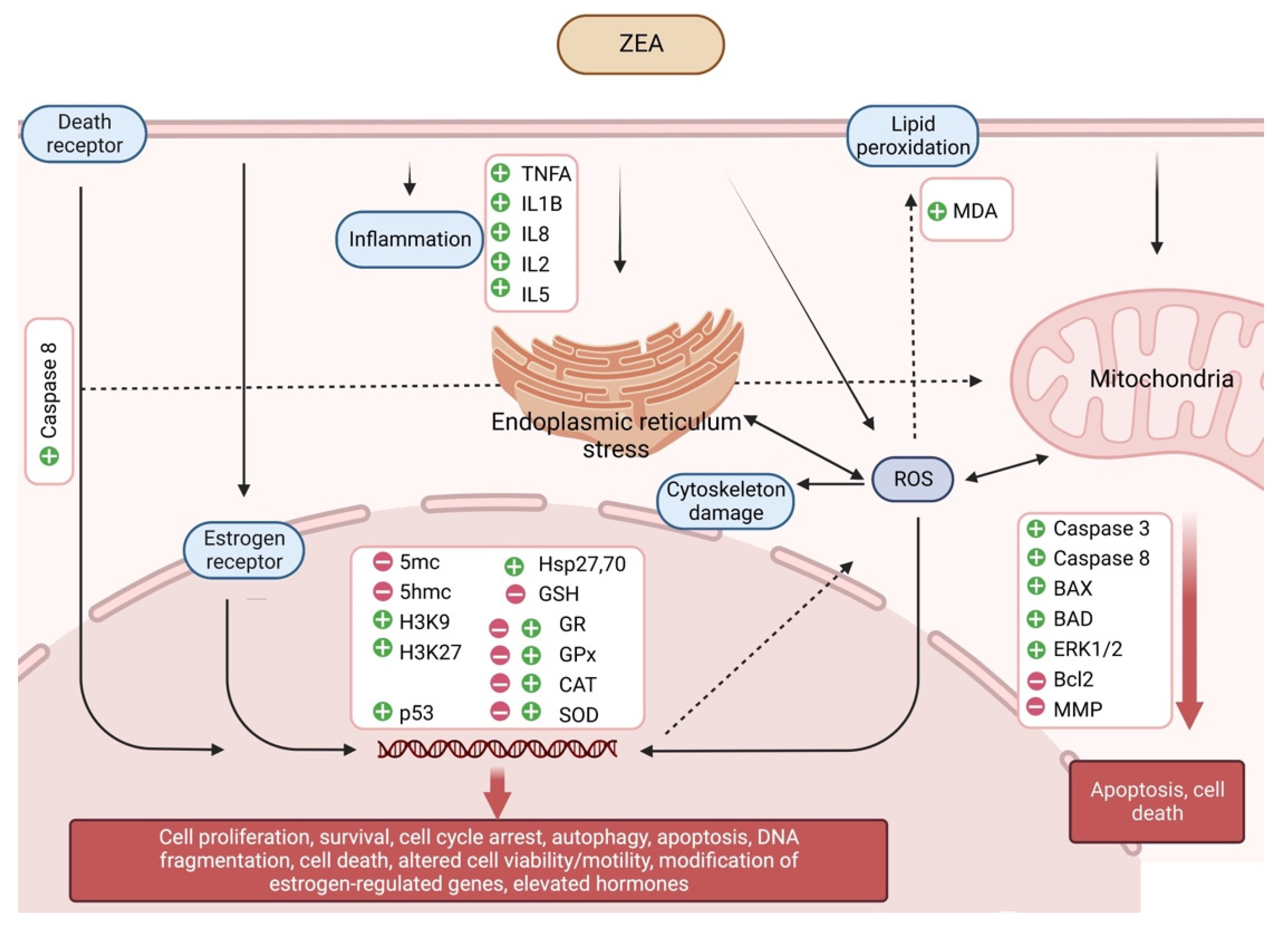
4.1. Estrogenic Effects
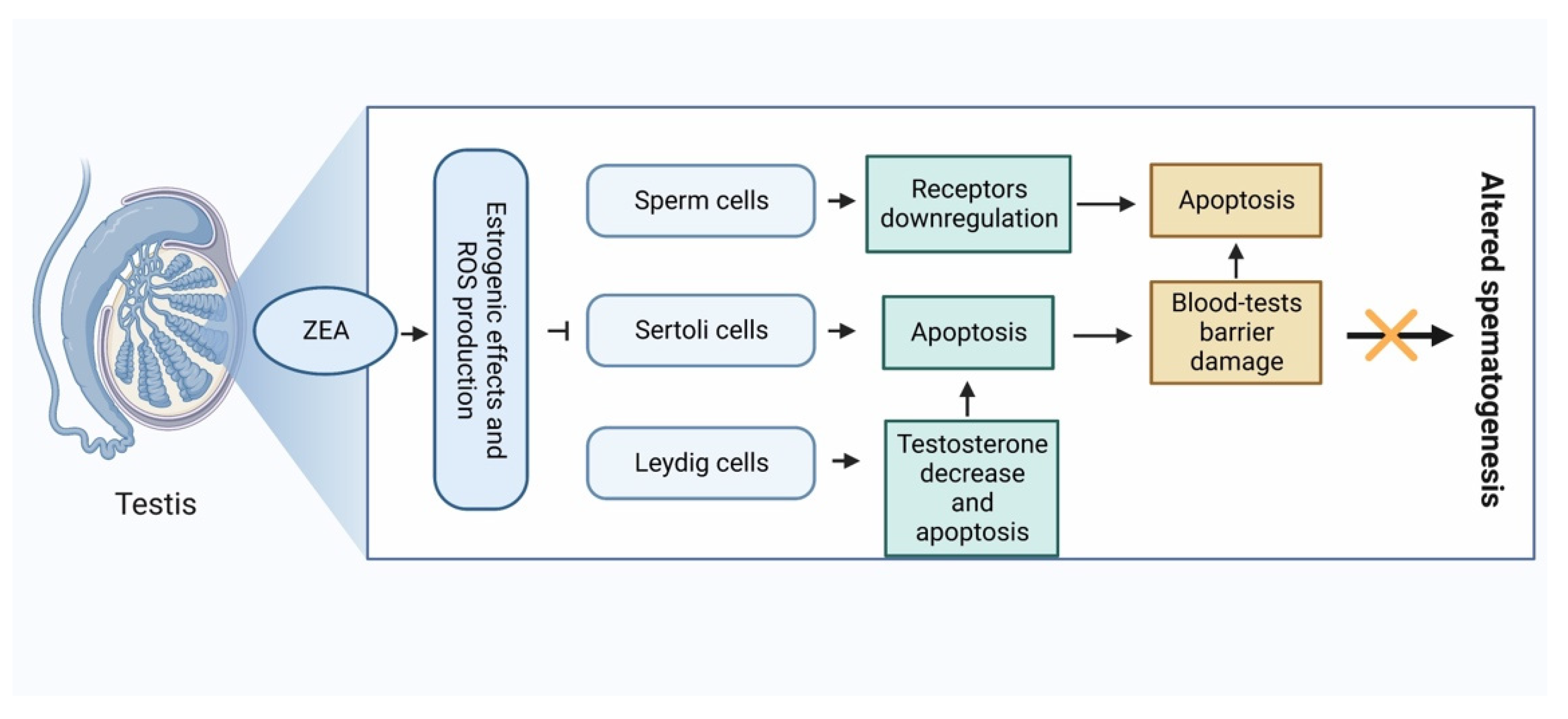
4.2. Non-Estrogenic Effects

{kind=link}
{kind=link}
{kind=link}
| First Author | Reference Number | Species | In Vitro/In Vivo | Method (Oral, ip, etc.) | Dose | Exposure Period | Results |
|---|---|---|---|---|---|---|---|
| Benzoni | [34] | Swine sperm | In vitro | Single exposure | 10−8–20 μM ZEA, α-zearalenol and β-zearalenol | 24 h and 48 h | Reduced cell viability and motility, increased rate of apoptosis and DNA fragmentation. |
| Bielas | [35] | Wild boars | In vivo | (i) Orally once a day for 7 days in every 2 months, (ii) orally once a day | (i) 150 μg/kg bw and (ii) 50 μg/kg bw ZEA | 1 y | Reduced sperm movement parameters. |
| Chao | [36] | Sertoli cells | In vitro | Single exposure | 0–50 μM ZEA | 24 h and 48 h | Reduced cell viability, increased rate of apoptosis and ROS production, increased expression of γH2AX and RAD51 DNA repairing enzymes, decrement in the expression of occludin and connexin 43, proteins that are present in the testis–blood barrier and gap junctions of Sertoli cells, respectively. |
| Chen | [37] | Mouse Leydig (TM3) cells | In vitro | Single exposure | 50 μM/L ZEA | 24 h | Increased rate of apoptosis via PTEN, inhibition of PI3K/AKT signal pathway. |
| Filannino | [38] | Stallion sperm | In vitro | Single exposure | 1 pM–0.1 mM ZEA, α-zearalenol and β-zearalenol | 2 h | Hyperactivated motility of equine sperm cells, premature completion of the acrosome reaction and diminished sperm physiology. The α form of zearalenol still possessed the estrogenic ability to induce hyperactivated motility, whereas its β stereo-isomere had lost this property. |
| Krejcárková | [40] | Pig sperm | In vitro | Single exposure | 0–20 μM ZEA and α-zearalenol | 2 h and 4 h | Inhibitory effects of ZEA concentrations above 5 μM on sperm motility. |
| Savard | [45] | Immature Sertoli TM4 cell line | In vitro | Single exposure | 0–100 μM ZEA | 24 h | Activation of MAPK signaling pathway, increased ROS formation, ZEA could be detrimental to the early steps of Sertoli cell differentiation. |
| Tassis | [47] | Boar semen | In vitro | Single exposure | 62.8 μM ZEA | 4 h | Reduced cell viability and increased rate of cellular abnormalities. |
| Wang | [48] | Rat Leydig cells | In vitro | Single exposure | 0–20 μg/mL ZEA | 12 h | Inhibition of cell proliferation and increased rate of apoptosis, increased Bax expression and cytochrome c release, activation of Caspase-3 and Caspase-9, cleavage of PARP, upregulation of LC3-II and Beclin-1. |
| Tsakmakidis | [89] | Pietrain boar semen | In vitro | Single exposure | 0–250 μm ZEA | 4 h | Decreased sperm motility and viability, disrupted acrosome reaction. |
| Zheng | [92] | Rat Leydig cells | In vitro | Single exposure | 0–20 μg/mL ZEA | 24 h | Disruption of α-tubulin filaments and F-actin bundles, and damage to the nucleus of Sertoli cells. Decrease in the levels of inhibin-β and transferrin in the cultural supernatants. |
| Xu | [93] | Sprague Dawley rats Sertoli cells | In vitro | Single exposure | 0–100 μM ZEA | 24 h | Increased ratio of Bax/Bcl-2and expressions of FasL, caspases-3, 28, and 29. Reduced proliferation of Sertoli cells, induction of apoptosis and necrosis in cells via extrinsic and intrinsic apoptotic pathways. |
| Abid-Essefi | [109] | Vero and Caco-2 cells | In vitro | Single exposure | 0–40 μM ZEA | 24 h | Reduces cell viability correlated to cell cycle perturbation, inhibited protein and DNA syntheses and increased MDA formation. |
| Gazzah | [110] | HepG2 cells | In vitro | Single exposure | 0–100 µM ZEA | 60 h | Induction of Hsp70 protein, increased ROS formation, DNA fragmentation and cell-cycle arrest. Increased Bax expression, decreased Bcl-2 expression and mitochondrial membrane potential, increase cytochrome c release, activation of caspase-3 and caspase-9. |
| Kouadio | [112] | Caco-2 cells | In vitro | Single exposure | 0–150 μM ZEA, deoxynivalenol and fumonisin B1 | 72 h | The three mycotoxins inducing lipid peroxidation (MDA production) in potency order fumonisin B1 > deoxynivalenol > ZEA. This effect seems to be related to their common target that is the mitochondria. Deoxynivalenol and ZEA also adversely affect lysosomes in contrast to fumonisin B1. The three mycotoxins inhibit protein synthesis. DNA synthesis seems to be restored for fumonisin B1 and deoxynivalenol suggesting a promoter activity. |
| Ouanes | [114] | Vero cells and Balb/c male mice | In vitro and in vivo | Single exposure | 0–20 μM and 0–40 mg 6 kg bw ZEA | 24 h and 48 h | ZEA induce micronuclei in cultured Vero cells as well as in mouse bone marrow cells. Vitamin E was found to prevent these toxic effects, most likely acting either as a structural analogue of ZEA or as an antioxidant. |
| Bouaziz | [116] | HepG2 cells | In vitro | Single exposure | 0–120 µM ZEA, 0–60 nM T-2 toxin, 0–100 µM ochratoxin | 0–48 h | The three mycotoxins-induced a caspase-dependent mitochondrial apoptotic pathway. The mitochondrial alterations include: bax relocalisation into the mitochondrial outer membrane, loss of the mitochondrial transmembrane potential, PTPC opening, and cytochrome c release. In the presence of ZEA and T-2 toxin, ROS level was highly increased at an early stage even before mitochondrial alterations were observed. |
| Ayed-Boussema | [117] | HepG2 cells | In vitro | Single exposure | 0–220 μM ZEA | 24 h | Reduces cell proliferation, induced an upregulation of ATM and p53 genes family accompanied by an upregulation of GADD45 to arrest the cell cycle and to allow the repair mechanisms to take place. Increased the ratio of pro-apoptotic factors/anti-apoptotic factors which led to the loss of mitochondrial potential, Bax translocation and cytochrome c release. |
| Lee | [118] | GC-1 spermatogonia cells | In vitro | Single exposure | 0–100 μM ZEA | 24 h | The cleaved caspase-3 and -8, Bad, Bax, and phosphorylation of p53 and ERK1/2, were increased. Expression levels of the autophagy-related genes Atg5, Atg3, Beclin 1, LC3, Ulk1, Bnip 3, and p62 were higher. The protein levels of both LC3A/B and Atg12 were remarkably increased. ZEA has toxic effects on tGC-1 spermatogonia cells and induces both apoptosis and autophagy. |
| Hassen | [120] | HepG2 cells | In vitro | Single exposure | 0–350 μM ZEA | 3 h and 24 h | Reduced cell viability, increased rate of DNA breaks, elevated Hsp70 and 90, reduced total GSH in cells. |
| Abid-Essefi | [121] | Caco-2 cells | In vitro | Single exposure | 0–100 μM ZEA, α-zearalenol and β-zearalenol | 48 h | Reduced cell viability, increased MDA level, increased DNA fragmentation, increased Caspase-3 activity and decreased Bcl-2 level. |
| Abid-Essefi | [122] | Vero cells | In vitro | Single exposure | 0–120 μM ZEA | 24 h | Reduced cell viability, increased ROS production, increased CAT activity and DNA damage. Reduced negative effects after co-treatment of an antioxidant. |
| Bennour | [123] | HepG2 and Vero cells | In vitro | Single exposure | 0–120 μM ZEA | 0–40 h | Inhibited cell proliferation, induction of Hsp70 and 27, increased ROS production, antioxidants protect against ZEA. |
| Stadnik | [124] | Liver cells of male Wistar rats | In vivo | Orally once a day | 50–500 μg/kg bw ZEA | 10 d | Reduced activities of SOD and GPx. |
| Othmen | [125] | Vero cells | In vitro | Single exposure | 0–180 μM α-zearalenol and β-zearalenol | 24 h | Increased MDA level, inhibited protein and DNA synthesis, elevated Hsp27 and 70. |
| Mike | [127] | Schizosaccharomyces pombe | In vitro | Single exposure | 0–500 μM ZEA | 1 h | Decreased activities of GSH, GPx and G6PD, increased activities of SOD, CAT, GR and GST, increased ROS production, DNA fragmentation and cell cycle arrest. |
| Fan | [128] | IPEC-J2 | In vitro | Single exposure | 0–8 μg/mL ZEA | 2 h and 48 h | Increased MDA level and ROS production, decreased GSH, CAT and SOD activities, decreased mitochondrial membrane potential. |
| Liu | [129] | Jejunal tissue of pregnant Sprague-Dawley rats | In vivo | Orally once a day | 0–146 mg/kg bw ZEA | Gestation day 1 to 7 | Increased MDA and GPx levels, decreased SOD and CAT activities. |
| Salah-Abbés | [130] | Balb/c mice | In vivo | Orally once a day | 40 mg/kg bw ZEA | 28 d | Increased MDA level, decreased GPx, CAT and SOD activities, reduced testis, seminal vesicle and prostate weights, diminished sperm count and motility. |
| Boeira | [131] | Liver, kidney and testis of Swiss albino mice | In vivo | Single exposure, orally | 40 mg/kg bw ZEA | 48 h | Increased CAT and SOD activities, reduced GST activity. |
| Fu | [132] | Bovine mammary epithelial cell line MAC-T | In vitro | Single exposure | 0–100 μM ZEA | 24 h and 48 h | Reduced cell viability, increased ROS production, reduced mitochondrial membrane potential, altered expression of endoplasmic reticulum stress-related genes (Grp78, Hsp70, Eif2a, Atf6, Ask1 and Chop). |
| Kim | [133] | Male Sprague-Dawley rats | In vivo | Single exposure, i.p. | 5 mg/kg bw ZEA | 0–48 h | Germ cell degeneration and apoptosis. |
| Elweza | [134] | Bovine endometrial epithelial cells | In vitro | Single exposure | 0–1000 ng/mL ZEA | 24 h | Increased expression of inflammatory cytokines TNFA, IL1B and chemokine IL8, altered sperm-uterine crosstalk, reduced sperm motility. |
| Cai | [136] | Sertoli cells | In vitro | Single exposure | 0–20 μM/L | 24 h | Increased rate of apoptotic cells, decreased expression of Bcl-2, increased expression of Bax, tBID, Fas, FasL, FADD, Caspase-8 and Caspase-9, increased release of cytochrome c. |
5. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [Green Version]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chemic. Soc. 2010, 15, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.C.; Mostrom, M.S.; Evans, T.J. Chapter 76—Zearalenone. In Veterinary Toxicology, 3rd ed.; Gupta, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 1055–1063. [Google Scholar]
- Ropejko, K.; Twarużek, M. Zearalenone and Its Metabolites-General Overview, Occurrence, and Toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- Omar, H.E.-D.M. Mycotoxins-induced oxidative stress and disease. In Mycotoxin and Food Safety in Developing Countries, 1st ed.; Makun, H.A., Ed.; InTech: Rijeka, Croatia, 2013. [Google Scholar] [CrossRef] [Green Version]
- Mirocha, C.J.; Pathre, S.V.; Robison, T.S. Comparative metabolism of zearalenone and transmission into bovine milk. Food Cosmet. Toxicol. 1981, 19, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Prelusky, D.B.; Scott, P.M.; Trenholm, H.L.; Lawrence, G.A. Minimal transmission of zearalenone to milk of dairy cows. J. Environ. Sci. Health B 1990, 25, 87–103. [Google Scholar] [CrossRef]
- Coffey, R.; Cummins, E.; Ward, S. Exposure assessment of mycotoxins in dairy milk. Food Control. 2009, 20, 239–249. [Google Scholar] [CrossRef]
- Kuiper-Goodman, T.; Scott, P.M.; Watanabe, H. Risk assessment of the mycotoxin zearalenone. Regul Toxicol Pharmacol. 1987, 7, 253–306. [Google Scholar] [CrossRef]
- Schwake-Anduschus, C.; Proske, M.; Sciurba, E.; Muenzing, K.; Koch, M.; Maul, R. Distribution of deoxynivalenol, zearalenone, and their respective modified analogues in milling fractions of naturally contaminated wheat grains. World Mycotoxin J. 2015, 8, 433–443. [Google Scholar] [CrossRef]
- Bennett, G.A.; Peplinski, A.J.; Brekke, O.L.; Jackson, L.K.; Wichser, W.R. Zearalenone—distribution in dry-milled fractions of contaminated corn. Cereal Chem. 1976, 53, 299–307. [Google Scholar]
- Bories, G.F.; Perdu-Durand, E.F.; Sutra, J.F.; Tulliez, J.E. Evidence for glucuronidation and sulfation of zeranol and metabolites (taleranol and zearalanone) by rat and pig hepatic subfractions. Drug Metab. Dispos. 1991, 19, 140–143. [Google Scholar]
- Gromadzka, K.; Waskiewicz, A.; Chelkowski, J.; Golinski, P. Zearalenone and its metabolites: Occurrence, detection, toxicity and guidelines. World Mycotoxin J. 2008, 1, 209–220. [Google Scholar] [CrossRef]
- Peillod, C.; Laborde, M.; Travel, A.; Mika, A.; Bailly, J.D.; Cleva, D.; Boissieu, C.; Le Guennec, J.; Albaric, O.; Labrut, S.; et al. Toxic Effects of Fumonisins, Deoxynivalenol and Zearalenone Alone and in Combination in Ducks Fed the Maximum EUTolerated Level. Toxins 2021, 13, 152. [Google Scholar] [CrossRef] [PubMed]
- Gruber-Dorninger, C.; Faas, J.; Doupovec, B.; Aleschko, M.; Stoiber, C.; Höbartner-Gußl, A.; Schöndorfer, K.; Killinger, M.; Zebeli, Q.; Schatzmayr, D. Metabolism of Zearalenone in the Rumen of Dairy Cows with and without Application of a Zearalenone-Degrading Enzyme. Toxins 2021, 13, 84. [Google Scholar] [CrossRef] [PubMed]
- Al-Jaal, B.; Latiff, A.; Salama, S.; Hussain, H.M.; Al-Thani, N.A.; Al-Naimi, N.; Al-Qasmi, N.; Horvatovich, P.; Jaganjac, M. Analysis of Multiple Mycotoxins in the Qatari Population and Their Relation to Markers of Oxidative Stress. Toxins 2021, 13, 267. [Google Scholar] [CrossRef]
- Yan, Z.; Wang, L.; Wang, J.; Tan, Y.; Yu, D.; Chang, X.; Fan, Y.; Zhao, D.; Wang, C.; De Boevre, M.; et al. A QuEChERS-Based Liquid Chromatography-Tandem Mass Spectrometry Method for the Simultaneous Determination of Nine Zearalenone-Like Mycotoxins in Pigs. Toxins 2018, 10, 129. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Huangfu, B.; Xu, T.; Xu, W.; Asakiya, C.; Huang, K.; He, X. Research Progress of Safety of Zearalenone: A Review. Toxins 2022, 14, 386. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxifcation, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- European Commission Commission recommendation (2006/576/EU) of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union. 2006, 49, 7–9.
- European Commission Commission regulation (1126/2007) of 28 September 2007 amending Regulation (1881/2006) on setting maximum levels for certain contaminants in foodstuffs as regards Fusarium toxins in maize and maize products. Off. J. Eur. Union. 2007, 50, 14–17.
- Minervini, F.; Giannoccaro, A.; Cavallini, A.; Visconti, A. Investigations on cellular proliferation induced by zearalenone and its derivatives in relation to the estrogenic parameters. Toxicol. Lett. 2005, 159, 272–283. [Google Scholar] [CrossRef]
- Leblanc, J.C.; Tard, A.; Volatier, J.L.; Verger, P. Estimated dietary exposure to principal food mycotoxins from the first French Total Diet Study. Food Addit. Contam. 2005, 22, 652–672. [Google Scholar] [CrossRef]
- FAO/WHO (Food and Agriculture Organization—World Health Organization). Zearalenone. Prepared by the Fifty-third meeting of the Joint FAO/WHO Expert Committee on Food Additives (JECFA). In Safety Evaluation of Certain Food Additives and Contaminants, WHO Food Additives Series 44; International Programme on Chemical Safety, World Health Organization: Geneva, Switzerland, 2000. [Google Scholar]
- EFSA—European Food Safety Authority. Panel on contaminants in the food chain. Scientific opinion on the risks for human and animal health related to the presence of modified forms of certain mycotoxins in food and feed. EFSA J. 2014, 12, 3916. [Google Scholar]
- Lemke, S.L.; Grant, P.G.; Phillips, T.D. Adsorption of zearalenone by organophilic montmorillonite clay. J. Agric. Food Chem. 1998, 46, 3789–3796. [Google Scholar] [CrossRef]
- Ryu, D.; Hanna, M.A.; Bullerman, L.B. Stability of zearalenone during extrusion of corn grits. J. Food Prot. 1999, 62, 1482–1484. [Google Scholar] [CrossRef]
- Carreau, S.; Bouraima-Lelong, H.; Delalande, C. Role of estrogens in spermatogenesis. Front. Biosci. 2012, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Carreau, S.; Bourguiba, S.; Lambard, S.; Galeraud-Denis, I.; Genissel, C.; Levallet, J. Reproductive system: Aromatase and estrogens. Mol. Cell. Endocrinol. 2002, 193, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, P.N. Aromatase inhibitors for male infertility. Fertil. Steril. 2012, 98, 1359–1362. [Google Scholar] [CrossRef] [PubMed]
- Gill-Sharma, M.K.; Dsouza, S.; Padwal, V.; Balasinor, N.; Aleem, M.; Parte, P.; Juneja, H.S. Antifertility effects of estradiol in adult male rats. J. Endocrinol. Investig. 2001, 24, 598–607. [Google Scholar] [CrossRef]
- MacCalman, C.D.; Getsios, S.; Farookhi, R.; Blaschuk, O.W. Estrogens potentiate the stimulatory effects of follicle-stimulating hormone on N-cadherin messenger ribonucleic acid levels in cultured mouse Sertoli cells. Endocrinology 1997, 138, 41–48. [Google Scholar] [CrossRef]
- MacCalman, C.D.; Farookhi, R.; Blaschuk, O.W. Estradiol regulates E-cadherin mRNA levels in the surface epithelium of the mouse ovary. Clin. Exp. Metastasis 1994, 12, 276–282. [Google Scholar] [CrossRef]
- Benzoni, E.; Minervini, F.; Giannoccaro, A.; Fornelli, F.; Vigo, D.; Visconti, A. Influence of in vitro exposure to mycotoxin zearalenone and its derivatives on swine sperm quality. Reprod. Toxicol. 2008, 25, 461–467. [Google Scholar] [CrossRef]
- Bielas, W.; Niżański, W.; Nicpoń, J.; Nicpoń, J.E.; Partyka, A.; Mordak, R.; Nowak, M.; Ciaputa, R. Effect of zearalenone on circulating testosterone concentration, testicular and epididymal morphology and epididymal sperm characteristics in wild boars. Theriogenology 2017, 102, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Chao, H.H.; Wang, L.; Ma, H.H.; Zhao, A.H.; Xiao, H.W.; Zhang, X.F. Identification of apoptotic pathways in zearalenone-treated mouse sertoli cells. J. Toxicol. Sci. 2022, 47, 257–268. [Google Scholar] [CrossRef]
- Chen, J.; Wang, M.; Wang, H.; Long, M. Zearalenone promotes apoptosis of mouse Leydig cells by targeting phosphatase and tensin homolog and thus inhibiting the PI3K/AKT signal pathway. Environ. Sci. Pollut. Res. Int. 2021, 28, 67779–67787. [Google Scholar] [CrossRef]
- Filannino, A.; Stout, T.A.; Gadella, B.M.; Sostaric, E.; Pizzi, F.; Colenbrander, B.; Dell’Aquila, M.E.; Minervini, F. Dose-response effects of estrogenic mycotoxins (zearalenone, alpha- and beta-zearalenol) on motility, hyperactivation and the acrosome reaction of stallion sperm. Reprod. Biol. Endocrinol. 2011, 9, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Zhao, Y.; Zhang, H.; Zhang, P.; Liu, J.; Feng, Y.; Men, Y.; Li, L.; Shen, W.; Sun, Z.; et al. Pubertal exposure to low doses of zearalenone disrupting spermatogenesis through ERα related genetic and epigenetic pathways. Toxicol. Lett. 2019, 315, 31–38. [Google Scholar] [CrossRef]
- Krejcárková, A.; Folková, P.; Šimoník, O.; Šašková, M.; Krejčířová, R.; Drábek, O.; Rajmon, R. Effects of zearalenone, α-zearalenol, and genistein on boar sperm motility in vitro. Czech J. Anim. Sci. 2017, 62, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Zhang, T.; Ren, X.; Li, B.; Wang, S. Male reproductive toxicity of zearalenone-meta-analysis with mechanism review. Ecotoxicol. Environ. Saf. 2021, 221, 112457. [Google Scholar] [CrossRef]
- Lin, P.; Chen, F.; Sun, J.; Zhou, J.; Wang, X.; Wang, N.; Li, X.; Zhang, Z.; Wang, A.; Jin, Y. Mycotoxin zearalenone induces apoptosis in mouse Leydig cells via an endoplasmic reticulum stress-dependent signalling pathway. Reprod. Toxicol. 2015, 52, 71–77. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, Y.; Gu, J.; Yuan, Y.; Liu, X.; Zheng, W.; Huang, Q.; Liu, Z.; Bian, J. Zearalenone inhibits testosterone biosynthesis in mouse Leydig cells via the crosstalk of estrogen receptor signaling and orphan nuclear receptor Nur77 expression. Toxicol. Vitr. 2014, 28, 647–656. [Google Scholar] [CrossRef]
- Long, M.; Yang, S.; Dong, S.; Chen, X.; Zhang, Y.; He, J. Characterization of semen quality, testicular marker enzyme activities and gene expression changes in the blood testis barrier of Kunming mice following acute exposure to zearalenone. Environ. Sci. Pollut. Res. Int. 2017, 24, 27235–27243. [Google Scholar] [CrossRef] [PubMed]
- Savard, C.; Gawhary, S.; Boyer, A.; Chorfi, Y. Assessment of Zearalenone-Induced Cell Survival and of Global Gene Regulation in Mouse TM4 Sertoli Cells. Toxins 2022, 14, 98. [Google Scholar] [CrossRef]
- She, J.; Feng, N.; Zheng, W.; Zheng, H.; Cai, P.; Zou, H.; Yuan, Y.; Gu, J.; Liu, Z.; Bian, J. Zearalenone Exposure Disrupts Blood-Testis Barrier Integrity through Excessive Ca2+-Mediated Autophagy. Toxins 2021, 13, 875. [Google Scholar] [CrossRef]
- Tassis, P.D.; Reisinger, N.; Nagl, V.; Tzika, E.; Schatzmayr, D.; Mittas, N.; Basioura, A.; Michos, I.; Tsakmakidis, I.A. Comparative Effects of Deoxynivalenol, Zearalenone and Its Modified Forms De-Epoxy-Deoxynivalenol and Hydrolyzed Zearalenone on Boar Semen In Vitro. Toxins 2022, 14, 497. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, W.; Bian, X.; Yuan, Y.; Gu, J.; Liu, X.; Liu, Z.; Bian, J. Zearalenone induces apoptosis and cytoprotective autophagy in primary Leydig cells. Toxicol. Lett. 2014, 226, 182–191. [Google Scholar] [CrossRef]
- Filipiak, E.; Walczak-Jedrzejowska, R.; Oszukowska, E.; Guminska, A.; Marchlewska, K.; Kula, K.; Slowikowska-Hilczer, J. Xenoestrogens diethylstilbestrol and zearalenone negatively influence pubertal rat’s testis. Folia Histochem. Cytobiol. 2009, 47, S113–S120. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Wang, Y.; Ma, L.; Chen, X.; Lü, Y.; Ge, F.; Chen, Y.; Chen, X.; Lian, Q.; Jin, X.D.; et al. Zearalenone Delays Rat Leydig Cell Regeneration. Toxicol. Sci. 2018, 164, 60–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dostalova, P.; Zatecka, E.; Dvorakova-Hortova, K. Of Oestrogens and Sperm: A Review of the Roles of Oestrogens and Oestrogen Receptors in Male Reproduction. Int. J. Mol. Sci. 2017, 18, 904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrell, D.T.; Hammoud, S.S. The human sperm epigenome and its potential role in embryonic development. Mol. Hum. Reprod. 2010, 16, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Biermann, K.; Steger, K. Epigenetics in male germ cells. J. Androl. 2007, 28, 466–480. [Google Scholar] [CrossRef]
- Faulk, C.; Dolinoy, D.C. Timing is everything: The when and how of environmentally induced changes in the epigenome of animals. Epigenetics 2011, 6, 791–797. [Google Scholar] [CrossRef] [Green Version]
- Baccarelli, A.; Bollati, V. Epigenetics and environmental chemicals. Curr. Opin. Pediatr. 2009, 21, 243–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, E. Chromatin modification and epigenetic reprogramming in mammalian development. Nat. Rev. Genet. 2002, 3, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Li, E. Structure and function of eukaryotic DNA methyltransferases. Curr. Top. Dev. Biol. 2004, 60, 55–89. [Google Scholar] [CrossRef] [PubMed]
- Goll, M.G.; Bestor, T.H. Eukaryotic cytosine methyltransferases. Annu. Rev. Biochem. 2005, 74, 481–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, H.; Wen, L.; Liao, S.; Lin, X.; Ma, T.; Liu, J.; Song, C.X.; Wang, M.; He, C.; Han, C.; et al. Dynamics of 5-hydroxymethylcytosine during mouse spermatogenesis. Nat. Commun. 2013, 4, 1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, K.R.; Veselovska, L.; Kelsey, G. Establishment and functions of DNA methylation in the germline. Epigenomics 2016, 10, 1399–1413. [Google Scholar] [CrossRef] [Green Version]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourc’his, D.; Bestor, T.H. Meiotic catastrophe and retrotransposon reactivation in male germ cells lacking Dnmt3L. Nature 2004, 431, 96–99. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology. 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Dumasia, K.; Kumar, A.; Deshpande, S.; Balasinor, N.H. Estrogen, through estrogen receptor 1, regulates histone modifications and chromatin remodeling during spermatogenesis in adult rats. Epigenetics 2017, 12, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Godmann, M.; Auger, V.; Ferraroni-Aguiar, V.; Sauro, A.; Sette, C.; Behr, R.; Kimmins, S. Dynamic regulation of histone H3 methylation at lysine 4 in mammalian spermatogenesis. Biol. Reprod. 2007, 77, 754–764. [Google Scholar] [CrossRef] [Green Version]
- Peters, A.H.; O’Carroll, D.; Scherthan, H.; Mechtler, K.; Sauer, S.; Schöfer, C.; Weipoltshammer, K.; Pagani, M.; Lachner, M.; Kohlmaier, A.; et al. Loss of the Suv39h histone methyltransferases impairs mammalian heterochromatin and genome stability. Cell 2001, 107, 323–337. [Google Scholar] [CrossRef]
- Mu, W.; Starmer, J.; Fedoriw, A.M.; Yee, D.; Magnuson, T. Repression of the soma-specific transcriptome by Polycomb-repressive complex 2 promotes male germ cell development. Genes Dev. 2014, 28, 2056–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielonka, Ł.; Gajęcka, M.; Zmudzki, J.; Gajęcki, M. The effect of selected environmental Fusarium mycotoxins on the ovaries of female wild boars (Sus scrofa). Pol. J. Vet. Sci. 2015, 18, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Cooke, P.S.; Nanjappa, M.K.; Ko, C.; Prins, G.S.; Hess, R.A. Estrogens in male physiology. Physiol. Rev. 2017, 97, 995–1043. [Google Scholar] [CrossRef]
- Swan, S.H.; Elkin, E.P.; Fenster, L. The question of declining sperm density revisited: An analysis of 101 studies published 1934-1996. Environ. Health Perspect. 2000, 108, 961–966. [Google Scholar] [CrossRef]
- Travison, T.G.; Araujo, A.B.; O’Donnell, A.B.; Kupelian, V.; McKinlay, J.B. A population-level decline in serum testosterone levels in American men. J. Clin. Endocrinol. Metab. 2007, 92, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Kavlock, R.J.; Daston, G.P.; DeRosa, C.; Fenner-Crisp, P.; Gray, L.E.; Kaattari, S.; Lucier, G.; Luster, M.; Mac, M.J.; Maczka, C.; et al. Research needs for the risk assessment of health and environmental effects of endocrine disruptors: A report of the U.S. EPA-sponsored workshop. Environ. Health Perspect. 1996, 104, 715–740. [Google Scholar] [CrossRef] [Green Version]
- Pettersson, K.; Gustafsson, J.A. Role of estrogen receptor β in estrogen action. Annu. Rev. Physiol. 2001, 63, 165–192. [Google Scholar] [CrossRef]
- Lee, K.H.; Hess, R.A.; Bahr, J.M.; Lubahn, D.B.; Taylor, J.; Bunick, D. Estrogen receptor α has a functional role in the mouse rete testis and efferent ductules. Biol. Reprod. 2000, 63, 1873–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummings, A.M. Methoxychlor as a model for environmental estrogens. Crit. Rev. Toxicol. 1997, 27, 367–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelce, W.R.; Monosson, E.; Gamcsik, M.P.; Laws, S.C.; Gray Jr, L.E. Environmental hormone disruptors: Evidence that vinclozolin developmental toxicity is mediated by antiandrogenic metabolites. Toxicol. Appl. Pharmacol. 1994, 126, 276–285. [Google Scholar] [CrossRef]
- Voccia, I.; Blakley, B.; Brousseau, P.; Fournier, M. Immunotoxicity of pesticides: A review. Toxicol. Ind. Health 1999, 15, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.L.; Goldman, J.M.; Stoker, T.E. Neuroendocrine and reproductive effects of contemporary-use pesticides. Toxicol. Ind. Health 1999, 15, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.S. Environmental anti-androgens and male reproductive health: Focus on phthalates and testicular dysgenesis syndrome. Reproduction 2004, 127, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Men, Y.; Zhao, Y.; Zhang, P.; Zhang, H.; Gao, Y.; Liu, J.; Feng, Y.; Li, L.; Shen, W.; Sun, Z.; et al. Gestational exposure to low-dose zearalenone disrupting offspring spermatogenesis might be through epigenetic modifications. Basic Clin. Pharmacol. Toxicol. 2019, 125, 382–393. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Li, Y.W.; Chen, Q.L.; Liu, Z.H. Long-term exposure of xenoestrogens with environmental relevant concentrations disrupted spermatogenesis of zebrafish through altering sex hormone balance, stimulating germ cell proliferation, meiosis and enhancing apoptosis. Environ. Pollut. 2018, 244, 486–494. [Google Scholar] [CrossRef]
- Olea, N.; Pazos, P.; Exposito, J. Inadvertent exposure to xenoestrogens. Eur. J. Cancer Prev. 1998, 7 (Suppl. S1), S17–S23. [Google Scholar] [CrossRef]
- Singleton, D.W.; Khan, S.A. Xenoestrogen exposure and mechanisms of endocrine disruption. Front. Biosci. 2003, 8, S110–S118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, S.O.; Korach, K.S. Mechanisms of Estrogen Receptor-Mediated Agonistic and Antagonistic Effects. In Endocrine Disruptors—Part I. The Handbook of Environmental Chemistry; Metzler, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2001; Volume 3L, pp. 1–25. [Google Scholar]
- Hall, J.M.; Couse, J.F.; Korach, K.S. The Multifaceted Mechanisms of Estradiol and Estrogen Receptor Signaling. J. Biol. Chem. 2001, 276, 36869–36872. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Li, S.S. Epigenetic effects of environmental chemicals bisphenol A and phthalates. Int. J. Mol. Sci. 2012, 13, 10143–10153. [Google Scholar] [CrossRef] [PubMed]
- Mirocha, C.J.; Pathre, S.V.; Christensen, C.M. Zearalenone. In Mycotoxins in Human and Animal Health; Rodricks, J.V., Hesseltine, C.W., Mehlman, M.A., Eds.; Pathotox Publisher Inc.: Park Forest South, IL, USA, 1977; pp. 346–364. [Google Scholar]
- Tsakmakidis, I.A.; Lymberopoulos, A.G.; Alexopoulos, C.; Boscos, C.M.; Kyriakis, S.C. In vitro effect of zearalenone and α-zearalenol on boar sperm characteristics and acrosome reaction. Reprod. Dom. Anim. 2006, 41, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Zhou, Q.; Sun, X.; Li, L.; Zhou, B.; Zeng, F.; Zhao, Y.; Shen, W.; Sun, Z. Effect of low-dose zearalenone exposure on reproductive capacity of male mice. Toxicol. Appl. Pharmacol. 2017, 333, 60–67. [Google Scholar] [CrossRef]
- Koraïchi, F.; Inoubli, L.; Lakhdari, N.; Meunier, L.; Vega, A.; Mauduit, C.; Benahmed, M.; Prouillac, C.; Lecoeur, S. Neonatal exposure to zearalenone induces long term modulation of ABC transporter expression in testis. Toxicology 2013, 310, 29–38. [Google Scholar] [CrossRef]
- Zheng, W.; Pan, S.; Wang, G.; Wang, Y.J.; Liu, Q.; Gu, J.; Yuan, Y.; Liu, X.Z.; Liu, Z.P.; Bian, J.C. Zearalenone impairs the male reproductive system functions via inducing structural and functional alterations of sertoli cells. Environ. Toxicol. Pharmacol. 2016, 42, 146–155. [Google Scholar] [CrossRef]
- Xu, M.L.; Hu, J.; Guo, B.P.; Niu, Y.R.; Xiao, C.; Xu, Y.X. Exploration of intrinsic and extrinsic apoptotic pathways in zearalenone-treated rat sertoli cells. Environ. Toxicol. 2016, 31, 1731–1739. [Google Scholar] [CrossRef]
- Li, D.K.; Zhou, Z.; Miao, M.; He, Y.; Wang, J.; Ferber, J.; Herrinton, L.J.; Gao, E.; Yuan, W. Urine bisphenol-A (BPA) level in relation to semen quality. Fertil. Steril. 2011, 95, 625–630.e4. [Google Scholar] [CrossRef]
- Chitra, K.C.; Latchoumycandane, C.; Mathur, P.P. Induction of oxidative stress by bisphenol A in the epididymal sperm of rats. Toxicology 2003, 185, 119–127. [Google Scholar] [CrossRef]
- Quignot, N.; Arnaud, M.; Robidel, F.; Lecomte, A.; Tournier, M.; Cren-Olive, C.; Barouki, R.; Lemazurier, E. Characterization of endocrine-disrupting chemicals based on hormonal balance disruption in male and female adult rats. Reprod. Toxicol. 2012, 33, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Herath, C.B.; Jin, W.; Watanabe, G.; Arai, K.; Suzuki, A.K.; Taya, K. Adverse effects of environmental toxicants, octylphenol and bisphenol A, on male reproductive functions in pubertal rats. Endocrine 2004, 25, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Duan, W.; Li, R.; Xu, S.; Zhang, L.; Chen, C.; He, M.; Lu, Y.; Wu, H.; Pi, H.; et al. Exposure to bisphenol A disrupts meiotic progression during spermatogenesis in adult rats through estrogen-like activity. Cell Death Dis. 2013, 4, e676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, M.; Bu, P.; Li, F.; Lan, S.; Wu, H.; Yuan, L.; Wang, Y. Neonatal bisphenol A exposure induces meiotic arrest and apoptosis of spermatogenic cells. Oncotarget 2016, 7, 10606–10615. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.-L.; Zhang, X.-F.; Feng, Y.-M.; Li, L.; Huynh, E.; Sun, X.-F.; Sun, Z.Y.; Shen, W. Exposure to bisphenol A results in a decline in mouse spermatogenesis. Reprod. Fertil. Dev. 2013, 25, 847–859. [Google Scholar] [CrossRef]
- Salian, S.; Doshi, T.; Vanage, G. Neonatal exposure of male rats to Bisphenol A impairs fertility and expression of sertoli cell junctional proteins in the testis. Toxicology 2009, 12, 56–67. [Google Scholar] [CrossRef]
- Salian, S.; Doshi, T.; Vanage, G. Perinatal exposure of rats to bisphenol A affects the fertility of male offspring. Life Sci. 2009, 85, 742–752. [Google Scholar] [CrossRef]
- Ullah, A.; Pirzada, M.; Jahan, S.; Ullah, H.; Turi, N.; Ullah, W.; Siddiqui, M.F.; Zakria, M.; Lodhi, K.Z.; Khan, M.M. Impact of low-dose chronic exposure to bisphenol A and its analogue bisphenol B, bisphenol F and bisphenol S on hypothalamo-pituitary- testicular activities in adult rats: A focus on the possible hormonal mode of action. Food Chem. Toxicol. 2018, 121, 24–36. [Google Scholar] [CrossRef]
- Doshi, T.; Mehta, S.S.; Dighe, V.; Balasinor, N.; Vanage, G. Hypermethylation of estrogen receptor promoter region in adult testis of rats exposed neonatally to bisphenol A. Toxicology 2011, 289, 74–82. [Google Scholar] [CrossRef]
- Frizzell, C.; Ndossi, D.; Verhaegen, S.; Dahl, E.; Eriksen, G.; Sørlie, M.; Ropstad, E.; Muller, M.; Elliott, C.T.; Connolly, L. Endocrine disrupting effects of zearalenone, alpha-and beta-zearalenol at the level of nuclear receptor binding and steroidogenesis. Toxicol. Lett. 2011, 206, 210–217. [Google Scholar] [CrossRef]
- Bentov, Y.; Jurisicova, A.; Kenigsberg, S.; Casper, R.F. What maintains the high intra-follicular estradiol concentration in pre-ovulatory follicles? J. Assist. Reprod. Genet. 2016, 33, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poór, M.; Kunsági-Máté, S.; Bálint, M.; Hetényi, C.; Gerner, Z.; Lemli, B. Interaction of mycotoxin zearalenone with human serum albumin. J. Photochem. Photobiol. B 2017, 170, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Mauro, T.; Hao, L.; Pop, L.C.; Buckley, B.; Schneider, S.H.; Bandera, E.V.; Shapses, S.A. Circulating zearalenone and its metabolites differ in women due to body mass index and food intake. Food Chem. Toxicol. 2018, 116, 227–232. [Google Scholar] [CrossRef]
- Abid-Essefi, S.; Ouanes, Z.; Hassen, W.; Baudrimont, I.; Creppy, E.; Bacha, H. Cytotoxicity, inhibition of DNA and protein syntheses and oxidative damage in cultured cells exposed to zearalenone. Toxicol. Vitr. 2004, 18, 467–474. [Google Scholar] [CrossRef]
- Gazzah, A.C.; Golli Bennour, E.; Bouaziz, C.; Abid, S.; Ladjimi, M.; Bacha, H. Sequential events of apoptosis induced by zearalenone in cultured hepatocarcinoma cells. Mycotox. Res. 2010, 26, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Arai, N.; Ström, A.; Rafter, J.J.; Gustaffson, J.-Å. Estrogen Receptor β mRNA in Colon Cancer Cells: Growth Effects of Estrogen and Genistein. Biochem. Biophys. Res. Commun. 2000, 270, 425–431. [Google Scholar] [CrossRef]
- Kouadio, J.H.; Mobio, T.A.; Baudrimont, I.; Moukha, S.; Dano, S.D.; Creppy, E.E. Comparative study of cytotoxicity and oxidative stress induced by deoxynivalenol, zearalenone or fumonisin B1 in human intestinal cell line Caco-2. Toxicology 2005, 213, 56–65. [Google Scholar] [CrossRef]
- Yiannokouris, A.; Francois, J.; Poughon, L.; Dussap, C.-G.; Bertin, G.; Jeminet, G.; Jouany, J.-P. Alkali extraction of β-D-glucans from Saccharomyces cerevisiae cell wall and study of their adsorptive properties toward zearalenone. J. Agric. Food Chem. 2004, 52, 3666–3673. [Google Scholar] [CrossRef]
- Ouanes, Z.; Abid, S.; Ayed, I.; Anane, R.; Mobio, T.; Creppy, E.E.; Bacha, H. Induction of micronuclei by zearalenone in Vero monkey kidney cells and in bone marrow cells of mice: Protective effect of vitamin E. Mutat. Res. 2003, 538, 63–70. [Google Scholar] [CrossRef]
- Wu, T.S.; Liao, Y.C.; Yu, F.Y.; Chang, C.H.; Liu, B.H. Mechanism of patulin-induced apoptosis in human leukemia cells (HL-60). Toxicol. Lett. 2008, 183, 105–111. [Google Scholar] [CrossRef]
- Bouaziz, C.; Sharif el dein, O.; El Golli, E.; Abid-Essefi, S.; Brenner, C.; Lemaire, C.; Bacha, H. Different apoptotic pathways induced by zearalenone, T-2 toxin and ochratoxin A in human hepatoma cells. Toxicology 2008, 254, 19–28. [Google Scholar] [CrossRef]
- Ayed-Boussema, I.; Bouaziz, C.; Rjiba, K.; Valenti, K.; Laporte, F.; Bacha, H.; Hassen, W. The mycotoxin zearalenone induces apoptosis in human hepatocytes (HepG2) via p53-dependent mitochondrial signaling pathway. Toxicol. Vitr. 2008, 22, 1671–1780. [Google Scholar] [CrossRef]
- Lee, R.; Kim, D.W.; Lee, W.Y.; Park, H.J. Zearalenone Induces Apoptosis and Autophagy in a Spermatogonia Cell Line. Toxins 2022, 14, 148. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 4th ed.; Oxford University Press: New York, NY, USA, 2007. [Google Scholar]
- Hassen, W.; Ayed-Boussema, I.; Oscoz, A.A.; Lopez, A.D.C.; Bacha, H. The role of oxidative stress in zearalenone-mediated toxicity in HepG2 cells: Oxidative DNA damage, gluthatione depletion and stress proteins induction. Toxicology 2007, 232, 294–302. [Google Scholar] [CrossRef]
- Abid-Essefi, S.; Bouaziz, C.; El Golli-Bennour, E.; Ouanes, Z.; Bacha, H. Comparative study of toxic effects of zearalenone and its two major metabolites α-zearalenol and β-zearalenol on cultured human Caco-2 cells. J. Biochem. Mol. Toxic. 2009, 23, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Abid-Essefi, S.; Zaied, C.; Bouaziz, C.; Salem, I.B.; Kaderi, R.; Bacha, H. Protective effect of aquosus extract of Allium sativum against zearalenone toxicity mediated by oxidative stress. Exp. Toxicol. Pathol. 2012, 64, 689–695. [Google Scholar] [CrossRef]
- Bennour, E.G.; Bouaziz, C.; Ladjimi, M.; Renaud, F.; Bacha, H. Comparative mechanisms of zearalenone and ochratoxin A toxicities on cultured HepG2 cells: Oxidative stress a common process? Environ. Toxicol. 2008, 24, 538–548. [Google Scholar] [CrossRef]
- Stadnik, A.; Wójtowitz-Chomicz, K.; Borzecki, A. Influence of zearalenone on free radical reactions in rat liver cells. Bull. Vet. Inst. Pulawy 2010, 54, 611–615. [Google Scholar]
- Othmen, Z.O.; Golli, E.E.; Abid-Essefi, S.; Bacha, H. Cytotoxicity effects induced by Zearalenone metabolites, alpha Zearalenol and beta Zearalenol, on cultured Vero cells. Toxicology 2008, 252, 72–77. [Google Scholar] [CrossRef]
- Hartl, F.U. Molecular chaperones in cellular protein folding. Nature 1996, 381, 571–579. [Google Scholar] [CrossRef]
- Mike, N.; Papp, G.; Certik, M.; Czibulya, Z.; Kunsági-Máté, S.; Ember, I.; Vágvölgyi, C.; Pesti, M.; Gazdag, Z. Regulation of cytotoxic, non-estrogenic, oxidative stress-induced processes of zearalenone in the fission yeast Schizosaccharomyces pombe. Toxicon 2013, 73, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Shen, T.; Ding, Q.; Lv, Y.; Li, L.; Huang, K.; Yan, L.; Song, S. Zearalenone induces ROS-mediated mitochondrial damage in porcine IPEC-J2 cells. J. Biochem. Mol. Toxicol. 2017, 31, e21944. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Gao, R.; Meng, Q.; Zhang, Y.; Bi, C.; Shan, A. Toxic effects of maternal zearalenone exposure on intestinal oxidative stress, barrier function, immunological and morphological changes in rats. PLoS ONE 2014, 9, e106412. [Google Scholar] [CrossRef] [Green Version]
- Salah-Abbès, J.B.; Abbès, S.; Abdel-Wahhab, M.A.; Oueslati, R. Raphanus sativus extract protects against Zearalenone induced reproductive toxicity, oxidative stress and mutagenic alterations in male Balb/c mice. Toxicon 2009, 53, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Boeira, S.P.; Filho, C.B.; Del’Fabbro, L.; Royes, L.F.F.; Jessé, C.R.; Oliveira, M.S.; Furian, A.F. Possible role for glutathione S-transferase in the oligozoospermia elicited by acute zearalenone administration in Swiss albino mice. Toxicon 2012, 60, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Jin, Y.; Zhao, Y.; Shan, A.; Fang, H.; Shen, J.; Zhou, C.; Yu, H.; Zhou, Y.F.; Wang, X.; et al. Zearalenone induces apoptosis in bovine mammary epithelial cells by activating endoplasmic reticulum stress. J. Dairy Sci. 2019, 102, 10543–10553. [Google Scholar] [CrossRef]
- Kim, I.H.; Son, H.Y.; Cho, S.W.; Ha, C.S.; Kang, B.H. Zearalenone induces male germ cell apoptosis in rats. Toxicol. Lett. 2003, 138, 185–192. [Google Scholar] [CrossRef]
- Elweza, A.E.; Marey, M.A.; Elesh, I.F.; Zinnah, M.A.; Akthar, I.; Kanno, C.; Takagi, M.; Miyamoto, A. Zearalenone interferes with the sperm-triggered inflammation in the bovine uterus in vitro: Negative impact on sperm motility and survival. Reprod. Toxicol. 2022, 107, 81–89. [Google Scholar] [CrossRef]
- Nieschlag, E.; Behre, H.M.; Nieschlag, S. Andrology: Male Reproductive Health and Dysfunction, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 12–16. [Google Scholar]
- Cai, G.; Si, M.; Li, X.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Liu, Z.; Bian, J. Zearalenone induces apoptosis of rat Sertoli cells through Fas-Fas ligand and mitochondrial pathway. Environ. Toxicol. 2019, 34, 424–433. [Google Scholar] [CrossRef]
- Alahmar, A.T. Role of Oxidative Stress in Male Infertility: An Updated Review. J. Hum. Reprod. Sci. 2019, 12, 4–18. [Google Scholar] [CrossRef]
- Agarwal, A.; Parekh, N.; Panner Selvam, M.K.; Henkel, R.; Shah, R.; Homa, S.T.; Ramasamy, R.; Ko, E.; Tremellen, K.; Esteves, S.; et al. Male Oxidative Stress Infertility (MOSI): Proposed Terminology and Clinical Practice Guidelines for Management of Idiopathic Male Infertility. World J. Men’s Health 2019, 37, 296–312. [Google Scholar] [CrossRef]
- Bisht, S.; Faiq, M.; Tolahunase, M.; Dada, R. Oxidative stress and male infertility. Nat. Rev. Urol. 2017, 14, 470–485. [Google Scholar] [CrossRef]
- Takeshima, T.; Usui, K.; Mori, K.; Asai, T.; Yasuda, K.; Kuroda, S.; Yumura, J. Oxidative stress and male infertility. Reprod. Med. Biol. 2021, 20, 41–52. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M. Oxidative stress, DNA damage and apoptosis in male infertility: A clinical approach. BJU Int. 2005, 95, 503–507. [Google Scholar] [CrossRef]
- Barbato, V.; Talevi, R.; Braun, S.; Merolla, A.; Sudhakaran, S.; Longobardi, S.; Gualtieri, R. Supplementation of sperm media with zinc, D-aspartate and co-enzyme Q10 protects bull sperm against exogenous oxidative stress and improves their ability to support embryo development. Zygote 2017, 25, 168–175. [Google Scholar] [CrossRef]
- Lopes, S.; Jurisicova, A.; Sun, J.G.; Casper, R.F. Reactive oxygen species: Potential cause for DNA fragmentation in human spermatozoa. Hum. Reprod. 1998, 13, 896–900. [Google Scholar] [CrossRef]



| First Author | Reference Number | Species | In Vitro/In Vivo | Method (Oral, ip, etc) | Dose | Exposure Period | Results |
|---|---|---|---|---|---|---|---|
| Gao | [39] | CD-1 male mice | In vivo | Orally once a day | 20 μg/kg and 40 μg/kg bw ZEA | 5 w | Disrupted process of meiosis, inhibited spermatogenesis and diminished semen quality with the decrease in spermatozoa motility and concentration. The DNA methylation markers 5mC and 5hmC were decreased, the histone methylation marker H3K27 was increased, at the same time estrogen receptor alpha was diminished. |
| Lin | [42] | MLTC-1 cells | In vitro | Single exposure | 0–200 μg/mL ZEA | 24 h | Reduced cell viability, increased rate of apoptosis, increased caspase-3 activity, reduced testosterone levels. |
| Liu | [43] | Mouse Leydig cells | In vitro | Single exposure | 0–20 μg/mL | 24 h | Reduced production of testosterone via crosstalk of estrogen receptor signaling, elevated cellular cAMP levels, reduced mitochondrial membrane potential, decreased expression of P450scc, 17b-HSD, and P450c17, increased expression of StAR and 3b- HSD, Nur77 expression was significantly inhibited. |
| Long | [44] | Kunming male mice | In vivo | Once a day, i.p. | 0- 75 mg/kg bw ZEA | 5 d | Reduced weights of testis and epididymis, sperm tail defects and head abnormalities, reduced concentration of testosterone, increased enzyme activities of LDH, AKP and ACP, increased mRNA expressions of Vim and Cldn11. |
| She | [46] | Male Balb/c mice and TM4 mouse Sertoli cells line | In vivo and in vitro | Orally once a day and single exposure | 40 mg/kg bw and 0–20 μM/L ZEA | 5–7 d and 24 h | Ultrastructural changes in testis, reduced sperm motility, altered expression of blood-testis-barrier proteins and autophagy-related proteins, increased level of cytoplasmic Ca2+. |
| Filipiak | [49] | Wistar rats | In vivo | Once a day, s.c. | 4 or 40 μg ZEA | 10 d | Reduced testes weight, seminiferous tubule diameter and length, reduced number of Sertoli cells. |
| Zhou | [50] | Sprague Dawley rats and immature Leydig cells | In vivo and in vitro | Once a day, intratesticularly and single exposure | 0–300 ng/testis and 0–50 μM ZEA | 21 d and 24 h | Reduced serum testosterone levels, reduced Leydig cell number and cell-specific gene/protein expression, reduced expression of steroidogenic factor 1 (Nr5a1), inhibited androgen production and steroidogenic enzyme activities in immature Leydig cells by downregulating expression levels of cholesterol side cleavage enzyme (Cyp11a1), 3β-hydroxysteroid dehydrogenase 1 (Hsd3b1), and steroid 5α-reductase 1 (Srd5a1). |
| Men | [81] | ICR pregnant mice and its male offspring | In vivo | Orally once a day | 0–40 µg/kg bw ZEA | 7 d | Disrupted meiosis, alteration in DNA/histone methylation (reduced 5hmC, increased H3K27me3, H3K9me2, G9a), higher ratio of ERα-positive Leydig cells, decreased sperm motility and concentration, disrupted sperm acrosome integrity, decreased body-weight, reduced testis weight, liver damage - increased AST. |
| Pang | [90] | CD-1 male mice | In vivo | Orally once a day | 20 or 40 μg/kg ZEA | 42 d | The spermatogenic cells were declined, increased rate of DNA breaks. |
| Koraïchi | [91] | Sprague Dawley rats and SerW3 Sertoli cell line | In vivo and in vitro | Once a day s.c. and single exposure | 0–100 μg /day ZEA | 5 d post-natal | Modulations of mRNA and protein levels of Abcb1, Abcc1, Abcg2, Abcc4 and Abcc5 were observed, along with Abcc4 protein cellular delocalization. |
| Frizell | [105] | RGA cell lines were generated from human mammary gland cell and H295R human adrenocortical carcinoma cells | In vitro | Single exposure | 0–100 μM ZEA, α-zearalenol and β-zearalenol | 48 h | α -zearalenol exhibited the strongest estrogenic potency, slightly less potent than 17 β - estradiol. ZEA was ∼70 times less potent than α -zearalenol and twice as potent as β -zearalenol. Binding of progesterone to the progestagen receptor was shown to be synergistically increased in the presence of ZEA, α -zearalenol or β -zearalenol. ZEA, α -zearalenol or β -zearalenol increased production of progesterone, estradiol, testosterone and cortisol hormones in the H295R steroidogenesis assay. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balló, A.; Busznyákné Székvári, K.; Czétány, P.; Márk, L.; Török, A.; Szántó, Á.; Máté, G. Estrogenic and Non-Estrogenic Disruptor Effect of Zearalenone on Male Reproduction: A Review. Int. J. Mol. Sci. 2023, 24, 1578. https://doi.org/10.3390/ijms24021578
Balló A, Busznyákné Székvári K, Czétány P, Márk L, Török A, Szántó Á, Máté G. Estrogenic and Non-Estrogenic Disruptor Effect of Zearalenone on Male Reproduction: A Review. International Journal of Molecular Sciences. 2023; 24(2):1578. https://doi.org/10.3390/ijms24021578
Chicago/Turabian StyleBalló, András, Kinga Busznyákné Székvári, Péter Czétány, László Márk, Attila Török, Árpád Szántó, and Gábor Máté. 2023. "Estrogenic and Non-Estrogenic Disruptor Effect of Zearalenone on Male Reproduction: A Review" International Journal of Molecular Sciences 24, no. 2: 1578. https://doi.org/10.3390/ijms24021578