

miR-136 Regulates the Proliferation and Adipogenic Differentiation of Adipose-Derived Stromal Vascular Fractions by Targeting HSD17B12
 , ,
, ,
Abstract
:1. Introduction
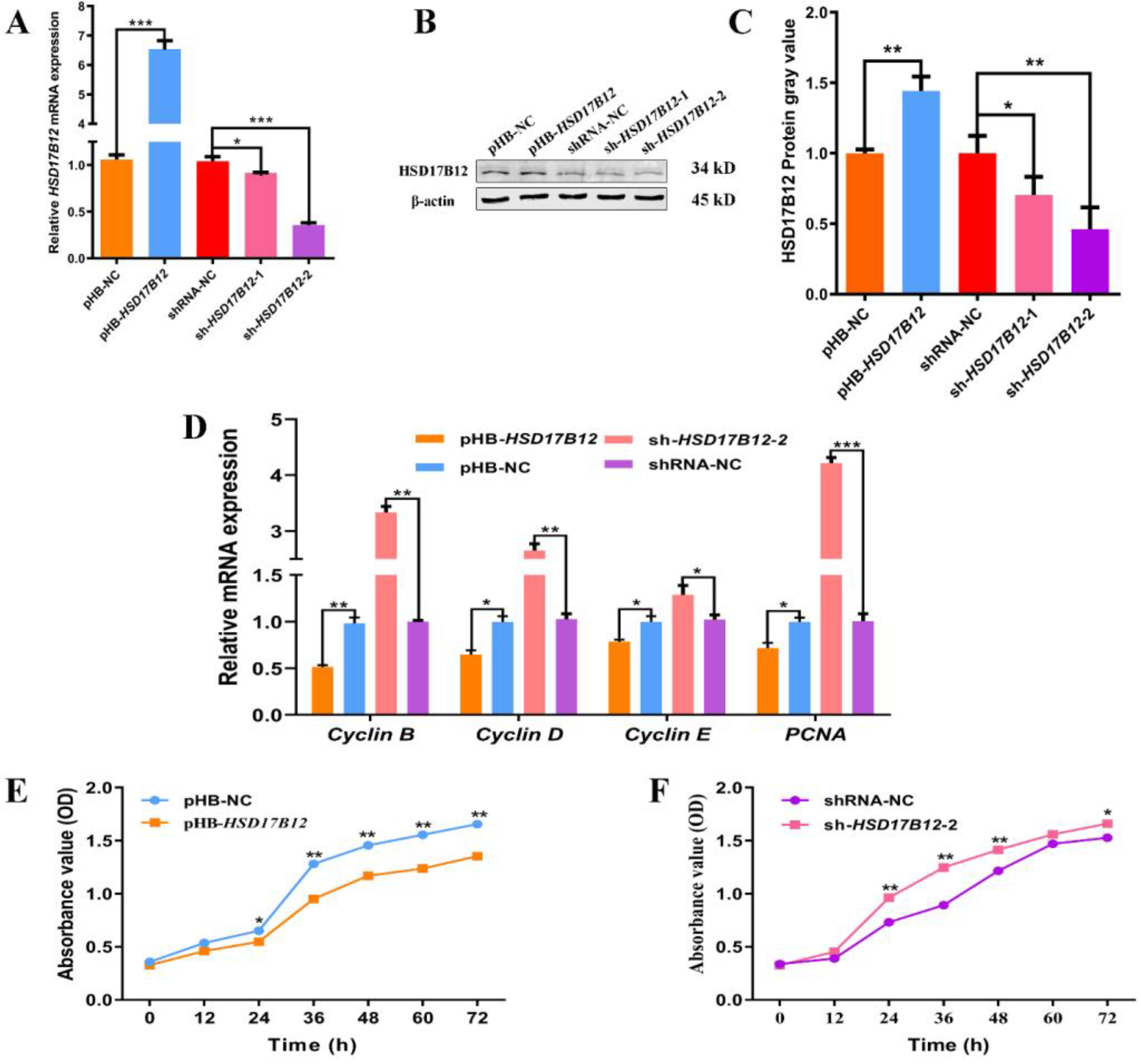
2. Results
2.1. Identification of Ovine SVFs
2.2. miR-136 Targets HSD17B12 3′-UTR
2.3. miR-136 Promotes the Proliferation of Ovine SVFs
2.4. miR-136 Inhibits the Adipogenic Differentiation of Ovine SVFs
2.5. HSD17B12 Suppresses the Proliferation of Ovine SVFs
2.6. HSD17B12 Facilitates the Adipogenic Differentiation of Ovine SVFs
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Isolation and Culture of Ovine SVFs
4.3. Adipogenic Induction and ORO Staining
4.4. Target Gene Prediction and Luciferase Reporter Assays
4.5. Transfection of Mir-136 Mimics and Inhibitors into Ovine Preadipocytes
4.6. Lentiviral Infection
4.7. Quantitative Real-Time PCR (qRT-PCR)
4.8. Cell Count Determination
4.9. Western Blotting
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, Z.; Liu, J.; Han, J.; Yang, B. Association between BMP2 functional polymorphisms and sheep tail type. Animals 2020, 10, 739. [Google Scholar] [CrossRef] [PubMed]
- Bak, R.O.; Mikkelsen, J.G. miRNA sponges: Soaking up miRNAs for regulation of gene expression. Wiley Interdiscip. Rev. RNA 2014, 5, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Heikkinen, L.; Wang, C.; Yang, Y.; Sun, H.; Wong, G. Trends in the development of miRNA bioinformatics tools. Brief. Bioinform. 2019, 20, 1836–1852. [Google Scholar] [CrossRef]
- Fabian, M.R.; Sonenberg, N. The mechanics of miRNA-mediated gene silencing: A look under the hood of miRISC. Nat. Struct. Mol. Biol. 2012, 19, 586–593. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, Y.; Chen, X.; Peng, Y.; Chen, F.; He, Y.; Pang, W.; Yang, G.; Yu, T. MiR-127 attenuates adipogenesis by targeting MAPK4 and HOXC6 in porcine adipocytes. J. Cell. Physiol. 2019, 234, 21838–21850. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Cui, Y.; Dong, X.; Zhang, T.; Xing, H. MicroRNA-130b attenuates dexamethasone-induced increase of lipid accumulation in porcine preadipocytes by suppressing PPAR-γ expression. Oncotarget 2017, 8, 87928–87943. [Google Scholar] [CrossRef]
- Liu, J.; Liang, Y.; Qiao, L.; Xia, D.; Pan, Y.; Liu, W. MiR-128-1-5p regulates differentiation of ovine stromal vascular fraction by targeting the KLF11 5′-UTR. Domest. Anim. Endocrinol. 2022, 80, 106711. [Google Scholar] [CrossRef]
- Zhao, B.; Pan, Y.; Qiao, L.; Liu, J.; Yang, K.; Liang, Y.; Liu, W. miR-301a inhibits adipogenic differentiation of adipose-derived stromal vascular fractions by targeting HOXC8 in sheep. Anim. Sci. J. 2021, 92, e13661. [Google Scholar] [CrossRef]
- Davoli, R.; Gaffo, E.; Zappaterra, M.; Bortoluzzi, S.; Zambonelli, P. Identification of differentially expressed small RNAs and prediction of target genes in Italian Large White pigs with divergent backfat deposition. Anim. Genet. 2018, 49, 205–214. [Google Scholar] [CrossRef]
- Meale, S.J.; Romao, J.M.; He, M.L.; Chaves, A.V.; McAllister, T.A.; Guan, L.L. Effect of diet on microRNA expression in ovine subcutaneous and visceral adipose tissues. J. Anim. Sci. 2014, 92, 3328–3337. [Google Scholar] [CrossRef]
- Rantakari, P.; Lagerbohm, H.; Kaimainen, M.; Suomela, J.P.; Strauss, L.; Sainio, K.; Pakarinen, P.; Poutanen, M. Hydroxysteroid (17{beta}) dehydrogenase 12 is essential for mouse organogenesis and embryonic survival. Endocrinology 2010, 151, 1893–1901. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Meng, Y.; Zhang, J.; Ma, L.; Jiang, L.; Zhang, Y.; Yuan, M.; Ren, A.; Zhu, W.; Li, S.; et al. Functional genetic variant of HSD17B12 in the fatty acid biosynthesis pathway predicts the outcome of colorectal cancer. J. Cell. Mol. Med. 2020, 24, 14160–14170. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, B.; Mazeaud, C.; Baril, M.; Poirier, D.; Sow, A.A.; Chatel-Chaix, L.; Titorenko, V.; Lamarre, D. Very-long-chain fatty acid metabolic capacity of 17-beta-hydroxysteroid dehydrogenase type 12 (HSD17B12) promotes replication of hepatitis C virus and related flaviviruses. Sci. Rep. 2020, 10, 4040. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.A.; Horton, J.D. Identification of two mammalian reductases involved in the two-carbon fatty acyl elongation cascade. J. Biol. Chem. 2003, 278, 7335–7343. [Google Scholar] [CrossRef] [PubMed]
- Nagasaki, S.; Miki, Y.; Akahira, J.; Suzuki, T.; Sasano, H. Transcriptional regulation of 17beta-hydroxysteroid dehydrogenase type 12 by SREBP-1. Mol. Cell. Endocrinol. 2009, 307, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Bellemare, V.; Phaneuf, D.; Luu-The, V. Target deletion of the bifunctional type 12 17β-hydroxysteroid dehydrogenase in mice results in reduction of androgen and estrogen levels in heterozygotes and embryonic lethality in homozygotes. Horm. Mol. Biol. Clin. Investig. 2010, 2, 311–318. [Google Scholar] [CrossRef]
- Bellemare, V.; Laberge, P.; Noël, S.; Tchernof, A.; Luu-The, V. Differential estrogenic 17beta-hydroxysteroid dehydrogenase activity and type 12 17beta-hydroxysteroid dehydrogenase expression levels in preadipocytes and differentiated adipocytes. J. Steroid Biochem. Mol. Biol. 2009, 114, 129–134. [Google Scholar] [CrossRef]
- Nguyen, A.; Guo, J.; Banyard, D.A.; Fadavi, D.; Toranto, J.D.; Wirth, G.A.; Paydar, K.Z.; Evans, G.R.; Widgerow, A.D. Stromal vascular fraction: A regenerative reality? Part 1: Current concepts and review of the literature. J. Plast. Reconstr. Aesthet. Surg. 2016, 69, 170–179. [Google Scholar] [CrossRef]
- Wang, Q.; Pan, Y.; Zhao, B.; Qiao, L.; Liu, J.; Liang, Y.; Liu, W. MiR-33a inhibits the adipogenic differentiation of ovine adipose-derived stromal vascular fraction cells by targeting SIRT6. Domest. Anim. Endocrinol. 2021, 74, 106513. [Google Scholar] [CrossRef]
- Hepler, C.; Vishvanath, L.; Gupta, R.K. Sorting out adipocyte precursors and their role in physiology and disease. Genes Dev. 2017, 31, 127–140. [Google Scholar] [CrossRef]
- Xia, S.F.; Jiang, Y.Y.; Qiu, Y.Y.; Huang, W.; Wang, J. Role of diets and exercise in ameliorating obesity-related hepatic steatosis: Insights at the microRNA-dependent thyroid hormone synthesis and action. Life Sci. 2020, 242, 117182. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Du, J.; Zhang, P.; Zhao, X.; Li, Q.; Jiang, A.; Jiang, D.; Tang, G.; Jiang, Y.; Wang, J.; et al. MicroRNA-125a-5p mediates 3T3-L1 preadipocyte proliferation and differentiation. Molecules 2018, 23, 317. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cai, R.; Tang, G.; Zhang, W.; Pang, W. MiR-146a-5p targeting SMAD4 and TRAF6 inhibits adipogenensis through TGF-β and AKT/mTORC1 signal pathways in porcine intramuscular preadipocytes. J. Anim. Sci. Biotechnol. 2021, 12, 12. [Google Scholar] [CrossRef]
- Saloniemi, T.; Jokela, H.; Strauss, L.; Pakarinen, P.; Poutanen, M. The diversity of sex steroid action: Novel functions of hydroxysteroid (17β) dehydrogenases as revealed by genetically modified mouse models. J. Endocrinol. 2012, 212, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Nagasaki, S.; Suzuki, T.; Miki, Y.; Akahira, J.; Kitada, K.; Ishida, T.; Handa, H.; Ohuchi, N.; Sasano, H. 17Beta-hydroxysteroid dehydrogenase type 12 in human breast carcinoma: A prognostic factor via potential regulation of fatty acid synthesis. Cancer Res. 2009, 69, 1392–1399. [Google Scholar] [CrossRef]
- Yang, Y.; Fang, X.; Yang, R.; Yu, H.; Jiang, P.; Sun, B.; Zhao, Z. MiR-152 regulates apoptosis and triglyceride production in MECs via targeting ACAA2 and HSD17B12 genes. Sci. Rep. 2018, 8, 417. [Google Scholar] [CrossRef]
- Selbach, M.; Schwanhäusser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread changes in protein synthesis induced by microRNAs. Nature 2008, 455, 58–63. [Google Scholar] [CrossRef]
- Xu, Y. MicroRNA-136-3p inhibits glioma tumorigenesis in vitro and in vivo by targeting KLF7. World J. Surg. Oncol. 2020, 18, 169. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Sequences (5′→3′) | Sequences Length (bp) |
|---|---|---|
| HSD17B12-F | GATCTATTTCCGGTGAATTCTGAGGCCTGGTTGAAAGCCAT | 939 |
| HSD17B12-R | ACTAGTATCGATGGATCCTCGCTTAGCTGGCGCATCT |
| Names | Sequences (5′→3′) |
|---|---|
| shRNA-1 F | GATCCGGACAAACTGAACCAGGTTTCTTCAAGAGAGAAACCTGGTTCAGTTTGTCCTTTTTTG |
| shRNA-1 R | AATTCAAAAAAGGACAAACTGAACCAGGTTTCTCTCTTGAAGAAACCTGGTTCAGTTTGTCCG |
| shRNA-2 F | GATCCGGACCAATGGATACTTCATCCTTCAAGAGAGGATGAAGTATCCATTGGTCCTTTTTTG |
| shRNA-2 R | AATTCAAAAAAGGACCAATGGATACTTCATCCTCTCTTGAAGGATGAAGTATCCATTGGTCCG |
| Genes | Sequences (5′→3′) | Sequences Length (bp) |
|---|---|---|
| Cyclin B | F: CGATACTCCGTCTCCAAGCC | 261 |
| R: AGCCAGTCAATCAGGATGGC | ||
| Cyclin D | F: GATGCCAACCTCCTCAACGA | 221 |
| R: GGAAGCGGTCCAGGTAGTTC | ||
| PCNA | F: ATCAGCTCAAGTGGCGTGAA | 231 |
| R: TGCCAAGGTGTCCGCATTAT | ||
| PPARγ | F: ATCTTGACGGGAAAGACGAC | 156 |
| R: AAACTGACACCCCTGGAAGAT | ||
| C/EBPα | F: TCCGTGGACAAGAACAGCAA | 137 |
| R: TCATTGTCACTGGTCAGCTCC | ||
| Adiponectin | F: ATCCCCGGGCTGTACTACTT | 129 |
| R: CTGGTCCACGTTCTGGTTCT | ||
| FABP4 | F: AAACTGGGATGGGAAATCAACC | 109 |
| R: TGCTCTCTCGTAAACTCTGGTAGC | ||
| β-Actin | F: TGATGATATTGCTGCGCTCG | 194 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Che, Y.; Cai, K.; Zhao, B.; Qiao, L.; Pan, Y.; Yang, K.; Liu, W. miR-136 Regulates the Proliferation and Adipogenic Differentiation of Adipose-Derived Stromal Vascular Fractions by Targeting HSD17B12. Int. J. Mol. Sci. 2023, 24, 14892. https://doi.org/10.3390/ijms241914892
Liu J, Che Y, Cai K, Zhao B, Qiao L, Pan Y, Yang K, Liu W. miR-136 Regulates the Proliferation and Adipogenic Differentiation of Adipose-Derived Stromal Vascular Fractions by Targeting HSD17B12. International Journal of Molecular Sciences. 2023; 24(19):14892. https://doi.org/10.3390/ijms241914892
Chicago/Turabian StyleLiu, Jianhua, Yutong Che, Ke Cai, Bishi Zhao, Liying Qiao, Yangyang Pan, Kaijie Yang, and Wenzhong Liu. 2023. "miR-136 Regulates the Proliferation and Adipogenic Differentiation of Adipose-Derived Stromal Vascular Fractions by Targeting HSD17B12" International Journal of Molecular Sciences 24, no. 19: 14892. https://doi.org/10.3390/ijms241914892