
MicroRNA PC-3p-2869 Regulates Antler Growth and Inhibits Proliferation and Migration of Human Osteosarcoma and Chondrosarcoma Cells by Targeting CDK8, EEF1A1, and NTN1

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. MiR-PC-2869 Is Widely Expressed in Antler Tissues and Regulates Antler Cell Proliferation
2.2. Heterologous Expression of miR-PC-2869 Inhibits the Proliferation and Migration of Osteosarcoma and Chondrosarcoma Cells
2.3. Screening of miR-PC-2869 Targets Using the Reporter Library
2.4. miR-PC-2869 Directly Regulates 18 Target Genes
2.5. Cross-Species Regulation of miR-PC-2869 on CDK8, EEF1A1, and NTN1
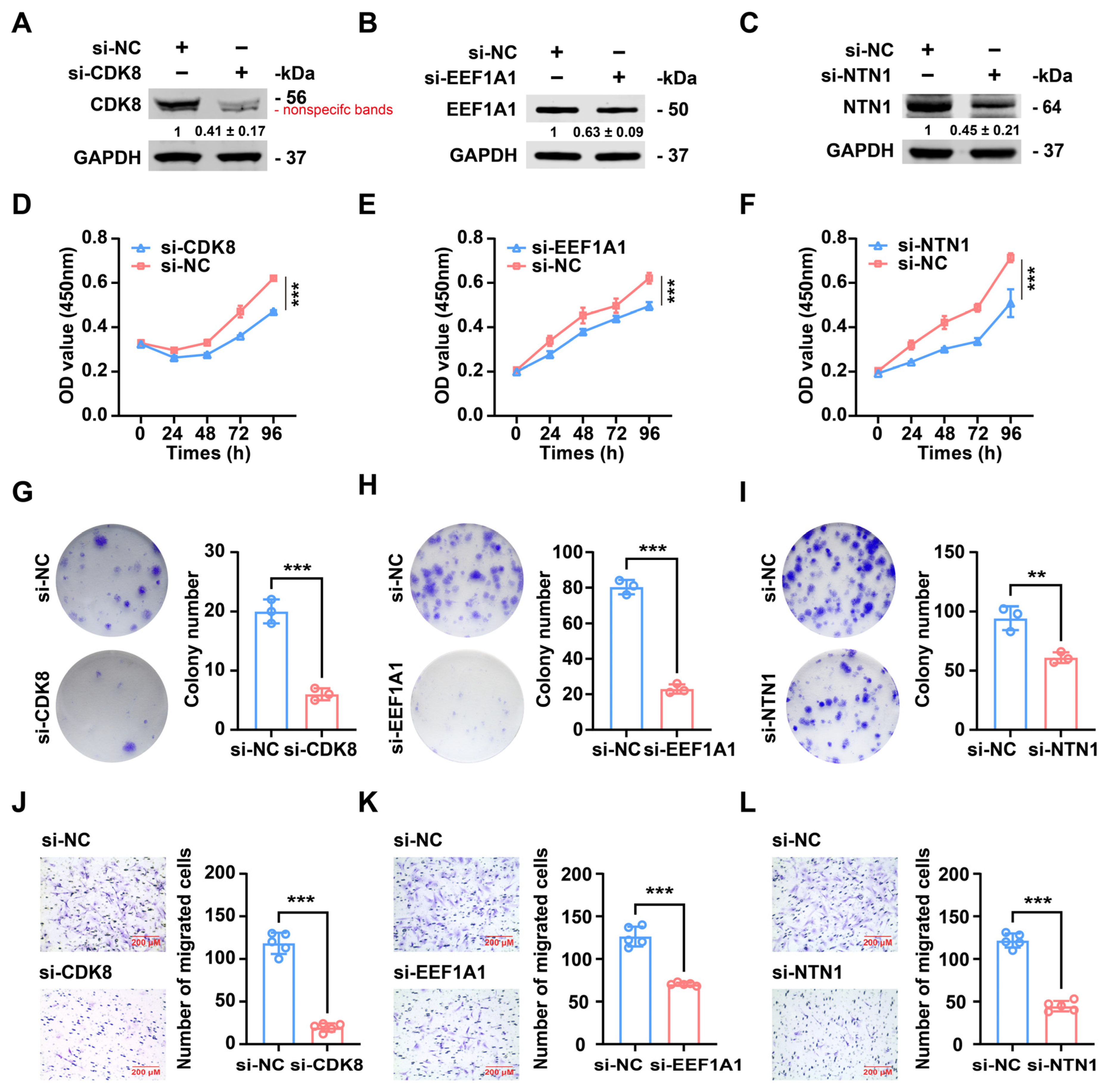
2.6. The Knockdown of CDK8, EEF1A1, or NTN1 Results in Similar Inhibitory Effects on the Proliferation and Migration of Antler Cartilage Cells to the Overexpression of miR-PC-2869
2.7. Silencing of CDK8, EEF1A1, or NTN1 Imitates the Tumor-Suppressor Activity of Heterologous miR-PC-2869 Expression in Osteosarcoma Cells
2.8. Inhibition of CDK8, EEF1A1, or NTN1 Mimics the Tumor-Suppressive Effects of Heterologous miR-PC-2869 Expression in Chondrosarcoma Cells
2.9. MiR-PC-2869 Represses c-Myc and Cyclin D1 Expression by Targeting CDK8, EEF1A1 and NTN1
3. Discussion
4. Materials and Methods
4.1. Tissue Collection and Cell Culture
4.2. Quantitative Real-Time PCR
4.3. Plasmids, miRNA Mimics, and siRNAs
4.4. Cell Transfection
4.5. Cell Proliferation Assay
4.6. Transwell Migration Assay
4.7. Colony Formation Assay
4.8. Dual-Luciferase Reporter Assay
4.9. Western Blot
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Price, J.S.; Allen, S.; Faucheux, C.; Althnaian, T.; Mount, J.G. Deer antlers: A zoological curiosity or the key to understanding organ regeneration in mammals? J. Anat. 2005, 207, 603–618. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, C.; Wang, N.; Li, Z.; Heller, R.; Liu, R.; Zhao, Y.; Han, J.; Pan, X.; Zheng, Z.; et al. Genetic basis of ruminant headgear and rapid antler regeneration. Science 2019, 364, eaav6335. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhao, H.; Liu, Z.; McMahon, C. Deer antler—A novel model for studying organ regeneration in mammals. Int. J. Biochem. Cell Biol. 2014, 56, 111–122. [Google Scholar] [CrossRef]
- Jia, B.; Zhang, L.; Zhang, Y.; Ge, C.; Yang, F.; Du, R.; Ba, H. Integrated analysis of miRNA and mRNA transcriptomic reveals antler growth regulatory network. Mol. Genet. Genom. 2021, 296, 689–703. [Google Scholar] [CrossRef]
- Hu, P.; Wang, T.; Liu, H.; Xu, J.; Wang, L.; Zhao, P.; Xing, X. Full-length transcriptome and microRNA sequencing reveal the specific gene-regulation network of velvet antler in sika deer with extremely different velvet antler weight. Mol. Genet. Genom. 2019, 294, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Papachristou, D.J.; Papavassiliou, A.G. Osteosarcoma and chondrosarcoma: New signaling pathways as targets for novel therapeutic interventions. Int. J. Biochem. Cell Biol. 2007, 39, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Aran, V.; Devalle, S.; Meohas, W.; Heringer, M.; Cunha Caruso, A.; Pinheiro Aguiar, D.; Leite Duarte, M.E.; Moura Neto, V. Osteosarcoma, chondrosarcoma and Ewing sarcoma: Clinical aspects, biomarker discovery and liquid biopsy. Crit. Rev. Oncol. Hematol. 2021, 162, 103340. [Google Scholar] [CrossRef]
- Ferguson, J.L.; Turner, S.P. Bone Cancer: Diagnosis and Treatment Principles. Am. Fam. Physician 2018, 98, 205–213. [Google Scholar]
- Speetjens, F.M.; de Jong, Y.; Gelderblom, H.; Bovée, J.V. Molecular oncogenesis of chondrosarcoma: Impact for targeted treatment. Curr. Opin. Oncol. 2016, 28, 314–322. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yang, F.; Zhao, M.; Xiao, H.; Chen, Y.; Liu, X.; Zheng, D. Antler-derived microRNA PC-5p-1090 inhibits HCC cell proliferation, migration, and invasion by targeting MARCKS, SMARCAD1, and SOX9. Funct. Integr. Genom. 2023, 23, 156. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Inoue, A.; Seers, T.; Hato, Y.; Igarashi, A.; Toyama, T.; Taganov, K.D.; Boldin, M.P.; Asahara, H. Identification of targets of tumor suppressor microRNA-34a using a reporter library system. Proc. Natl. Acad. Sci. USA 2017, 114, 3927–3932. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.X.; Liu, X.D.; Yang, X.G.; Liu, Y.H.; Pi, X.M.; Liu, Q.Z.; Zheng, D. Deep sequencing identifies conserved and novel microRNAs from antlers cartilage of Chinese red deer (Cervus elaphus). Genes Genom. 2015, 37, 419–427. [Google Scholar] [CrossRef]
- Zhang, J.F.; Zhang, J.S.; Zhao, Z.H.; Yang, P.B.; Ji, S.F.; Li, N.; Shi, Q.D.; Tan, J.; Xu, X.; Xu, C.B.; et al. MicroRNA-770 affects proliferation and cell cycle transition by directly targeting CDK8 in glioma. Cancer Cell Int. 2018, 18, 195. [Google Scholar] [CrossRef]
- Huang, J.; Zheng, C.; Shao, J.; Chen, L.; Liu, X.; Shao, J. Overexpression of eEF1A1 regulates G1-phase progression to promote HCC proliferation through the STAT1-cyclin D1 pathway. Biochem. Biophys. Res. Commun. 2017, 494, 542–549. [Google Scholar] [CrossRef]
- Yin, K.; Shang, M.; Dang, S.; Wang, L.; Xia, Y.; Cui, L.; Fan, X.; Qu, J.; Chen, J.; Xu, Z. Netrin-1 induces the proliferation of gastric cancer cells via the ERK/MAPK signaling pathway and FAK activation. Oncol. Rep. 2018, 40, 2325–2333. [Google Scholar] [CrossRef] [PubMed]
- Colitti, M.; Allen, S.P.; Price, J.S. Programmed cell death in the regenerating deer antler. J. Anat. 2005, 207, 339–351. [Google Scholar] [CrossRef]
- Matich, J.; Basford Nicholson, L.F.; Barling, P.M. Mitotic activity in the growing red deer antler. Cell Biol. Int. 2003, 27, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Li, Q.; Lin, D.; Zong, X.; Luo, X.; Yang, M.; Yue, X.; Ma, S. Peptidomic analysis of pilose antler and its inhibitory effect on triple-negative breast cancer at multiple sites. Food. Funct. 2020, 11, 7481–7494. [Google Scholar] [CrossRef]
- Chonco, L.; Landete-Castillejos, T.; Serrano-Heras, G.; Serrano, M.P.; Pérez-Barbería, F.J.; González-Armesto, C.; García, A.; de Cabo, C.; Lorenzo, J.M.; Li, C.; et al. Anti-tumour activity of deer growing antlers and its potential applications in the treatment of malignant gliomas. Sci. Rep. 2021, 11, 42. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wei, J.; Zhang, S.; Zhang, X. Shrimp miR-S8 Suppresses the Stemness of Human Melanoma Stem-like Cells by Targeting the Transcription Factor YB-1. Cancer Res. 2017, 77, 5543–5553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhang, X. Shrimp miRNA suppresses the stemness of human cancer stem cells via the PIN1 pathway. Faseb. J. 2019, 33, 10767–10779. [Google Scholar] [CrossRef] [Green Version]
- Chin, A.R.; Fong, M.Y.; Somlo, G.; Wu, J.; Swiderski, P.; Wu, X.; Wang, S.E. Cross-kingdom inhibition of breast cancer growth by plant miR159. Cell Res. 2016, 26, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Firestein, R.; Bass, A.J.; Kim, S.Y.; Dunn, I.F.; Silver, S.J.; Guney, I.; Freed, E.; Ligon, A.H.; Vena, N.; Ogino, S.; et al. CDK8 is a colorectal cancer oncogene that regulates beta-catenin activity. Nature 2008, 455, 547–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Jin, H.; Hu, J.; Li, X.; Ruan, H.; Xu, H.; Wei, L.; Dong, W.; Teng, F.; Gu, J.; et al. COL4A1 promotes the growth and metastasis of hepatocellular carcinoma cells by activating FAK-Src signaling. J. Exp. Clin. Cancer Res. 2020, 39, 148. [Google Scholar] [CrossRef]
- Fan, A.H.; Zhao, X.; Liu, H.; Li, D.; Guo, T.; Zhang, J.; Duan, L.; Cheng, H.; Nie, Y.; Fan, D.; et al. eEF1A1 promotes colorectal cancer progression and predicts poor prognosis of patients. Cancer Med. 2023, 12, 513–524. [Google Scholar] [CrossRef]
- Inoue, R.; Hirohashi, Y.; Kitamura, H.; Nishida, S.; Murai, A.; Takaya, A.; Yamamoto, E.; Matsuki, M.; Tanaka, T.; Kubo, T.; et al. GRIK2 has a role in the maintenance of urothelial carcinoma stem-like cells, and its expression is associated with poorer prognosis. Oncotarget 2017, 8, 28826–28839. [Google Scholar] [CrossRef] [Green Version]
- Yin, K.; Wang, L.; Xia, Y.; Dang, S.; Zhang, X.; He, Z.; Xu, J.; Shang, M.; Xu, Z. Netrin-1 promotes cell neural invasion in gastric cancer via its receptor neogenin. J. Cancer 2019, 10, 3197–3207. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Liu, S.; Shan, M.; Hagenaars, S.C.; Mesker, W.E.; Cohen, D.; Wang, L.; Zheng, Z.; Devilee, P.; Tollenaar, R.; et al. RNF12 is regulated by AKT phosphorylation and promotes TGF-β driven breast cancer metastasis. Cell Death Dis. 2022, 13, 44. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, S.; Gao, D.; Yu, K.; Chen, H.; Huang, Y.; Li, M.; Zhang, J.; Guo, K. Nucleotide sugar transporter SLC35A2 is involved in promoting hepatocellular carcinoma metastasis by regulating cellular glycosylation. Cell Oncol. 2023, 46, 283–297. [Google Scholar] [CrossRef]
- Huang, T.; Zhou, Y.; Zhang, J.; Wong, C.C.; Li, W.; Kwan, J.S.H.; Yang, R.; Chan, A.K.Y.; Dong, Y.; Wu, F.; et al. SRGAP1, a crucial target of miR-340 and miR-124, functions as a potential oncogene in gastric tumorigenesis. Oncogene 2018, 37, 1159–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elster, D.; Tollot, M.; Schlegelmilch, K.; Ori, A.; Rosenwald, A.; Sahai, E.; von Eyss, B. TRPS1 shapes YAP/TEAD-dependent transcription in breast cancer cells. Nat. Commun. 2018, 9, 3115. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yuan, R.; Wen, C.; Liu, T.; Feng, Q.; Deng, X.; Du, Y.; Peng, X. E3 ubiquitin ligase UBR5 promotes pancreatic cancer growth and aerobic glycolysis by downregulating FBP1 via destabilization of C/EBPα. Oncogene 2021, 40, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Menzl, I.; Witalisz-Siepracka, A.; Sexl, V. CDK8-Novel Therapeutic Opportunities. Pharmaceuticals 2019, 12, 92. [Google Scholar] [CrossRef] [Green Version]
- Morris, E.J.; Ji, J.Y.; Yang, F.; Di Stefano, L.; Herr, A.; Moon, N.S.; Kwon, E.J.; Haigis, K.M.; Näär, A.M.; Dyson, N.J. E2F1 represses beta-catenin transcription and is antagonized by both pRB and CDK8. Nature 2008, 455, 552–556. [Google Scholar] [CrossRef] [Green Version]
- Adler, A.S.; McCleland, M.L.; Truong, T.; Lau, S.; Modrusan, Z.; Soukup, T.M.; Roose-Girma, M.; Blackwood, E.M.; Firestein, R. CDK8 maintains tumor dedifferentiation and embryonic stem cell pluripotency. Cancer Res. 2012, 72, 2129–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novosylna, O.; Doyle, A.; Vlasenko, D.; Murphy, M.; Negrutskii, B.; El’skaya, A. Comparison of the ability of mammalian eEF1A1 and its oncogenic variant eEF1A2 to interact with actin and calmodulin. Biol. Chem. 2017, 398, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Akram, Z.; Ahmed, I.; Mack, H.; Kaur, R.; Silva, R.C.; Castilho, B.A.; Friant, S.; Sattlegger, E.; Munn, A.L. Yeast as a Model to Understand Actin-Mediated Cellular Functions in Mammals-Illustrated with Four Actin Cytoskeleton Proteins. Cells 2020, 9, 672. [Google Scholar] [CrossRef] [Green Version]
- Qi, Q.; Li, D.Y.; Luo, H.R.; Guan, K.L.; Ye, K. Netrin-1 exerts oncogenic activities through enhancing Yes-associated protein stability. Proc. Natl. Acad. Sci. USA 2015, 112, 7255–7260. [Google Scholar] [CrossRef] [Green Version]
- Dumartin, L.; Quemener, C.; Laklai, H.; Herbert, J.; Bicknell, R.; Bousquet, C.; Pyronnet, S.; Castronovo, V.; Schilling, M.K.; Bikfalvi, A.; et al. Netrin-1 mediates early events in pancreatic adenocarcinoma progression, acting on tumor and endothelial cells. Gastroenterology 2010, 138, 1595–1606.e1–e8. [Google Scholar] [CrossRef]
- Ylivinkka, I.; Sihto, H.; Tynninen, O.; Hu, Y.; Laakso, A.; Kivisaari, R.; Laakkonen, P.; Keski-Oja, J.; Hyytiäinen, M. Motility of glioblastoma cells is driven by netrin-1 induced gain of stemness. J. Exp. Clin. Cancer Res. 2017, 36, 9. [Google Scholar] [CrossRef] [Green Version]
- Boussouar, A.; Tortereau, A.; Manceau, A.; Paradisi, A.; Gadot, N.; Vial, J.; Neves, D.; Larue, L.; Battistella, M.; Leboeuf, C.; et al. Netrin-1 and Its Receptor DCC Are Causally Implicated in Melanoma Progression. Cancer Res. 2020, 80, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Clark, D.E.; Lord, E.A.; Stanton, J.A.; Suttie, J.M. Sampling technique to discriminate the different tissue layers of growing antler tips for gene discovery. Anat. Rec. 2002, 268, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Suttie, J.M. Tissue collection methods for antler research. Eur. J. Morphol. 2003, 41, 23–30. [Google Scholar] [CrossRef] [PubMed]
- McGeary, S.E.; Lin, K.S.; Shi, C.Y.; Pham, T.M.; Bisaria, N.; Kelley, G.M.; Bartel, D.P. The biochemical basis of microRNA targeting efficacy. Science 2019, 366, eaav1741. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; Wu, J.; Zhao, M.; Zheng, H.; Suo, J.; Liu, X.; Zheng, D. MicroRNA PC-3p-2869 Regulates Antler Growth and Inhibits Proliferation and Migration of Human Osteosarcoma and Chondrosarcoma Cells by Targeting CDK8, EEF1A1, and NTN1. Int. J. Mol. Sci. 2023, 24, 10840. https://doi.org/10.3390/ijms241310840
Yang F, Wu J, Zhao M, Zheng H, Suo J, Liu X, Zheng D. MicroRNA PC-3p-2869 Regulates Antler Growth and Inhibits Proliferation and Migration of Human Osteosarcoma and Chondrosarcoma Cells by Targeting CDK8, EEF1A1, and NTN1. International Journal of Molecular Sciences. 2023; 24(13):10840. https://doi.org/10.3390/ijms241310840
Chicago/Turabian StyleYang, Fan, Jin Wu, Mindie Zhao, Han Zheng, Jingyuan Suo, Xuedong Liu, and Dong Zheng. 2023. "MicroRNA PC-3p-2869 Regulates Antler Growth and Inhibits Proliferation and Migration of Human Osteosarcoma and Chondrosarcoma Cells by Targeting CDK8, EEF1A1, and NTN1" International Journal of Molecular Sciences 24, no. 13: 10840. https://doi.org/10.3390/ijms241310840