
Metabolite Profiling of Conifer Needles: Tracing Pollution and Climate Effects

 , , , and
, , , and
Abstract
:1. Introduction
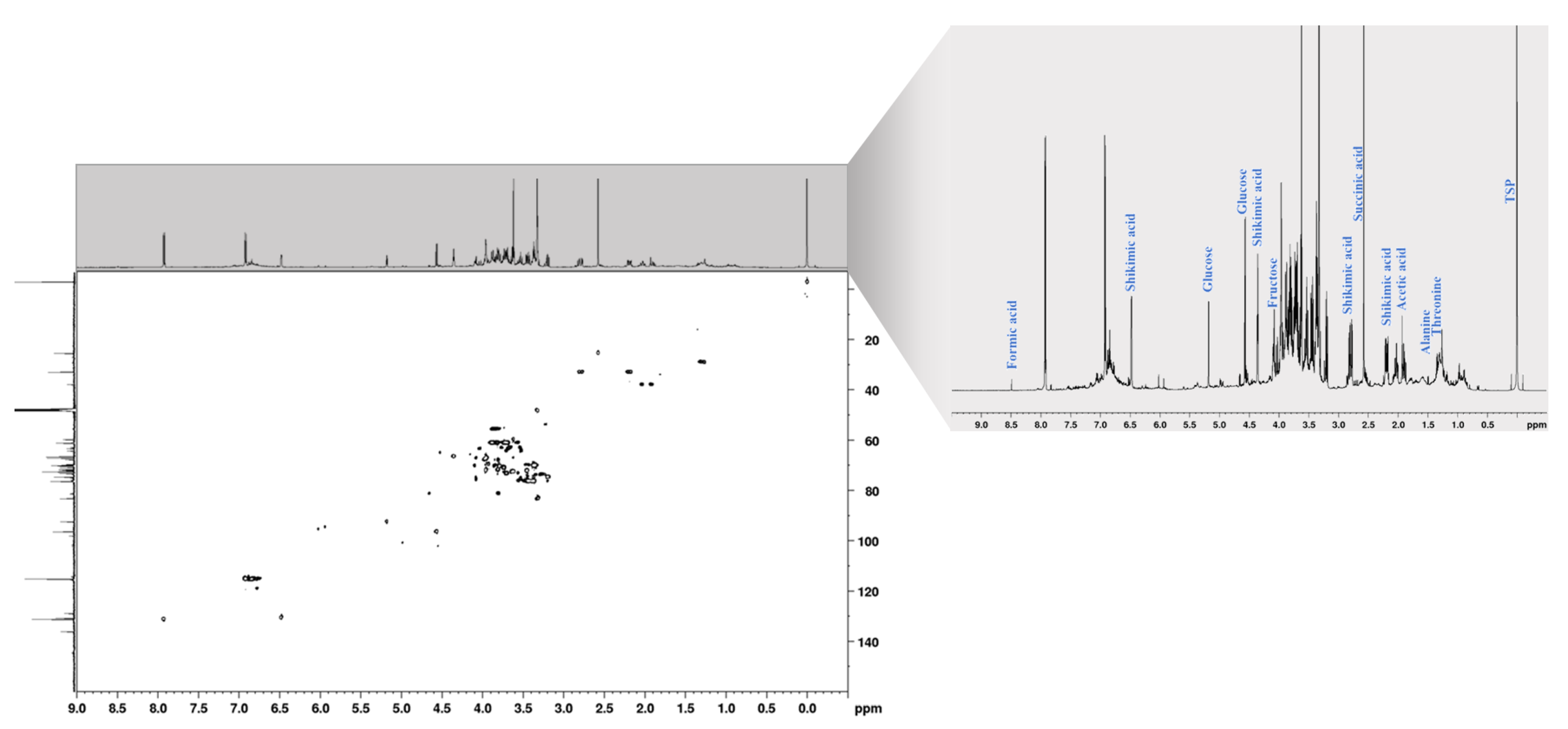
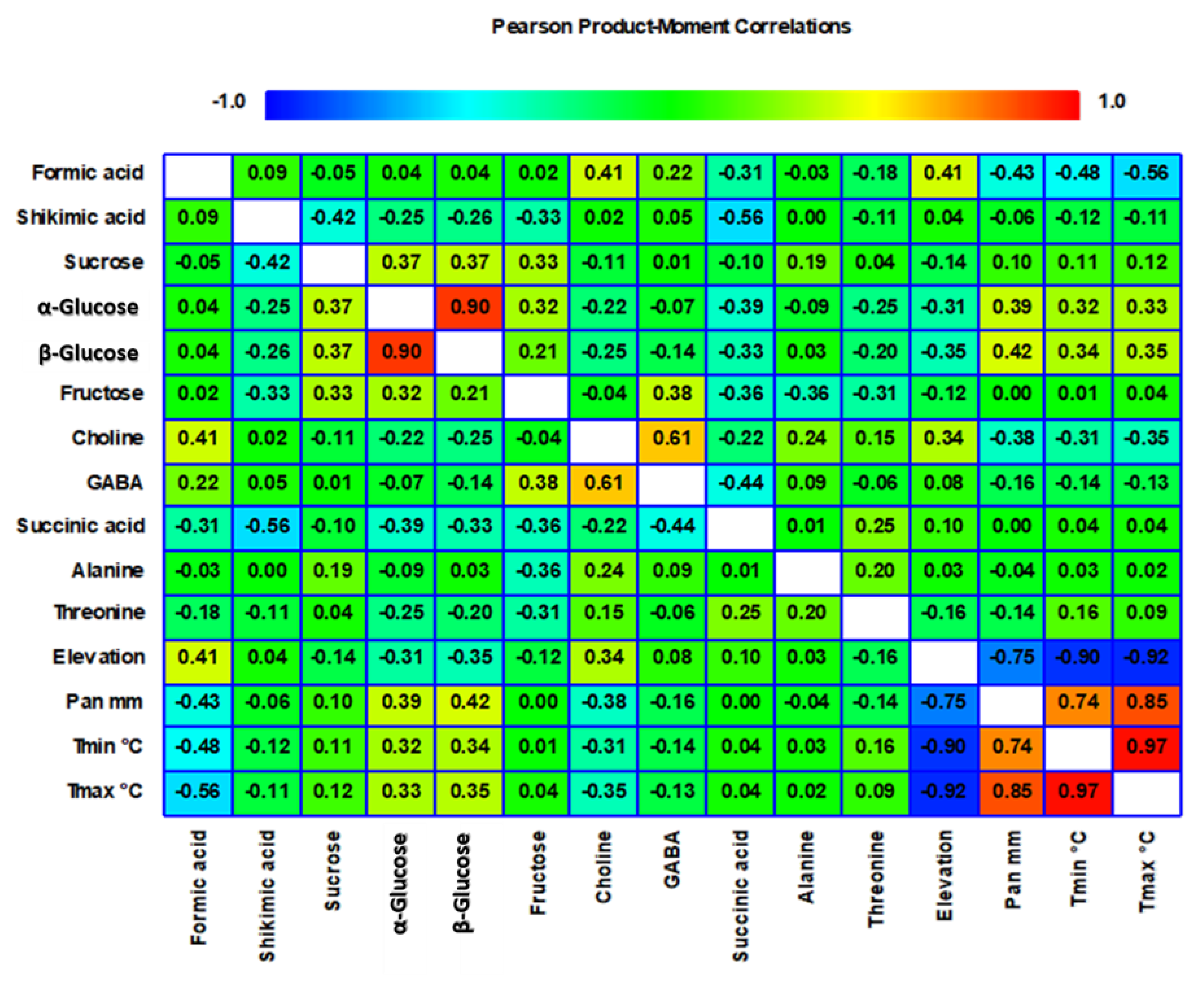
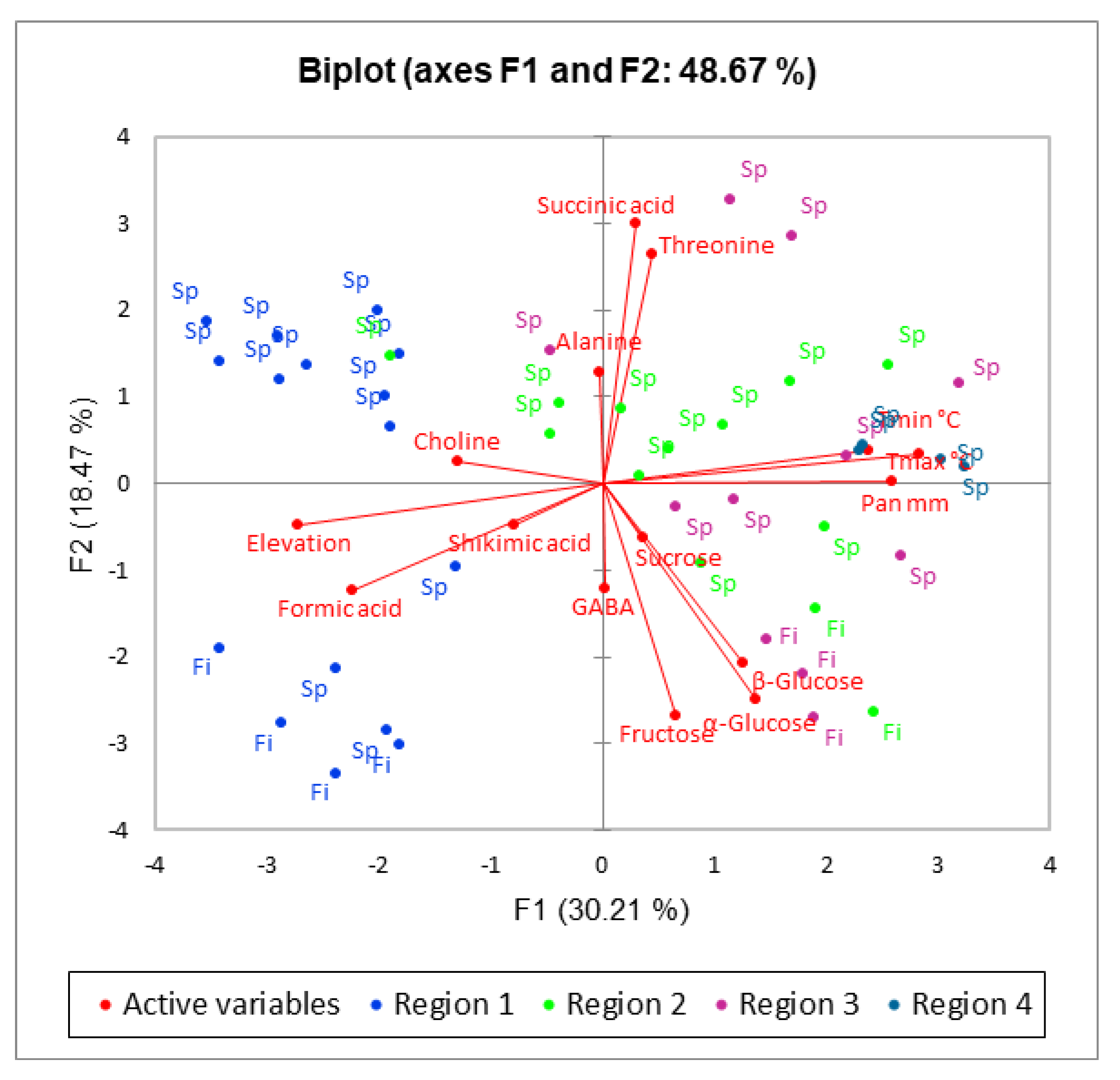
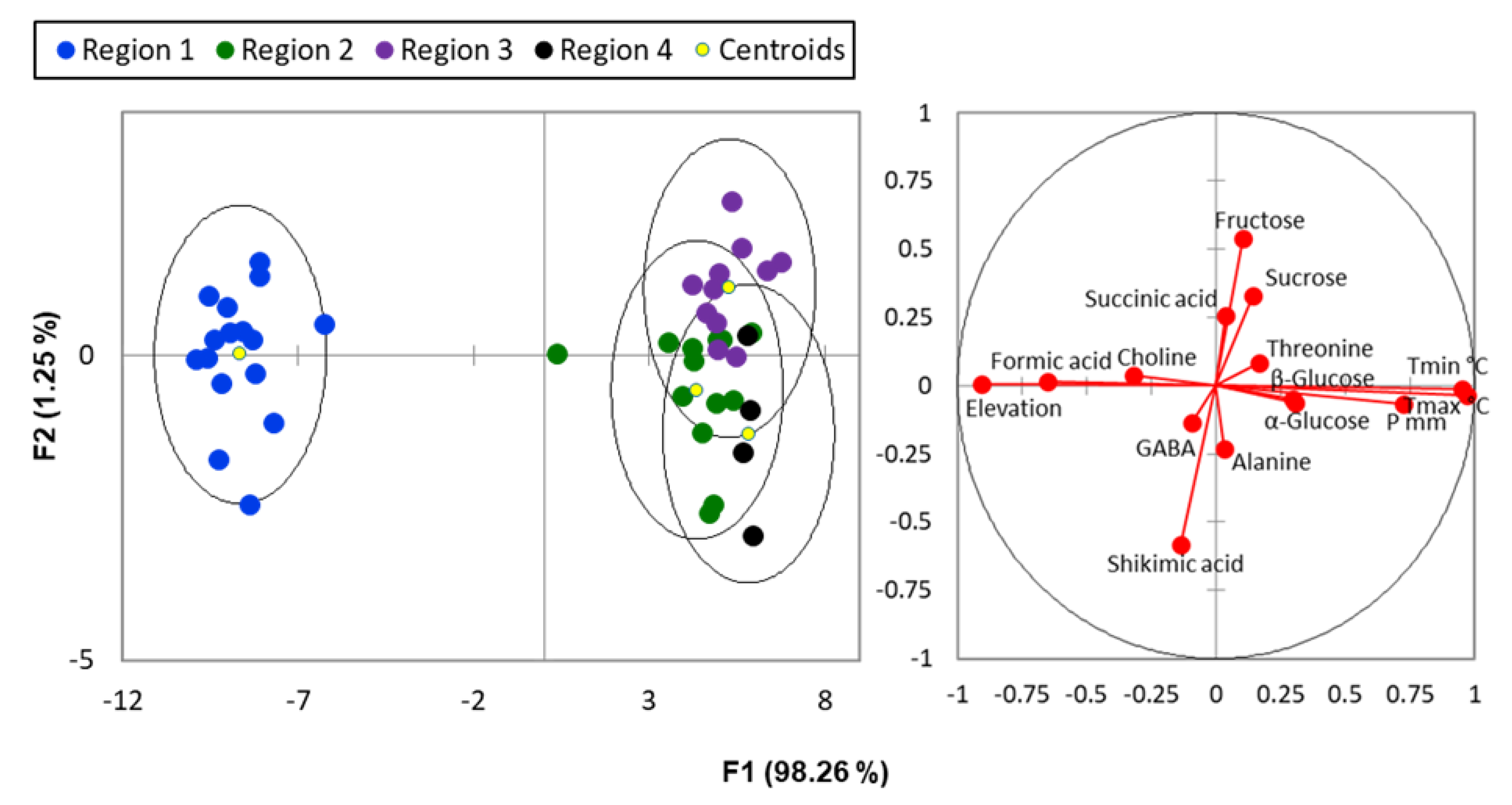
2. Results and Discussion
3. Materials and Methods
3.1. Sample Identification, Collection and Preparation
3.2. 1D and 2D NMR Spectroscopy
3.3. Metabolite Identification and Quantification
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kozlowski, T.T.; Pallardy, S.G. Acclimation and Adaptive Responses of Woody Plants to Environmental Stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Kramer, P.J.; Pallardy, S.G. Air Pollution. In The Physiological Ecology of Woody Plants; Elsevier: Amsterdam, The Netherlands, 1991; pp. 338–375. ISBN 978-0-12-424160-2. [Google Scholar]
- Rai, P.K. Impacts of Particulate Matter Pollution on Plants: Implications for Environmental Biomonitoring. Ecotoxicol. Environ. Saf. 2016, 129, 120–136. [Google Scholar] [CrossRef] [PubMed]
- Forbes, P.B.C.; van der Wat, L.; Kroukamp, E.M. Biomonitors. In Comprehensive Analytical Chemistry; Elsevier: Amsterdam, The Netherlands, 2015; Volume 70, pp. 53–108. ISBN 978-0-444-63553-2. [Google Scholar]
- De Nicola, F.; Spagnuolo, V.; Baldantoni, D.; Sessa, L.; Alfani, A.; Bargagli, R.; Monaci, F.; Terracciano, S.; Giordano, S. Improved Biomonitoring of Airborne Contaminants by Combined Use of Holm oak Leaves and Epiphytic Moss. Chemosphere 2013, 92, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Boltersdorf, S.H.; Pesch, R.; Werner, W. Comparative Use of Lichens, Mosses and Tree Bark to Evaluate Nitrogen Deposition in Germany. Environ. Pollut. 2014, 189, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Klánová, J.; Čupr, P.; Baráková, D.; Šeda, Z.; Anděl, P.; Holoubek, I. Can Pine Needles Indicate Trends in the Air Pollution Levels at Remote Sites? Environ. Pollut. 2009, 157, 3248–3254. [Google Scholar] [CrossRef]
- Rai, P.K. Biodiversity of Roadside Plants and Their Response to Air Pollution in an Indo-Burma Hotspot Region: Implications for Urban Ecosystem Restoration. J. Asia-Pac. Biodivers. 2016, 9, 47–55. [Google Scholar] [CrossRef]
- Shahid, M.; Dumat, C.; Khalid, S.; Schreck, E.; Xiong, T.; Niazi, N.K. Foliar Heavy Metal Uptake, Toxicity and Detoxification in Plants: A Comparison of Foliar and Root Metal Uptake. J. Hazard. Mater. 2017, 325, 36–58. [Google Scholar] [CrossRef]
- Anderson, K.A.; Smith, B.W. Effect of Season and Variety on the Differentiation of Geographic Growing Origin of Pistachios by Stable Isotope Profiling. J. Agric. Food Chem. 2006, 54, 1747–1752. [Google Scholar] [CrossRef]
- Armenise, L.; Simeone, M.C.; Piredda, R.; Schirone, B. Validation of DNA Barcoding as an Efficient Tool for Taxon Identification and Detection of Species Diversity in Italian Conifers. Eur. J. For. Res. 2012, 131, 1337–1353. [Google Scholar] [CrossRef]
- Stavrakou, T.; Müller, J.-F.; Peeters, J.; Razavi, A.; Clarisse, L.; Clerbaux, C.; Coheur, P.-F.; Hurtmans, D.; De Mazière, M.; Vigouroux, C.; et al. Satellite Evidence for a Large Source of Formic Acid from Boreal and Tropical Forests. Nat. Geosci. 2012, 5, 26–30. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Dutta, R.; Das, P. A Critical Review on Plant Biomonitors for Determination of Polycyclic Aromatic Hydrocarbons (PAHs) in Air through Solvent Extraction Techniques. Chemosphere 2020, 251, 126441. [Google Scholar] [CrossRef] [PubMed]
- Geana, E.I.; Ciucure, C.T.; Sandru, C.; Botoran, O.R.; Ionete, R.E. Coniferous Bark, Needles and Cones as Bioindicators of Air Pollution in Urban Areas with Intense Road Traffic. In Proceedings of the Book of Abstracts of the 21st International Conference “Life Sciences for Sustainable Development”; University og Agricultural Sciences and Veterinary Medicine Cluj-Napoca, Cluj Napoca, Romania, 15 September 2022. [Google Scholar]
- Winning, H.; Larsen, F.H.; Bro, R.; Engelsen, S.B. Quantitative Analysis of NMR Spectra with Chemometrics. J. Magn. Reson. 2008, 190, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Olate, V.; Soto, A.; Schmeda-Hirschmann, G. Seasonal Variation and Resin Composition in the Andean Tree Austrocedrus Chilensis. Molecules 2014, 19, 6489–6503. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.B.; Kozminski, M.A.; Santiago-Blay, J.A. Distinctions among Conifer Exudates by Proton Magnetic Resonance Spectroscopy. J. Nat. Prod. 2007, 70, 1283–1294. [Google Scholar] [CrossRef]
- Longobardi, F.; Ventrella, A.; Napoli, C.; Humpfer, E.; Schütz, B.; Schäfer, H.; Kontominas, M.G.; Sacco, A. Classification of Olive Oils According to Geographical Origin by Using 1H NMR Fingerprinting Combined with Multivariate Analysis. Food Chem. 2012, 130, 177–183. [Google Scholar] [CrossRef]
- Farag, M.A.; Porzel, A.; Wessjohann, L.A. Comparative Metabolite Profiling and Fingerprinting of Medicinal Licorice Roots Using a Multiplex Approach of GC–MS, LC–MS and 1D NMR Techniques. Phytochem. 2012, 76, 60–72. [Google Scholar] [CrossRef]
- Lassalle, G.; Fabre, S.; Credoz, A.; Hédacq, R.; Dubucq, D.; Elger, A. Mapping Leaf Metal Content over Industrial Brownfields Using Airborne Hyperspectral Imaging and Optimized Vegetation Indices. Sci. Rep. 2021, 11, 2. [Google Scholar] [CrossRef]
- Caggiano, R.; Speranza, A.; Imbrenda, V.; Afflitto, N.; Sabia, S. A Holistic Approach Based on Biomonitoring Techniques and Satellite Observations for Air Pollution Assessment and Health Risk Impact of Atmospheric Trace Elements in a Semi-Rural Area of Southern Italy (High Sauro Valley). Atmosphere 2022, 13, 1501. [Google Scholar] [CrossRef]
- Wägele, J.W.; Bodesheim, P.; Bourlat, S.J.; Denzler, J.; Diepenbroek, M.; Fonseca, V.; Frommolt, K.-H.; Geiger, M.F.; Gemeinholzer, B.; Glöckner, F.O.; et al. Towards a Multisensor Station for Automated Biodiversity Monitoring. Basic Appl. Ecol. 2022, 59, 105–138. [Google Scholar] [CrossRef]
- Melgarejo, P.; Núñez-Gómez, D.; Martínez-Nicolás, J.J.; Hernández, F.; Martínez-Font, R.; Lidón, V.; García-Sánchez, F.; Legua, P. Metabolomic Profile of Citrus limon Leaves (‘Verna’ Variety) by 1H-NMR and Multivariate Analysis Technique. Agronomy 2022, 12, 1060. [Google Scholar] [CrossRef]
- Hildebrandt, T.M. Synthesis versus Degradation: Directions of Amino Acid Metabolism during Arabidopsis Abiotic Stress Response. Plant. Mol. Biol. 2018, 98, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Amir, R.; Fernie, A.R. The Regulation of Essential Amino Acid Synthesis and Accumulation in Plants. Annu. Rev. Plant Biol. 2016, 67, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, T.; Tani, A. Emissions of Gaseous Formic and Acetic Acids from Major Tree Species in Japan. Atmos. Environ. 2021, 247, 118149. [Google Scholar] [CrossRef]
- Grodzinski, B. A Study of Formate Production and Oxidation in Leaf Peroxisomes during Photorespiration. Plant Physiol. 1979, 63, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Jardine, K.; Fernandes de Souza, V.; Oikawa, P.; Higuchi, N.; Bill, M.; Porras, R.; Niinemets, Ü.; Chambers, J. Integration of C1 and C2 Metabolism in Trees. Int. J. Mol. Sci. 2017, 18, 2045. [Google Scholar] [CrossRef]
- Fulgham, S.R.; Brophy, P.; Link, M.; Ortega, J.; Pollack, I.; Farmer, D.K. Seasonal Flux Measurements over a Colorado Pine Forest Demonstrate a Persistent Source of Organic Acids. ACS Earth Space Chem. 2019, 3, 2017–2032. [Google Scholar] [CrossRef]
- Khare, P.; Kumar, N.; Kumari, K.M.; Srivastava, S.S. Atmospheric Formic and Acetic Acids: An Overview. Rev. Geophys. 1999, 37, 227–248. [Google Scholar] [CrossRef]
- Liedvogel, B.; Stumpf, P.K. Origin of Acetate in Spinach Leaf Cell. Plant Physiol. 1982, 69, 897–903. [Google Scholar] [CrossRef]
- Jardine, K.J.; Sommer, E.D.; Saleska, S.R.; Huxman, T.E.; Harley, P.C.; Abrell, L. Gas Phase Measurements of Pyruvic Acid and Its Volatile Metabolites. Environ. Sci. Technol. 2010, 44, 2454–2460. [Google Scholar] [CrossRef]
- Li, C.; Liu, Y.; Tian, J.; Zhu, Y.; Fan, J. Changes in Sucrose Metabolism in Maize Varieties with Different Cadmium Sensitivities under Cadmium Stress. PLoS ONE 2020, 15, e0243835. [Google Scholar] [CrossRef]
- Jeandet, P.; Formela-Luboińska, M.; Labudda, M.; Morkunas, I. The Role of Sugars in Plant Responses to Stress and Their Regulatory Function during Development. Int. J. Mol. Sci. 2022, 23, 5161. [Google Scholar] [CrossRef]
- Mu, Q.; Zhang, M.; Li, Y.; Feng, F.; Yu, X.; Nie, J. Metabolomic Analysis Reveals the Effect of Insecticide Chlorpyrifos on Rice Plant Metabolism. Metabolites 2022, 12, 1289. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Han, S.; Wang, L.; Li, W. Carbon and Nitrogen Metabolic Regulation in Freshwater Plant Ottelia alismoides in Response to Carbon Limitation: A Metabolite Perspective. Front. Plant Sci. 2022, 13, 962622. [Google Scholar] [CrossRef]
- Wu, C.; Wang, Y.; Sun, H. Targeted and Untargeted Metabolomics Reveals Deep Analysis of Drought Stress Responses in Needles and Roots of Pinus taeda Seedlings. Front. Plant Sci. 2023, 13, 1031466. [Google Scholar] [CrossRef]
- Halabalaki, M.; Bertrand, S.; Stefanou, A.; Gindro, K.; Kostidis, S.; Mikros, E.; Skaltsounis, L.A.; Wolfender, J.-L. Sample Preparation Issues in NMR-Based Plant Metabolomics: Optimisation for Vitis Wood Samples: Sample Preparation Issues in NMR-Based Metabolimics of Vitis Wood. Phytochem. Anal. 2014, 25, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Zainal, B.; Ding, P.; Ismail, I.S.; Saari, N. H NMR Metabolomics Profiling Unveils the Compositional Changes of Hydro-Cooled Rockmelon (Cucumis melo L. Reticulatus Cv Glamour) during Storage Related to in Vitro Antioxidant Activity. Sci. Hortic. 2019, 246, 618–633. [Google Scholar] [CrossRef]
- Mishra, S.; Gogna, N.; Dorai, K. NMR-Based Investigation of the Altered Metabolic Response of Bougainvillea Spectabilis Leaves Exposed to Air Pollution Stress during the Circadian Cycle. Environ. Exp. Bot. 2019, 164, 58–70. [Google Scholar] [CrossRef]
- Ontiveros-Rodríguez, J.C.; Serrano-Contreras, J.I.; Villagómez-Ibarra, J.R.; García-Gutiérrez, H.A.; Gerardo Zepeda-Vallejo, L. A Semi-Targeted NMR-Based Chemical Profiling of Retail Samples of Mexican Gordolobo. J. Pharm. Biomed. Anal. 2022, 212, 114651. [Google Scholar] [CrossRef]
- Dhuli, P.; Rohloff, J.; Strimbeck, G.R. Metabolite Changes in Conifer Buds and Needles during Forced Bud Break in Norway Spruce (Picea abies) and European Silver Fir (Abies alba). Front. Plant Sci. 2014, 5, 706. [Google Scholar] [CrossRef]
- Tajidin, N.E.; Shaari, K.; Maulidiani, M.; Salleh, N.S.; Ketaren, B.R.; Mohamad, M. Metabolite Profiling of Andrographis paniculata (Burm. f.) Nees. Young and Mature Leaves at Different Harvest Ages Using 1H NMR-Based Metabolomics Approach. Sci. Rep. 2019, 9, 16766. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | 1H δ (ppm) * | Multiplicity ** | J (Hz) | Group | 13C δ (ppm) * |
|---|---|---|---|---|---|
| Amino acids | |||||
| Threonine | 1.33 | d | 7.2 | CH3 | 19.8 |
| 4.04 | m | - | CH | n.d. *** | |
| Alanine | 1.49 | d | 7.2 | CH3 | 17.2 |
| 3.70 | m | - | CH | n.d. | |
| GABA | 2.32 | t | 7.1 | CH2(NH2) | 35.5 |
| 1.91 | m | - | CH2 | 25.2 | |
| 3.00 | t | 7.3 | CH2(COOH) | 40.5 | |
| Carbohydrates | |||||
| Fructose | 3.92 | CH | 69.5 | ||
| C | 98.0 | ||||
| C | 101.7 | ||||
| C | 104.4 | ||||
| β-glucose | 4.56 | d | 8.0 | CH(O) | 97.6 |
| 3.19 | dd | 8.0; 9.3 | CH | 75.8 | |
| 3.43 | t | 9.3 | CH | 77.5 | |
| 3.31 | dd | 7.9; 9.3 | CH | 71.2 | |
| 3.37 | m | - | CH | 77.6 | |
| 3.87, 3.69 | m | - | CH2 | 62.4 | |
| α-glucose | 5.17 | d | 3.9 | CH(O) | 93.6 |
| 3.45 | dd | 3.9; 9.5 | CH | 73.3 | |
| 3.62 | t | 9.5 | CH | 74.4 | |
| 3.34 | m | - | CH | 71.3 | |
| 3.82 | m | - | CH | 72.9 | |
| 3.81, 3.72 | m | - | CH2 | 62.3 | |
| Sucrose | 5.41 | d | 3.8 | CH | 93.5 |
| 4.16 | d | 8.6 | CH | 78.4 | |
| Organic acids | |||||
| Succinic acid | 2.52 | s | - | CH2 | 26.6 |
| Shikimic acid | 6.49 | m | - | CH(=) | 131.8 |
| - | C | 137.3 | |||
| 4.35 | m | - | CH | 67.8 | |
| 3.95 | m | - | CH | 68.3 | |
| 3.61 | dd | 4.4; 8.8 | CH | 73.8 | |
| 2.78, 2.19 | m | - | CH2 | 34.1 | |
| Formic acid | 8.48 | s | - | CH | 169.1 |
| Other | |||||
| Choline | 3.21 | s | - | CH3 | 54.9 |
| Formic Acid | Shikimic Acid | Sucrose | α-Glucose | β-Glucose | Fructose | Choline | GABA | Succinic Acid | Alanine | Threonine | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Region 4 | 0.820 b | 33.834 ab | 1.326 a | 13.676 a | 21.588 a | 6.940 a | 3.171 a | 2.157 a | 12.732 a | 2.430 a | 1.327 a |
| Region 3 | 0.888 b | 26.460 b | 1.611 a | 13.069 a | 20.670 a | 12.674 a | 3.786 a | 1.619 a | 16.255 a | 1.739 a | 1.444 a |
| Region 1 | 2.591 a | 35.932 a | 0.586 a | 11.551 a | 18.493 a | 8.247 a | 5.643 a | 1.948 a | 12.308 a | 1.860 a | 0.902 a |
| Region 2 | 0.956 b | 38.954 a | 0.365 a | 13.010 a | 20.425 a | 7.476 a | 3.986 a | 1.671 a | 10.248 a | 1.919 a | 1.220 a |
| Pr > F | <0.0001 | 0.036 | 0.207 | 0.262 | 0.279 | 0.147 | 0.225 | 0.792 | 0.690 | 0.746 | 0.745 |
| Significant | Yes | Yes | No | No | No | No | No | No | No | No | No |
| Site | Code | Geographical Coordinates | Altitude (m) | Species | Tmax (°C) | Tmin (°C) | Pan (mm) | Pollution Level/Sampling Area Type |
|---|---|---|---|---|---|---|---|---|
| Mihaesti | S_1 | 45.043233, 24.248252 | 236 | Spruce | 16.8 | 7.0 | 65.5 | Low-medium, rural, Region 2 |
| Govora | S_2 | 45.072151, 24.205248 | 270 | Spruce | 16.3 | 6.8 | 65.5 | Low-medium, rural, Region 2 |
| Govora | S_3 | 45.072818, 24.195674 | 282 | Fir | 16.3 | 6.8 | 65.5 | Low-medium, rural, Region 2 |
| Govora | S_4 | 45.075123, 24.192590 | 299 | Fir | 16.3 | 6.8 | 65.5 | Low-medium, rural, Region 2 |
| Baile Govora | S_5 | 45.081617, 24.177123 | 329 | Spruce | 16.0 | 6.2 | 65.5 | Low-medium, balneo resort, Region 2 |
| Baile Govora | S_6 | 45.082184, 24.168034 | 392 | Spruce | 16.0 | 6.2 | 65.5 | Low-medium, balneo resort, Region 2 |
| Baile Govora | S_7 | 45.078622, 24.184012 | 307 | Spruce | 16.0 | 6.2 | 65.5 | Low-medium, balneo resort, Region 2 |
| Baile Govora | S_8 | 45.076675, 24.188106 | 300 | Spruce | 16.0 | 6.2 | 65.5 | Low-medium, balneo resort, Region 2 |
| Govora | S_9 | 45.086692, 24.218175 | 272 | Fir | 16.3 | 6.8 | 65.5 | Medium, rural, Region 3 |
| Ocnele Mari | S_10 | 45.08831, 24.29659 | 266 | Spruce | 16.3 | 6.6 | 65.5 | Low-Medium, rural, Region 2 |
| Ocnele Mari | S_11 | 45.086189, 24.302747 | 264 | Spruce | 16.3 | 6.6 | 65.5 | Medium, rural, Region 3 |
| Ocnele Mari | S_12 | 45.081915, 24.309449 | 259 | Spruce | 16.3 | 6.6 | 65.5 | Medium, rural, Region 3 |
| Ocnele Mari | S_13 | 45.078993, 24.311667 | 250 | Spruce | 16.3 | 6.6 | 65.5 | Medium, rural, Region 3 |
| Troian | S_14 | 45.072444, 24.330117 | 246 | Spruce | 16.8 | 7.0 | 65.5 | Medium, rural, Region 3 |
| Vladesti | S_15 | 45.119613, 24.305616 | 292 | Spruce | 16.3 | 6.8 | 65.5 | Medium, rural, Region 3 |
| Vladesti | S_16 | 45.112791, 24.323620 | 278 | Fir | 16.3 | 6.8 | 65.5 | Medium, rural, Region 3 |
| Vladesti | S_17 | 45.113277, 24.322721 | 278 | Spruce | 16.3 | 6.8 | 65.5 | Medium, rural, Region 3 |
| Vladesti | S_18 | 45.127225, 24.271221 | 313 | Fir | 16.3 | 6.8 | 65.5 | Medium, rural, Region 3 |
| Pausesti Maglasi | S_19 | 45.140121, 24.246315 | 338 | Spruce | 15.0 | 7.6 | 59.2 | Medium, rural, Region 3 |
| Olanesti | S_20 | 45.172527, 24.257951 | 378 | Spruce | 15.0 | 7.6 | 59.2 | Low-medium, rural, Region 2 |
| Baile Olanesti | S_21 | 45.203455, 24.241029 | 434 | Spruce | 14.0 | 6.6 | 59.2 | Low-medium, balneo resort, Region 2 |
| Baile Olanesti | S_22 | 45.206098, 24.237576 | 422 | Spruce | 14.0 | 6.6 | 59.2 | Low-medium, balneo resort, Region 2 |
| Pausesti Maglasi | S_23 | 45.152544, 24.248061 | 356 | Spruce | 15.0 | 7.6 | 59.2 | Medium, rural, Region 3 |
| Ramnicu Valcea | S_24 | 45.106805, 24.363972 | 253 | Spruce | 16.8 | 7 | 65.5 | High, urban, Region 4 |
| Ramnicu Valcea | S_25 | 45.109167, 24.363379 | 260 | Spruce | 16.8 | 7 | 65.5 | High, urban, Region 4 |
| Raureni | S_26 | 45.03541, 24.28569 | 220 | Spruce | 16.8 | 7.1 | 65.5 | High, industrial, Region 4 |
| Raureni | S_27 | 45.03541, 24.28569 | 220 | Spruce | 16.8 | 7.1 | 65.5 | High, industrial, Region 4 |
| Cozia National Park | S_28 | 45.29020, 24.41727 | 654 | Spruce | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_29 | 45.29296, 24.41088 | 723 | Spruce | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_30 | 45.29834, 24.40141 | 850 | Fir | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_31 | 45.30348, 24.39652 | 855 | Spruce | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_32 | 45.30817, 24.38673 | 907 | Fir | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_33 | 45.31366, 24.37694 | 950 | Spruce | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_34 | 45.31998, 24.37551 | 1036 | Spruce | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_35 | 45.32211, 24.37143 | 1110 | Spruce | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_36 | 45.32744, 24.37021 | 1160 | Spruce | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_37 | 45.32851, 24.36409 | 1180 | Fir | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_38 | 45.32898, 24.35904 | 1311 | Fir | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_39 | 45.32631, 24.35373 | 1310 | Spruce | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_40 | 45.32079, 24.33799 | 1554 | Spruce | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Cozia National Park | S_41 | 45.32382, 24.34144 | 1488 | Spruce | 7.8 | 0.4 | 59.2 | Low, mountain, Region 1 |
| Malaia | S_42 | 45.35751, 24.01593 | 521 | Spruce | 14.6 | 7.3 | 64.0 | Low-medium, rural, Region 2 |
| Voineasa | S_43 | 45.42373, 23.96654 | 745 | Spruce | 12.3 | 5.3 | 64.0 | Low, mountain resort, Region 1 |
| Voineasa | S_44 | 45.41644, 23.96439 | 671 | Spruce | 12.3 | 5.3 | 64.0 | Low, mountain resort, Region 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miricioiu, M.G.; Ionete, R.E.; Simova, S.; Gerginova, D.; Botoran, O.R. Metabolite Profiling of Conifer Needles: Tracing Pollution and Climate Effects. Int. J. Mol. Sci. 2023, 24, 14986. https://doi.org/10.3390/ijms241914986
Miricioiu MG, Ionete RE, Simova S, Gerginova D, Botoran OR. Metabolite Profiling of Conifer Needles: Tracing Pollution and Climate Effects. International Journal of Molecular Sciences. 2023; 24(19):14986. https://doi.org/10.3390/ijms241914986
Chicago/Turabian StyleMiricioiu, Marius Gheorghe, Roxana Elena Ionete, Svetlana Simova, Dessislava Gerginova, and Oana Romina Botoran. 2023. "Metabolite Profiling of Conifer Needles: Tracing Pollution and Climate Effects" International Journal of Molecular Sciences 24, no. 19: 14986. https://doi.org/10.3390/ijms241914986