
Pathomechanism of Pruritus in Psoriasis and Atopic Dermatitis: Novel Approaches, Similarities and Differences
 , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Data Sources and Study Selection
3. Results
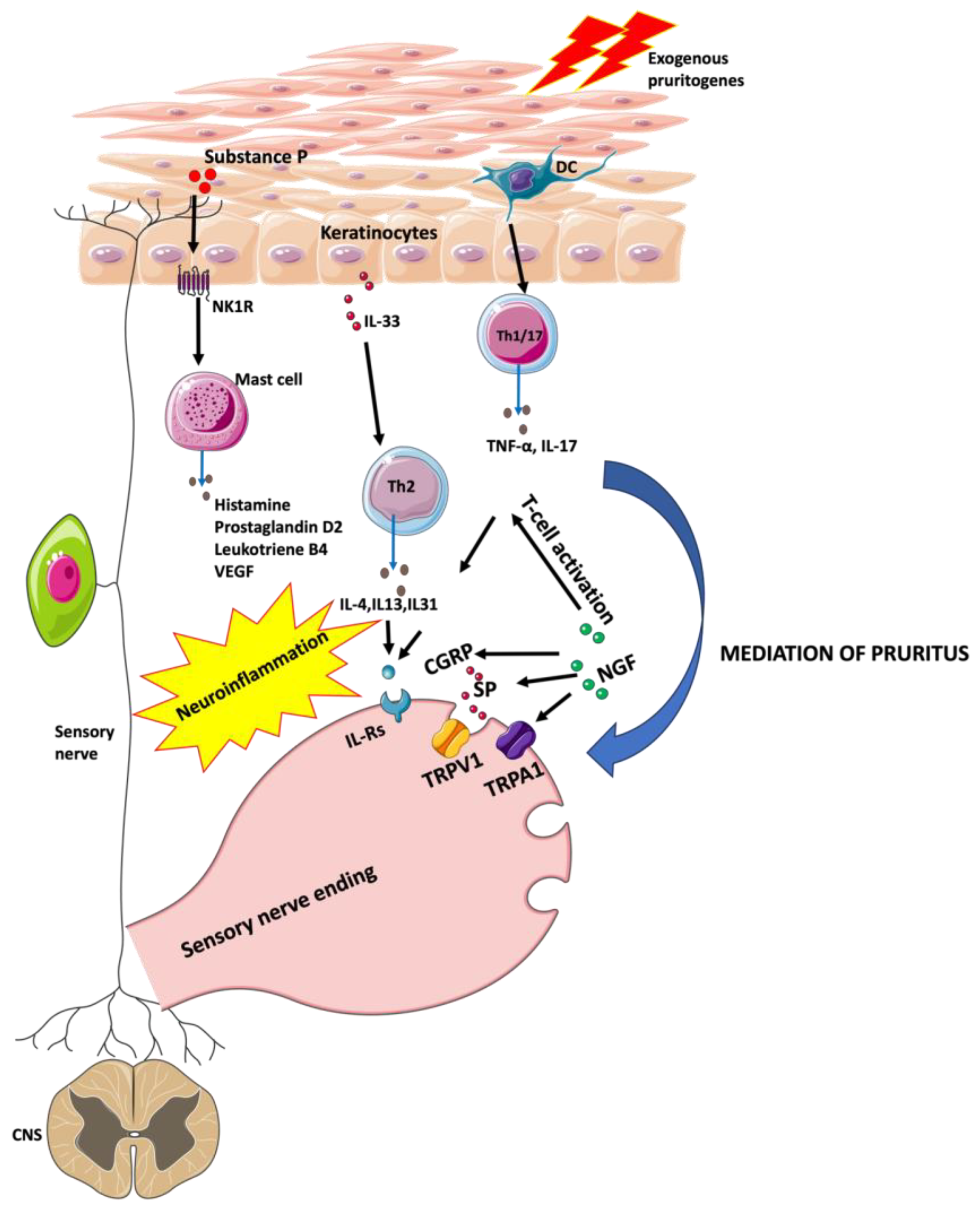
3.1. Pruritus in Psoriasis
3.1.1. Chemokines
3.1.2. Interleukins
3.1.3. Substance P and Neurokinin 1 Receptor
3.1.4. CD26/Dipeptidyl Peptidase IV Enzyme Activity
3.1.5. Transient Receptor Potential (TRP) Channels
3.1.6. Nerve Growth Factor
3.1.7. Opioid Receptors
3.1.8. Histamine
3.2. Pruritus in Atopic Dermatitis
3.2.1. Thymic Stromal Lymphopoietin (TSLP)
3.2.2. TRP Cation Channels
3.2.3. Histidine Decarboxylase
3.2.4. Role of the Microbiota
3.2.5. Artemin
3.2.6. Mas-Related G-Protein Coupled Receptors (MRGPR)
3.2.7. Interleukins
3.3. Differences and Similarities in Pruritus in PSO and AD
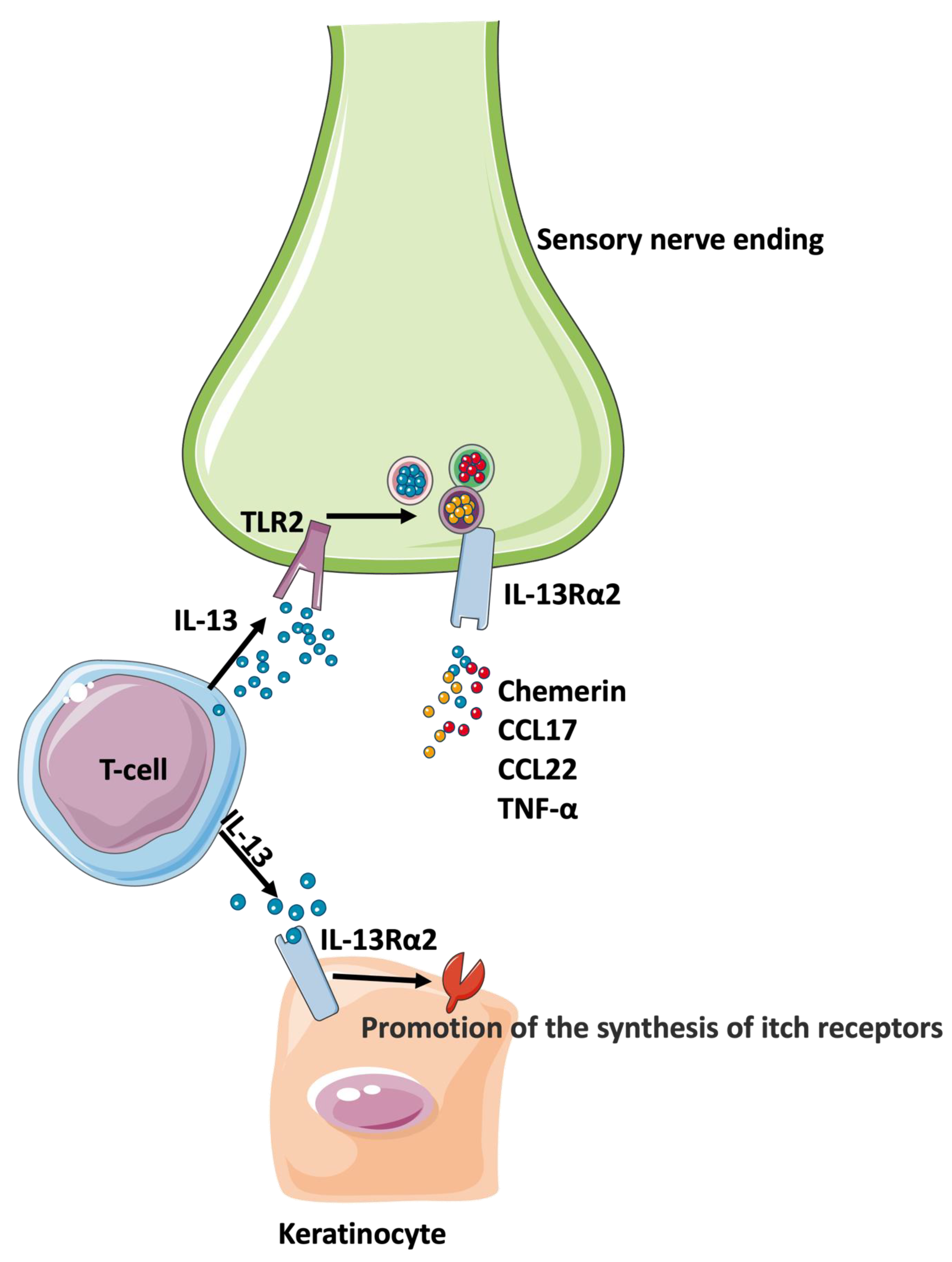
Interleukin 13 Receptor Alpha 2 (IL-13Rα2)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | atopic dermatitis |
| CCL | C-C Motif Chemokine Ligand (CCL) CGRP- calcitonin gene-related peptide |
| DDP IV | dipeptidyl peptidase IV |
| GDNF | glial cell line-derived neurotrophic factor |
| GM-CSF | granulocyte-macrophage colony-stimulating factor |
| GPCRs | G protein-coupled receptors |
| H1R | histamine-1 receptor |
| HDC | histidine decarboxylase |
| IL-1β | interleukin-1β |
| IL-4 | interleukin-4 |
| IL-8 | interleukin-8 |
| IL-12 | interleukin-12 |
| IL-13Rα2 | interleukin 13 receptor alpha 2 |
| IL-17 | interleukin-17 |
| IL-19 | interleukin-19 |
| IL-20 | interleukin-20 |
| IL-23 | interleukin-13 |
| IL-26 | interleukin-26 |
| IL-31 | interleukin-31 |
| IL36A | interleukin-36 alpha |
| IL36G | interleukin-36 gamma |
| ISS | itch severity scale |
| JAK | the Janus kinases |
| LTA | lipoteichoic acid |
| NB-UVB | narrow-band ultraviolet radiation |
| NK1R | neurokinin 1 receptor |
| OPRK | κ-opioid receptor |
| OPRM | μ-opioid receptor |
| PAR-2 | protease-activated receptor 2 |
| pDC | plasmacytoid dendritic cells |
| P-NRS | Pruritus–Numerical Rating Scale |
| PSO | psoriasis |
| S. aureus | Staphylococcus aureus |
| SP | substance P |
| TACR-1 | tachykinin receptor 1 |
| Th1 | T helper cell type 1 |
| Th17 | T helper cell type 17 |
| TNF-α | tumor necrosis factor alpha |
| TLR2 | Toll-like receptor 2 |
| TRP | transient receptor potential |
| TRPV-1 | TRP vanilloid 1 |
| TSLP | thymic stromal lymphopoietin |
| MRGPR | Mas-related G-protein coupled receptors |
| VAS | Visual Analogue Scale |
References
- Armstrong, A.W.; Mehta, M.D.; Schupp, C.W.; Gondo, G.C.; Bell, S.J.; Griffiths, C.E.M. Psoriasis Prevalence in Adults in the United States. JAMA Dermatol. 2021, 157, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, J.; Grewal, S.; Langan, S.M.; Mehta, N.N.; Ogdie, A.; Van Voorhees, A.S.; Gelfand, J.M. Psoriasis and comorbid diseases: Epidemiology. J. Am. Acad. Dermatol. 2017, 76, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Yeung, H.; Takeshita, J.; Mehta, N.N.; Kimmel, S.E.; Ogdie, A.; Margolis, D.J.; Shin, D.B.; Attor, R.; Troxel, A.B.; Gelfand, J.M. Psoriasis severity and the prevalence of major medical comorbidity: A population-based study. JAMA Dermatol. 2013, 149, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.W.; Read, C. Pathophysiology, Clinical Presentation, and Treatment of Psoriasis: A Review. JAMA 2020, 323, 1945–1960. [Google Scholar] [CrossRef]
- Alwan, W.; Nestle, F.O. Pathogenesis and treatment of psoriasis: Exploiting pathophysiological pathways for precision medicine. Clin. Exp. Rheumatol. 2015, 33, S2–S6. [Google Scholar]
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Nair, R.P.; Stuart, P.E.; Nistor, I.; Hiremagalore, R.; Chia, N.V.; Jenisch, S.; Weichenthal, M.; Abecasis, G.R.; Lim, H.W.; Christophers, E.; et al. Sequence and Haplotype Analysis Supports HLA-C as the Psoriasis Susceptibility 1 Gene. Am. J. Hum. Genet. 2006, 78, 827–851. [Google Scholar] [CrossRef]
- Menter, A.; Korman, N.J.; Elmets, C.A.; Feldman, S.R.; Gelfand, J.M.; Gordon, K.B.; Gottlieb, A.; Koo, J.Y.; Lebwohl, M.; Lim, H.W.; et al. Guidelines of care for the management of psoriasis and psoriatic arthritis: Section 5. Guidelines of care for the treatment of psoriasis with phototherapy and photochemotherapy. J. Am. Acad. Dermatol. 2010, 62, 114–135. [Google Scholar] [CrossRef]
- Lebwohl, M.; Drake, L.; Menter, A.; Koo, J.; Gottlieb, A.B.; Zanolli, M.; Young, M.; McClelland, P. Consensus conference: Acitretin in combination with UVB or PUVA in the treatment of psoriasis. J. Am. Acad. Dermatol. 2001, 45, 544–553. [Google Scholar] [CrossRef]
- Menter, A.; Korman, N.J.; Elmets, C.A.; Feldman, S.R.; Gelfand, J.M.; Gordon, K.B.; Gottlieb, A.B.; Koo, J.Y.; Lebwohl, M.; Lim, H.W.; et al. Guidelines of care for the management of psoriasis and psoriatic arthritis: Section 4. Guidelines of care for the management and treatment of psoriasis with traditional systemic agents. J. Am. Acad. Dermatol. 2009, 61, 451–485. [Google Scholar] [CrossRef]
- Saurat, J.-H.; Stingl, G.; Dubertret, L.; Papp, K.A.; Langley, R.G.; Ortonne, J.-P.; Unnebrink, K.; Kaul, M.; Camez, A.; CHAMPION Study Investigators. Efficacy and safety results from the randomized controlled comparative study of adalimumab vs. methotrexate vs. placebo in patients with psoriasis (CHAMPION). Br. J. Dermatol. 2008, 158, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Reich, K.; O Nestle, F.; Papp, K.; Ortonne, J.-P.; Evans, R.; Guzzo, C.; Li, S.; Dooley, L.T.; Griffiths, C.E.; EXPRESS study investigators. Infliximab induction and maintenance therapy for moderate-to-severe psoriasis: A phase III, multicentre, double-blind trial. Lancet 2005, 366, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Torii, H.; Nakagawa, H.; Japanese Infliximab Study Ivestigators. Infliximab monotherapy in japanese patients with moder-ate-to-severe plaque psoriasis and psoriatic arthritis. A randomized, double-blind, placebo-controlled multicenter trial. J. Dermatol. Sci. 2010, 59, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Gordon, K.B.; Blauvelt, A.; Papp, K.A.; Langley, R.G.; Luger, T.; Ohtsuki, M.; Reich, K.; Amato, D.; Ball, S.G.; Braun, D.K.; et al. Phase 3 Trials of Ixekizumab in Moderate-to-Severe Plaque Psoriasis. New Engl. J. Med. 2016, 375, 345–356. [Google Scholar] [CrossRef]
- Lebwohl, M.; Strober, B.; Menter, A.; Gordon, K.; Weglowska, J.; Puig, L.; Papp, K.; Spelman, L.; Toth, D.; Kerdel, F.; et al. Phase 3 Studies Comparing Brodalumab with Ustekinumab in Psoriasis. New Engl. J. Med. 2015, 373, 1318–1328. [Google Scholar] [CrossRef]
- Langley, R.G.; Elewski, B.E.; Lebwohl, M.; Reich, K.; Griffiths, C.E.; Papp, K.; Puig, L.; Nakagawa, H.; Spelman, L.; Sigurgeirsson, B.; et al. Secukinumab in Plaque Psoriasis—Results of Two Phase 3 Trials. New Engl. J. Med. 2014, 371, 326–338. [Google Scholar] [CrossRef]
- Blauvelt, A.; Lomaga, M.; Burge, R.; Zhu, B.; Shen, W.; Shrom, D.; Dossenbach, M.; Pinter, A. Greater Cumulative Benefits from Ixekizumab versus Ustekinumab Treatment over 52 Weeks for Patients with Moder-ate-to-Severe Psoriasis in a Randomized, Double-Blinded Phase 3b Clinical Trial. J. Dermatol. Treat. 2019, 31, 141–146. [Google Scholar] [CrossRef]
- Dogra, S.; Jain, A.; Kanwar, A. Efficacy and Safety of Acitretin in Three Fixed Doses of 25, 35 and 50 mg in Adult Patients with Severe Plaque Type Psoriasis: A Randomized, Double Blind, Parallel Group, Dose Ranging Study. J. Eur. Acad. Dermatol. Venereol. 2013, 27, e305–e311. [Google Scholar] [CrossRef]
- Montes-Torres, A.; Aparicio, G.; Rivera, R.; Vilarrasa, E.; Marcellán, M.; Notario, J.; Soria, C.; Belinchón, I.; de la Cueva, P.; Ferrán, M.; et al. Safety and Effectiveness of Conventional Systemic Therapy and Biological Drugs in Patients with Moderate to Severe Psoriasis and HIV Infection: A Retrospective Multicenter Study. J. Dermatol. Treat. 2019, 30, 461–465. [Google Scholar] [CrossRef]
- Warren, R.B.; Mrowietz, U.; von Kiedrowski, R.; Niesmann, J.; Wilsmann-Theis, D.; Ghoreschi, K.; Zschocke, I.; Falk, T.M.; Blödorn-Schlicht, N.; Reich, K. An intensified dosing schedule of subcutaneous methotrexate in patients with moderate to severe plaque-type psoriasis (METOP): A 52 week, multicentre, randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 389, 528–537. [Google Scholar] [CrossRef]
- Gold, L.S.; Lebwohl, M.G.; Sugarman, J.L.; Pariser, D.M.; Lin, T.; Martin, G.; Pillai, R.; Israel, R.; Ramakrishna, T. Safety and efficacy of a fixed combination of halobetasol and tazarotene in the treatment of moderate-to-severe plaque psoriasis: Results of 2 phase 3 randomized controlled trials. J. Am. Acad. Dermatol. 2018, 79, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.Y.; Lowe, N.J.; Lew-Kaya, D.A.; Vasilopoulos, A.I.; Lue, J.C.; Sefton, J.; Gibson, J.R. Tazarotene plus UVB phototherapy in the treatment of psoriasis. J. Am. Acad. Dermatol. 2000, 43, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Dayal, S.; Kaura, R.; Sahu, P.; Jain, V.K. Tazarotene gel with narrow-band UVB phototherapy: A synergistic combination in psoriasis. An. Bras. De. Dermatol. 2018, 93, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Deckers, I.A.G.; McLean, S.; Linssen, S.; Mommers, M.; van Schayck, C.P.; Sheikh, A. Investigating international time trends in the incidence and prevalence of atopic eczema 1990-2010: A systematic review of epidemiological studies. PLoS ONE 2012, 7, e39803. [Google Scholar] [CrossRef]
- Williams, H.; Stewart, A.; von Mutius, E.; Cookson, W.; Anderson, H.R. Is eczema really on the increase worldwide? J. Allergy Clin. Immunol. 2008, 121, 947–954. [Google Scholar]
- Garmhausen, D.; Hagemann, T.; Bieber, T.; Dimitriou, I.; Fimmers, R.; Diepgen, T.; Novak, N. Characterization of different courses of atopic dermatitis in adolescent and adult patients. Allergy 2013, 68, 498–506. [Google Scholar] [CrossRef]
- Ramirez, F.D.; Chen, S.; Langan, S.M.; Prather, A.A.; McCulloch, C.E.; Kidd, S.A.; Cabana, M.D.; Chren, M.-M.; Abuabara, K. Association of Atopic Dermatitis with Sleep Quality in Children. JAMA Pediatr. 2019, 173, e190025–e1900252019. [Google Scholar] [CrossRef]
- Ravnborg, N.; Ambikaibalan, D.; Agnihotri, G.; Price, S.; Rastogi, S.; Patel, K.R.; Singam, V.; Andersen, Y.; Halling, A.-S.; Silverberg, J.I.; et al. Prevalence of asthma in patients with atopic dermatitis: A systematic review and meta-analysis. J. Am. Acad. Dermatol. 2021, 84, 471–478. [Google Scholar] [CrossRef]
- Drucker, A.M. Atopic dermatitis: Burden of illness, quality of life, and associated complications. Allergy Asthma Proc. 2017, 38, 3–8. [Google Scholar] [CrossRef]
- Wollenberg, A.; Kinberger, M.; Arents, B.; Aszodi, N.; Avila Valle, G.; Barbarot, S.; Bieber, T.; Brough, H.A.; Calzavara Pinton, P.; Christen-Zach, S.; et al. European guideline (EuroGuiDerm) on atopic eczema: Part I—Systemic therapy. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 1409–1431. [Google Scholar] [CrossRef]
- Megna, M.; Napolitano, M.; Patruno, C.; Villani, A.; Balato, A.; Monfrecola, G.; Ayala, F.; Balato, N. Systemic Treatment of Adult Atopic Dermatitis: A Review. Dermatol. Ther. 2017, 7, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Wollenberg, A.; Thomsen, S.F.; Lacour, J.-P.; Jaumont, X.; Lazarewicz, S. Targeting immunoglobulin E in atopic dermatitis: A review of the existing evidence. World Allergy Organ. J. 2021, 14, 100519. [Google Scholar] [CrossRef] [PubMed]
- Wollenberg, A.; Blauvelt, A.; Guttman-Yassky, E.; Worm, M.; Lynde, C.; Lacour, J.P.; Spelman, L.; Katoh, N.; Saeki, H.; Poulin, Y.; et al. Tralokinumab for moderate-to-severe atopic dermatitis: Results from two 52-week, randomized, double-blind, multicentre, placebo-controlled phase iii trials (ecztra 1 and ecztra 2). Br. J. Dermatol. 2021, 184, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Kabashima, K.; Matsumura, T.; Komazaki, H.; Kawashima, M. Trial of Nemolizumab and Topical Agents for Atopic Dermatitis with Pruritus. New Engl. J. Med. 2020, 383, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Guttman-Yassky, E.; Brunner, P.M.; Neumann, A.U.; Khattri, S.; Pavel, A.B.; Malik, K.; Singer, G.K.; Baum, D.; Gilleaudeau, P.; Sullivan-Whalen, M.; et al. Efficacy and safety of fezakinumab (an IL-22 monoclonal antibody) in adults with moderate-to-severe atopic dermatitis inadequately controlled by conventional treatments: A randomized, double-blind, phase 2a trial. J. Am. Acad. Dermatol. 2018, 78, 872–881.e6. [Google Scholar] [CrossRef]
- Simpson, E.L.; Parnes, J.R.; She, D.; Crouch, S.; Rees, W.; Mo, M.; van der Merwe, R. Tezepelumab, an anti–thymic stromal lymphopoietin monoclonal antibody, in the treatment of moderate to severe atopic dermatitis: A randomized phase 2a clinical trial. J. Am. Acad. Dermatol. 2019, 80, 1013–1021. [Google Scholar] [CrossRef]
- Bieber, T.; Thyssen, J.P.; Reich, K.; Simpson, E.L.; Katoh, N.; Torrelo, A.; De Bruin-Weller, M.; Thaci, D.; Bissonnette, R.; Gooderham, M.; et al. Pooled safety analysis of baricitinib in adult patients with atopic dermatitis from 8 randomized clinical trials. J. Eur. Acad. Dermatol. Venereol. 2021, 35, 476–485. [Google Scholar] [CrossRef]
- Pavel, A.B.; Song, T.; Kim, H.-J.; Del Duca, E.; Krueger, J.G.; Dubin, C.; Peng, X.; Xu, H.; Zhang, N.; Estrada, Y.D.; et al. Oral Janus kinase/SYK inhibition (ASN002) suppresses inflammation and improves epidermal barrier markers in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2019, 144, 1011–1024. [Google Scholar] [CrossRef]
- Samrao, A.; Berry, T.M.; Goreshi, R.; Simpson, E.L. A Pilot Study of an Oral Phosphodiesterase Inhibitor (Apremilast) for Atopic Dermatitis in Adults. Arch. Dermatol. 2012, 148, 890–897. [Google Scholar] [CrossRef]
- Dalgard, F.; Lien, L.; Dalen, I. Itch in the community: Associations with psychosocial factors among adults. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 1215–1219. [Google Scholar] [CrossRef]
- Prignano, F.; Ricceri, F.; Pescitelli, L.; Lotti, T. Itch in psoriasis: Epidemiology, clinical aspects and treatment options. Clin. Cosmet. Investig. Dermatol. 2009, 2, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Mollanazar, N.K.; Smith, P.K.; Yosipovitch, G. Mediators of Chronic Pruritus in Atopic Dermatitis: Getting the Itch Out? Clin. Rev. Allergy Immunol. 2016, 51, 263–292. [Google Scholar] [CrossRef] [PubMed]
- Szepietowski, J.; Reich, A. Pruritus in psoriasis: An update. Eur. J. Pain. 2016, 20, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Buddenkotte, J.; Berger, T.G.; Steinhoff, M. Management of Itch in Atopic Dermatitis. Semin. Cutan. Med. Surgery. Semin. Cutan. Med. Surg. 2011, 30, 71–86. [Google Scholar] [CrossRef]
- Jaworecka, K.; Rzepko, M.; Marek-Józefowicz, L.; Tamer, F.; Stefaniak, A.A.; Szczegielniak, M.; Chojnacka-Purpurowicz, J.; Gulekon, A.; Szepietowski, J.C.; Narbutt, J.; et al. The Impact of Pruritus on the Quality of Life and Sleep Disturbances in Pa-tients Suffering from Different Clinical Variants of Psoriasis. J. Clin. Med. 2022, 11, 5553. [Google Scholar] [CrossRef]
- Kaaz, K.; Szepietowski, J.; Matusiak, Ł. Influence of Itch and Pain on Sleep Quality in Atopic Dermatitis and Psoriasis. Acta Derm.-Venereol. 2019, 99, 175–180. [Google Scholar] [CrossRef]
- Kahremany, S.; Hofmann, L.; Gruzman, A.; Cohen, G. Advances in Understanding the Initial Steps of Pruritoceptive Itch: How the Itch Hits the Switch. Int. J. Mol. Sci. 2020, 21, 4883. [Google Scholar] [CrossRef]
- Huet, F.; Taieb, C.; Corgibet, F.; Brenaut, E.; Richard, M.A.; Misery, L. Pruritus, Pain, and Depression Associated with the Most Common Skin Diseases: Data from the French Study “Objectifs Peau”. Dermatology 2022, 238, 448–453. [Google Scholar] [CrossRef]
- Kwatra, S.G.; Kambala, A.; Dong, X. Neuropathic pruritus. J. Allergy Clin. Immunol. 2023, 152, 36–38. [Google Scholar] [CrossRef]
- Brandt, E.B.; Sivaprasad, U. Th2 Cytokines and Atopic Dermatitis. J. Clin. Cell. Immunol. 2011, 2, 1000110. [Google Scholar] [CrossRef]
- Gittler, J.K.; Shemer, A.; Suárez-Fariñas, M.; Fuentes-Duculan, J.; Gulewicz, K.J.; Wang, C.Q.; Mitsui, H.; Cardinale, I.; de Guzman Strong, C.; Krueger, J.G.; et al. Progressive activation of T(H)2/T(H)22 cytokines and selective epidermal proteins characterizes acute and chronic atopic dermatitis. J. Allergy Clin. Immunol. 2012, 130, 1344–1354. [Google Scholar] [CrossRef] [PubMed]
- Pasparakis, M.; Haase, I.; Nestle, F.O. Mechanisms regulating skin immunity and inflammation. Nat. Rev. Immunol. 2014, 14, 289–301. [Google Scholar] [CrossRef]
- Green, D.; Dong, X. The cell biology of acute itch. J. Cell Biol. 2016, 213, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, C.; Ishida, Y.; Kitoh, A.; Otsuka, A.; Kabashima, K. Interaction of peripheral nerves and mast cells, eosinophils, and basophils in the development of pruritus. Exp. Dermatol. 2019, 28, 1405–1411. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Dong, X. Peripheral and Central Mechanisms of Itch. Neuron 2018, 98, 482–494. [Google Scholar] [CrossRef]
- Greb, J.E.; Goldminz, A.M.; Elder, J.T.; Lebwohl, M.G.; Gladman, D.D.; Wu, J.J.; Mehta, N.N.; Finlay, A.Y.; Gottlieb, A.B. Psoriasis. Nat. Rev. Dis. Primers 2016, 2, 16082. [Google Scholar] [CrossRef]
- Campanati, A.; Marani, A.; Martina, E.; Diotallevi, F.; Radi, G.; Offidani, A. Psoriasis as an Immune-Mediated and Inflammatory Systemic Disease: From Pathophysiology to Novel Therapeutic Approaches. Biomedicines 2021, 9, 1511. [Google Scholar] [CrossRef]
- Elewski, B.; Alexis, A.F.; Lebwohl, M.; Gold, L.S.; Pariser, D.; Del Rosso, J.; Yosipovitch, G. Itch: An under-recognized problem in psoriasis. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 1465–1476. [Google Scholar] [CrossRef]
- Park, S.-M.; Kim, G.-W.; Kim, H.-S.; Ko, H.-C.; Kim, M.-B.; Kim, B.-S. Characteristics of Pruritus according to Morphological Phenotype of Psoriasis and Association with Neuropeptides and Interleukin-31. Ann. Dermatol. 2020, 32, 1–7. [Google Scholar] [CrossRef]
- Damiani, G.; Cazzaniga, S.; Conic, R.R.; Naldi, L.; Psocare Registry Network; Griseta, V.; Miracapillo, A.; Azzini, M.; Mocci, L.; Michelini, M.; et al. Pruritus characteristics in a large Italian cohort of psoriatic patients. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 1316–1324. [Google Scholar] [CrossRef]
- Hawro, T.; Hawro, M.; Zalewska-Janowska, A.; Weller, K.; Metz, M.; Maurer, M. Pruritus and sleep disturbances in patients with psoriasis. Arch. Dermatol. Res. 2020, 312, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, D.P.; Sonthalia, S. Koebner Phenomenon. In StatPearls; StatPearls Publishing LLC: Treasure Island, FL, USA, 2020. [Google Scholar]
- Sbidian, E.; Chaimani, A.; Garcia-Doval, I.; Doney, L.; Dressler, C.; Hua, C.; Hughes, C.; Naldi, L.; Afach, S.; Le Cleach, L.; et al. Systemic pharmacological treatments for chronic plaque psoriasis: A network meta-analysis. Cochrane Database Syst. Rev. 2020, 1, CD011535. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Khan, S.; Zaki, A.; Tabassum, G.; Mohsin, M.; Bhutto, H.N.; Ahmad, T.; Fatma, T.; Syed, M.A. Integration of chemokine signaling with non-coding RNAs in tumor microenvironment and heterogeneity in different cancers. Semin. Cancer Biol. 2022, 86, 720–736. [Google Scholar] [CrossRef]
- Mabuchi, T.; Chang, T.W.; Quinter, S.; Hwang, S.T. Chemokine receptors in the pathogenesis and therapy of psoriasis. J. Dermatol. Sci. 2012, 65, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Purzycka-Bohdan, D.; Nedoszytko, B.; Zabłotna, M.; Gleń, J.; Szczerkowska-Dobosz, A.; Nowicki, R.J. Chemokine Profile in Psoriasis Patients in Correlation with Disease Severity and Pruritus. Int. J. Mol. Sci. 2022, 23, 13330. [Google Scholar] [CrossRef]
- Kimura, T.; Sugaya, M.; Suga, H.; Morimura, S.; Miyamoto, A.; Kai, H.; Kagami, S.; Yanaba, K.; Fujita, H.; Asano, Y.; et al. Variations in serum TARC and I-TAC levels reflect minor changes in disease activity and pruritus in atopic dermatitis. Acta Derm. -Venereol. 2014, 94, 331–332. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, Y.; Kamata, M.; Shimizu, T.; Nagata, M.; Fukaya, S.; Hayashi, K.; Fukuyasu, A.; Tanaka, T.; Ishikawa, T.; Ohnishi, T.; et al. Thymus and activation-regulated chemokine (TARC) in patients with psoriasis: Increased serum TARC levels in patients with generalized pustular psoriasis. J. Dermatol. 2020, 47, 1149–1156. [Google Scholar] [CrossRef]
- Nattkemper, L.A.; Tey, H.L.; Valdes-Rodriguez, R.; Lee, H.; Mollanazar, N.K.; Albornoz, C.; Sanders, K.M.; Yosipovitch, G. The Genetics of Chronic Itch: Gene Expression in the Skin of Patients with Atopic Dermatitis and Psoriasis with Severe Itch. J. Investig. Dermatol. 2018, 138, 1311–1317. [Google Scholar] [CrossRef]
- Kabashima, K.; Irie, H. Interleukin-31 as a Clinical Target for Pruritus Treatment. Front. Med. 2021, 8, 69. [Google Scholar] [CrossRef]
- Taneda, K.; Tominaga, M.; Negi, O.; Tengara, S.; Kamo, A.; Ogawa, H.; Takamori, K. Evaluation of epidermal nerve density and opioid receptor levels in psoriatic itch. Br. J. Dermatol. 2011, 165, 277–284. [Google Scholar] [CrossRef]
- Lowes, M.A.; Suárez-Fariñas, M.; Krueger, J.G. Immunology of psoriasis. Annu. Rev. Immunol. 2014, 32, 227–255. [Google Scholar] [CrossRef]
- Salz, M.; Haeberle, S.; Hoffmann, J.; Enk, A.H.; Hadaschik, E.N. Elevated IL-31 serum levels in bullous pemphigoid patients correlate with eosinophil numbers and are associated with BP180-IgE. J. Dermatol. Sci. 2017, 87, 309–311. [Google Scholar] [CrossRef]
- Ohmatsu, H.; Sugaya, M.; Suga, H.; Morimura, S.; Miyagaki, T.; Kai, H.; Kagami, S.; Fujita, H.; Asano, Y.; Tada, Y.; et al. Serum IL-31 Levels are Increased in Patients with Cutaneous T-Cell Lymphoma. Acta Derm. Venereol. 2012, 92, 282–283. [Google Scholar] [CrossRef]
- Gangemi, S.; Quartuccio, S.; Casciaro, M.; Trapani, G.; Minciullo, P.L.; Imbalzano, E. Interleukin 31 and skin diseases: A sys-tematic review. Allergy Asthma Proc. 2017, 38, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Feld, M.; Garcia, R.; Buddenkotte, J.; Katayama, S.; Lewis, K.; Muirhead, G.; Hevezi, P.; Plesser, K.; Schrumpf, H.; Krjutskov, K.; et al. The Pruritus- and TH2-associated Cytokine IL-31 Promotes Growth of Sensory Nerves. J. Allergy Clin. Immunol. 2016, 138, 500–508.e24. [Google Scholar] [CrossRef] [PubMed]
- Purzycka-Bohdan, D.; Gleń, J.; Zabłotna, M.; Nedoszytko, B.; Szczerkowska-Dobosz, A.; Sokołowska-Wojdyło, M.; Rębała, K.; Nowicki, R. Significance of interleukin-31 (IL-31) gene polymorphisms and IL-31 serum level in psoriasis in correlation with pruritus. Postepy Dermatol. Allergol. 2021, 38, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Jegga, A.G.; Shanmukhappa, K.S.; Edukulla, R.; Khurana Hershey, G.H.; Medvedovic, M.; Dillon, S.R.; Madala, S.K. IL-31-driven skin remodeling involves epidermal cell proliferation and thickening that lead to impaired skin-barrier func-tion. PLoS ONE 2016, 11, e0161877. [Google Scholar] [CrossRef]
- Arai, I.; Tsuji, M.; Takeda, H.; Akiyama, N.; Saito, S. A single dose of interleukin-31 (IL-31) causes continuous itch-associated scratching behaviour in mice. Exp. Dermatol. 2013, 22, 669–671. [Google Scholar] [CrossRef]
- Dillon, S.R.; Sprecher, C.; Hammond, A.; Bilsborough, J.; Rosenfeld-Franklin, M.; Presnell, S.R.; Haugen, H.S.; Maurer, M.; Harder, B.; Johnston, J.; et al. Interleukin 31, a cytokine produced by activated T cells, induces dermatitis in mice. Nat. Immunol. 2004, 5, 752–760. [Google Scholar] [CrossRef]
- Czarnecka-Operacz, M.; Polańska, A.; Klimańska, M.; Teresiak-Mikołajczak, E.; Molińska-Glura, M.; Adamski, Z.; Jenerowicz, D. Itching sensation in psoriatic patients and its relation to body mass index and IL-17 and IL-31 concentrations. Adv. Dermatol. Allergol. 2015, 32, 426–430. [Google Scholar] [CrossRef]
- Nakashima, C.; Otsuka, A.; Kabashima, K. Interleukin-31 and interleukin-31 receptor: New therapeutic targets for atopic dermatitis. Exp. Dermatol. 2018, 27, 327–331. [Google Scholar] [CrossRef]
- Nocchi, L.; Roy, N.; D’attilia, M.; Dhandapani, R.; Maffei, M.; Traista, A.; Castaldi, L.; Perlas, E.; Chadick, C.H.; Heppenstall, P.A. Interleukin-31-mediated photoablation of pruritogenic epidermal neurons reduces itch-associated behaviours in mice. Nat. Biomed. Eng. 2019, 3, 114–125. [Google Scholar] [CrossRef]
- Narbutt, J.; Olejniczak, I.; Sobolewska-Sztychny, D.; Sysa-Jedrzejowska, A.; Słowik-Kwiatkowska, I.; Hawro, T.; Lesiak, A. Narrow band ultraviolet B irradiations cause alteration in interleukin-31 serum level in psoriatic patients. Arch. Dermatol. Res. 2013, 305, 191–195. [Google Scholar] [CrossRef]
- Bodoor, K.; Al-Qarqaz, F.; Al Heis, L.; Alfaqih, M.A.; Oweis, A.O.; Almomani, R.; Obeidat, M.A. IL-33/13 axis and IL-4/31 axis play distinct roles in inflammatory process and itch in psoriasis and atopic dermatitis. Clin. Cosm. Investig. Dermatol. 2020, 13, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Hollox, E.J.; Huffmeier, U.; Zeeuwen, P.L.; Palla, R.; Lascorz, J.; Rodijk-Olthuis, D.; Van De Kerkhof, P.C.; Traupe, H.; De Jongh, G.; Heijer, M.D.; et al. Psoriasis is associated with increased beta-defensin genomic copy number. Nat. Genet. 2008, 40, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Harper, E.G.; Guo, C.; Rizzo, H.; Lillis, J.V.; Kurtz, S.E.; Skorcheva, I.; Purdy, D.; Fitch, E.; Iordanov, M.; Blauvelt, A. Th17 cytokines stimulate CCL20 expression in keratinocytes in vitro and in vivo: Implications for psoriasis pathogenesis. J. Investig. Dermatol. 2009, 129, 2175–2183. [Google Scholar] [CrossRef]
- Kimball, A.; Luger, T.; Gottlieb, A.; Puig, L.; Kaufmann, R.; Burge, R.; Lin, C.; Yosipovitch, G. Long-term Impact of Ixekizumab on Psoriasis Itch Severity: Results from a Phase III Clinical Trial and Long-term Extension. Acta Derm. -Venereol. 2018, 98, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Kremer, A.E.; Feramisco, J.; Reeh, P.W.; Beuers, U.; Elferink, R.P.O. Receptors, cells and circuits involved in pruritus of systemic disorders. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1842, 869–892. [Google Scholar] [CrossRef] [PubMed]
- De Felipe, C.; Herrero, J.F.; O’Brien, J.A.; Palmer, J.A.; Doyle, C.A.; Smith, A.J.H.; Laird, J.M.A.; Belmonte, C.; Cervero, F.; Hunt, S.P. Altered nociception, analgesia and aggression in mice lacking the receptor for substance P. Nature 1998, 392, 394–397. [Google Scholar] [CrossRef]
- Pojawa-Gołąb, M.; Jaworecka, K.; Reich, A. NK-1 Receptor Antagonists and Pruritus: Review of Current Literature. Dermatol. Ther. 2019, 9, 391–405. [Google Scholar] [CrossRef]
- Muñoz, M.; Rosso, M.; Pérez, A.; Coveñas, R.; Rosso, R.; Zamarriego, C.; Piruat, J. The NK1 receptor is involved in the antitumoural action of L-733,060 and in the mitogenic action of substance P on neuroblastoma and glioma cell lines. Neuropeptides 2005, 39, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-Y.; Zhao, Y.-Z.; Peng, C.; Li, F.-Q.; Zhu, Q.-G.; Hu, J.-H. Effect of cetirizine hydrochloride on the expression of substance P receptor and cytokines production in human epidermal keratinocytes and dermal fibroblasts. Yao Xue Xue Bao = Acta Pharm. Sin. 2008, 43, 383–387. [Google Scholar]
- Reich, A.; Szepietowski, J.C. Mediators of pruritus in psoriasis. Mediat. Inflamm. 2007, 2007, 064727. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Toyoda, M.; Morohashi, M. Pruritogenic mediators in psoriasis vulgaris: Comparative evaluation of itch-associated cutaneous factors. Br. J. Dermatol. 2003, 149, 718–730. [Google Scholar] [CrossRef] [PubMed]
- Reich, A.; Orda, A.; Wiśnicka, B.; Szepietowski, J.C. Plasma neuropeptides and perception of pruritus in psoriasis. Acta Derm. -Venereol. 2007, 87, 299–304. [Google Scholar] [CrossRef]
- Kongthong, S.; Phumyen, A.; Meephansan, J. Effect of narrowband ultraviolet B therapy on serum levels of CD26/dipeptidyl-peptidase IV and truncated forms of substance P in psoriasis patients with pruritus. Clin. Cosmet. Investig. Dermatol. 2019, 12, 597–604. [Google Scholar] [CrossRef]
- Amatya, B.; Nordlind, K.; Wahlgren, C.-F. Responses to Intradermal Injections of Substance P in Psoriasis Patients with Pruritus. Ski. Pharmacol. Physiol. 2010, 23, 133–138. [Google Scholar] [CrossRef]
- Kobayashi, H.; Hosono, O.; Mimori, T.; Kawasaki, H.; Dang, N.H.; Tanaka, H.; Morimoto, C. Reduction of serum soluble CD26/dipeptidyl peptidase IV enzyme activity and its correlation with disease activity in systemic lupus erythematosus. J. Rheumatol. 2002, 29, 1858–1866. [Google Scholar]
- Stecca, B.A.; Nardo, B.; Chieco, P.; Mazziotti, A.; Bolondi, L.; Cavallari, A. Aberrant dipeptidyl peptidase IV (DPP IV/CD26) expression in human hepatocellular carcinoma. J. Hepatol. 1997, 27, 337–345. [Google Scholar] [CrossRef]
- Pang, R.; Law, W.L.; Chu, A.C.; Poon, J.T.; Lam, C.S.; Chow, A.K.; Ng, L.; Cheung, L.W.; Lan, X.R.; Lan, H.Y.; et al. A subpopulation of CD26+ cancer stem cells with metastatic capacity in human colorectal cancer. Cell Stem Cell 2010, 6, 603–615. [Google Scholar] [CrossRef]
- Inamoto, T.; Yamochi, T.; Ohnuma, K.; Iwata, S.; Kina, S.; Inamoto, S.; Tachibana, M.; Katsuoka, Y.; Dang, N.H.; Morimoto, C. Anti-CD26 monoclonal antibody-mediated G1-S arrest of human renal clear cell carcinoma Caki-2 is associated with reti-noblastoma substrate dephosphorylation, cyclin-dependent kinase 2 reduction, p27(kip1) enhancement, and disruption of binding to the extracellular matrix. Clin. Cancer. Res. 2006, 12, 3470–3477. [Google Scholar]
- Komiya, E.; Tominaga, M.; Kamata, Y.; Suga, Y.; Takamori, K. Molecular and cellular mechanisms of itch in psoriasis. Int. J. Mol. Sci. 2020, 21, 8406. [Google Scholar] [CrossRef] [PubMed]
- Komiya, E.; Hatano, R.; Otsuka, H.; Itoh, T.; Yamazaki, H.; Yamada, T.; Dang, N.H.; Tominaga, M.; Suga, Y.; Kimura, U.; et al. A possible role for CD26/DPPIV enzyme activity in the regulation of psoriatic pruritus. J. Dermatol. Sci. 2017, 86, 212–221. [Google Scholar] [CrossRef]
- Riol-Blanco, L.; Ordovas-Montanes, J.; Perro, M.; Naval, E.; Thiriot, A.; Alvarez, D.; Paust, S.; Wood, J.N.; Von Andrian, U.H. Nociceptive sensory neurons drive interleukin-23-mediated psoriasiform skin inflammation. Nature 2014, 510, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Escalera, J.; Balakrishna, S.; Fan, L.; Caceres, A.I.; Robinson, E.; Sui, A.; McKay, M.C.; McAlexander, M.A.; Herrick, C.A.; et al. TRPA1 controls inflammation and pruritogen responses in allergic contact dermatitis. FASEB J. 2013, 27, 3549–3563. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, S.M.; Belkadi, A.; Loyd, C.M.; Diaconu, D.; Ward, N.L. Cutaneous denervation of psoriasiform mouse skin im-proves acanthosis and inflammation in a sensory neuropeptide-dependent manner. J. Investig. Dermatol. 2011, 131, 1530–1538. [Google Scholar] [CrossRef]
- Kodji, X.; Arkless, K.L.; Kee, Z.; Cleary, S.J.; Aubdool, A.A.; Evans, E.; Caton, P.; Pitchford, S.C.; Brain, S.D. Sensory nerves mediate spontaneous behaviors in addition to inflammation in a murine model of psoriasis. FASEB J. 2019, 33, 1578–1594. [Google Scholar] [CrossRef]
- Özcan, S.; Gürel, G.; Çakır, M. Gene expression profiles of transient receptor potential (TRP) channels in the peripheral blood mononuclear cells of psoriasis patients. Hum. Exp. Toxicol. 2021, 40, 1234–1240. [Google Scholar] [CrossRef]
- Kim, H.O.; Cho, Y.S.; Park, S.Y.; Kwak, I.S.; Choi, M.G.; Chung, B.Y.; Park, C.W.; Lee, J.Y. Increased activity of TRPV3 in keratinocytes in hypertrophic burn scars with postburn pruritus. Wound Repair. Regen. 2016, 24, 841–850. [Google Scholar] [CrossRef]
- Seo, S.H.; Kim, S.; Kim, S.-E.; Chung, S.; Lee, S.E. Enhanced Thermal Sensitivity of TRPV3 in Keratinocytes Underlies Heat-Induced Pruritogen Release and Pruritus in Atopic Dermatitis. J. Investig. Dermatol. 2020, 140, 2199–2209.e6. [Google Scholar] [CrossRef]
- Fatima, M.; Slade, H.; Horwitz, L.; Shi, A.; Liu, J.; McKinstry, D.; Villani, T.; Xu, H.; Duan, B. Abnormal Somatosensory Behaviors Associated With a Gain-of-Function Mutation in TRPV3 Channels. Front. Mol. Neurosci. 2021, 14, 790435. [Google Scholar] [CrossRef] [PubMed]
- Um, J.Y.; Kim, H.B.; Kim, J.C.; Park, J.S.; Lee, S.Y.; Chung, B.Y.; Park, C.W.; Kim, H.O. TRPV3 and Itch: The Role of TRPV3 in Chronic Pruritus according to Clinical and Experimental Evidence. Int. J. Mol. Sci. 2022, 23, 14962. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Shi, Y.; Wu, H.; Niu, C.; Sun, X.; Wang, K. Inhibition of temperature-sensitive TRPV3 channel by two natural iso-chlorogenic acid isomers for alleviation of dermatitis and chronic pruritus. Acta Pharm. Sin. B 2022, 12, 723–734. [Google Scholar]
- Kang, S.Y.; Jung, H.W.; Nam, J.H.; Kim, W.K.; Kang, J.S.; Kim, Y.H.; Cho, C.W.; Cho, C.W.; Park, Y.K.; Bae, H.S. Effects of the Fruit Extract of Tribulus terrestris on Skin Inflammation in Mice with Oxazolone-Induced Atopic Dermatitis through Regu-lation of Calcium Channels, Orai-1 and TRPV3, and Mast Cell Activation. Evidence-based Complement. Altern. Med. 2017, 2017, 8312946. [Google Scholar]
- Nam, J.H.; Jung, H.W.; Chin, Y.W.; Yang, W.M.; Bae, H.S.; Kim, W.K. Spirodela polyrhiza extract modulates the activation of atopic dermatitis-related ion channels, Orai1 and TRPV3, and inhibits mast cell degranulation. Pharm. Biol. 2017, 55, 1324–1329. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, X.; Qi, H.; Ma, Q.; Zhou, Q.; Wang, W.; Wang, K. Pharmacological Inhibition of the Temperature-Sensitive and Ca2+-Permeable Transient Receptor Potential Vanilloid TRPV3 Channel by Natural Forsythoside B Attenuates Pruritus and Cytotoxicity of Keratinocytes. J. Pharmacol. Exp. Ther. 2019, 368, 21–31. [Google Scholar] [CrossRef]
- Sun, X.; Qi, H.; Wu, H.; Qu, Y.; Wang, K. Anti-pruritic and anti-inflammatory effects of natural verbascoside through selective inhibition of temperature-sensitive Ca2+-permeable TRPV3 channel. J. Dermatol. Sci. 2020, 97, 229–231. [Google Scholar] [CrossRef]
- Wang, Y.; Tan, L.; Jiao, K.; Xue, C.; Tang, Q.; Jiang, S.; Ren, Y.; Chen, H.; El-Aziz, T.M.A.; Abdelazeem, K.N.M.; et al. Scutellarein attenuates atopic dermatitis by selectively inhibiting transient receptor potential vanilloid 3 channels. Br. J. Pharmacol. 2022, 179, 4792–4808. [Google Scholar] [CrossRef]
- Bang, S.; Yoo, S.; Yang, T.; Cho, H.; Hwang, S. 17(R)-resolvin D1 specifically inhibits transient receptor potential ion channel vanilloid 3 leading to peripheral antinociception. Br. J. Pharmacol. 2012, 165, 683–692. [Google Scholar] [CrossRef]
- Han, Y.; Luo, A.; Kamau, P.M.; Takomthong, P.; Hu, J.; Boonyarat, C.; Luo, L.; Lai, R. A plant-derived TRPV3 inhibitor suppresses pain and itch. Br. J. Pharmacol. 2021, 178, 1669–1683. [Google Scholar] [CrossRef]
- Rocco, M.L.; Soligo, M.; Manni, L.; Aloe, L. Nerve Growth Factor: Early Studies and Recent Clinical Trials. Curr. Neuropharmacol. 2018, 16, 1455–1465. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.C.; Yang, J.H.; Chang, S.E.; Choi, J. Pruritus and nerve growth factor in psoriasis. Korean J. Dermatol. 2005, 43, 769–773. [Google Scholar]
- Shang, L.; Zhao, S.; Shi, H.; Xing, X.; Zhang, J.; He, Y. Nerve growth factor mediates activation of transient receptor potential vanilloid 1 in neurogenic pruritus of psoriasis. Int. Immunopharmacol. 2023, 118, 110063. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, J.; Aihara, M.; Kobayashi, Y.; Kambara, T.; Ikezawa, Z. Quantitative analysis of nerve growth factor (NGF) in the atopic dermatitis and psoriasis horny layer and effect of treatment on NGF in atopic dermatitis. J. Dermatol. Sci. 2009, 53, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Stein, C. Opioid receptors. Annu. Rev. Med. 2016, 67, 433–451. [Google Scholar] [CrossRef]
- Bigliardi, P.L.; Tobin, D.J.; Gaveriaux-Ruff, C.; Bigliardi-Qi, M. Opioids and the skin–where do we stand? Exp. Dermatol. 2009, 18, 424–430. [Google Scholar] [CrossRef]
- Kupczyk, P.; Reich, A.; Hołysz, M.; Gajda, M.; Wysokińska, E.; Kobuszewska, A.; Nevozhay, D.; Nowakowska, B.; Strzadała, L.; Jagodziński, P.P.; et al. Opioid Receptors in Psoriatic Skin: Relationship with Itch. Acta Derm. Venereol. 2017, 97, 564–570. [Google Scholar] [CrossRef]
- Bigliardi, P.L.; Stammer, H.; Jost, G.; Rufli, T.; Büchner, S.; Bigliardi-Qi, M. Treatment of Pruritus with Topically Applied Opiate Receptor Anatagonist. J. Am. Acad. Dermatol. 2007, 56, 979–988. [Google Scholar] [CrossRef]
- Wiśnicka, B.; Szepietowski, J.; Reich, A.; Orda, A. Histamine, Substance P and Calcitonin Gene-Related Peptide Plasma Concentration and Pruritus in Patients Suffering from Psoriasis. Dermatol. Psychosom. 2004, 5, 73–78. [Google Scholar] [CrossRef]
- Petersen, L.J.; Hansen, U.; Kristensen, J.K.; Nielsen, H.; Skov, P.S.; Nielsen, H.J. Studies on mast cells and histamine release in psoriasis: The effect of ranitidine. Acta Derm. Venereol. 1998, 78, 190–193. [Google Scholar]
- Domagała, A.; Szepietowski, J.; Reich, A. Antihistamines in the treatment of pruritus in psoriasis. Postep. Dermatol. Alergol. 2017, 34, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Mommert, S.; Gschwandtner, M.; Koether, B.; Gutzmer, R.; Werfel, T. Human Memory Th17 Cells Express a Functional His-tamine H4 Receptor. Am. J. Pathol. 2012, 180, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Rossbach, K.; Wahle, K.; Bruer, G.; Brehm, R.; Langeheine, M.; Rode, K.; Bäumer, W. Histamine 2 receptor agonism and his-tamine 4 receptor antagonism ameliorate inflammation in a model of psoriasis. Acta Derm. Venereol. 2020, 100, adv00342. [Google Scholar] [CrossRef] [PubMed]
- Boothe, W.D.; Tarbox, J.A.; Tarbox, M.B. Atopic Dermatitis: Pathophysiology. Adv. Exp. Med. Biol. 2017, 1027, 21–37. [Google Scholar] [CrossRef]
- Wilson, S.R.; Thé, L.; Batia, L.M.; Beattie, K.; Katibah, G.E.; McClain, S.P.; Pellegrino, M.; Estandian, D.M.; Bautista, D.M. The epithelial cell-derived atopic dermatitis cytokine TSLP activates neurons to induce itch. Cell 2013, 155, 285–295. [Google Scholar] [CrossRef]
- Vocks, E.; Busch, R.; Fröhlich, C.; Borelli, S.; Mayer, H.; Ring, J. Influence of weather and climate on subjective symptom intensity in atopic eczema. Int. J. Biometeorol. 2001, 45, 27–33. [Google Scholar] [CrossRef]
- Murota, H.; Yamaga, K.; Ono, E.; Murayama, N.; Yokozeki, H.; Katayama, I. Why does sweat lead to the development of itch in atopic dermatitis? Exp. Dermatol. 2019, 12, 1416–1421. [Google Scholar] [CrossRef]
- Czarnowicki, T.; Krueger, J.G.; Guttman-Yassky, E. Skin barrier and immune dysregulation in atopic dermatitis: An evolving story with important clinical implications. J. Allergy Clin. Immunol. Pract. 2014, 2, 371–379. [Google Scholar] [CrossRef]
- Oh, M.-H.; Oh, S.Y.; Lu, J.; Lou, H.; Myers, A.C.; Zhu, Z.; Zheng, T. TRPA1-Dependent Pruritus in IL-13–Induced Chronic Atopic Dermatitis. J. Immunol. 2013, 191, 5371–5382. [Google Scholar] [CrossRef]
- Yun, J.-W.; Seo, J.A.; Jeong, Y.S.; Bae, I.-H.; Jang, W.-H.; Lee, J.; Kim, S.-Y.; Shin, S.-S.; Woo, B.-Y.; Lee, K.-W.; et al. TRPV1 antagonist can suppress the atopic dermatitis-like symptoms by accelerating skin barrier recovery. J. Dermatol. Sci. 2011, 62, 8–15. [Google Scholar] [CrossRef]
- Fennell, L.M.; Fleming, J.V. Differential processing of mammalian l-histidine decarboxylase enzymes. Biochem. Biophys. Res. Commun. 2014, 445, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Andoh, T.; Yoshihisa, Y.; Shimizu, T. Histamine released from epidermal keratinocytes plays a role in α-melanocyte–stimulating hormone-induced itching in mice. Am. J. Pathol. 2015, 185, 3003–3010. [Google Scholar] [CrossRef] [PubMed]
- Inami, Y.; Nattkemper, L.; Sakai, K.; Yosipovitch, G.; Akiyama, T. Expression of histidine decarboxylase in the epidermis of primates with chronic itch. Acta Derm. Venereol. 2017, 97, 739–740. [Google Scholar] [CrossRef] [PubMed]
- Gutowska-Owsiak, D.; Greenwald, L.; Watson, C.; Selvakumar, T.; Wang, X.; Ogg, G. The histamine-synthesizing enzyme histidine decarboxylase is upregulated by keratinocytes in atopic skin. Br. J. Dermatol. 2014, 171, 771–778. [Google Scholar] [CrossRef]
- Li, W.; Yosipovitch, G. The Role of the Microbiome and Microbiome-Derived Metabolites in Atopic Dermatitis and Non-Histaminergic Itch. Am. J. Clin. Dermatol. 2020, 21, 44–50. [Google Scholar] [CrossRef]
- Kong, H.H.; Oh, J.; Deming, C.; Conlan, S.; Grice, E.A.; Beatson, M.A.; Nomicos, E.; Polley, E.C.; Komarow, H.D.; NISC Comparative Sequence Program; et al. Temporal shifts in the skin microbiome associated with disease flares and treatment in children with atopic dermatitis. Genome Res. 2012, 22, 850–859. [Google Scholar] [CrossRef]
- Blicharz, L.; Usarek, P.; Młynarczyk, G.; Skowroński, K.; Rudnicka, L.; Samochocki, Z. Is Itch Intensity in Atopic Dermatitis Associated with Skin Colonization by Staphylococcus aureus? Indian J. Dermatol. 2020, 65, 17–21. [Google Scholar]
- Travers, J.B.; Kozman, A.; Mousdicas, N.; Saha, C.; Landis, M.; Al-Hassani, M.; Yao, W.; Yao, Y.; Hyatt, A.-M.; Sheehan, M.P.; et al. Infected atopic dermatitis lesions contain pharmacologic amounts of lipoteichoic acid. J. Allergy Clin. Immunol. 2010, 125, 146–152.e1-2. [Google Scholar] [CrossRef]
- Airaksinen, M.S.; Saarma, M. The GDNF family: Signaling, biological functions and therapeutic value. Nat. Rev. Neurosci. 2002, 3, 383–394. [Google Scholar] [CrossRef]
- Murota, H.; Izumi, M.; Abd El-Latif, M.I.; Nishioka, M.; Terao, M.; Tani, M.; Matsui, S.; Sano, S.; Katayama, I. Artemin causes hypersensitivity to warm sensation, mimicking warmth-provoked pruritus in atopic dermatitis. J. Allergy Clin. Immunol. 2012, 130, 671–682.e4. [Google Scholar] [CrossRef]
- Michelsen, K.S.; Aicher, A.; Mohaupt, M.; Hartung, T.; Dimmeler, S.; Kirschning, C.J.; Schumann, R.R. The role of toll-like receptors (TLRs) in bacteria-induced maturation of murine dendritic cells (DCs): Peptidoglycan and lipoteichoic acid are inducers of DC maturation and require TLR2. J. Biol. Chem. 2001, 276, 25680–25686. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Tang, M.; Ma, X.; Yang, L.; He, J.; Peng, X.; Guo, G.; Zhou, L.; Luo, N.; Yuan, Z.; et al. AP-57/C10orf99 is a new type of multifunctional antimicrobial peptide. Biochem. Biophys. Res. Commun. 2015, 457, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Green, D.P.; Limjunyawong, N.; Gour, N.; Pundir, P.; Dong, X. A Mast-Cell-Specific Receptor Mediates Neurogenic Inflammation and Pain. Neuron 2019, 101, 412–420.e3. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Luo, Y.; Mai, G.; Zhang, M.; Wang, G.; Zhao, M.; Gao, L.; Li, F.; Zhou, F. Gene expression profile based classification models of psoriasis. Genomics 2014, 103, 48–55. [Google Scholar] [CrossRef]
- Tseng, P.-Y.; Hoon, M.A. GPR15L is an epithelial inflammation-derived pruritogen. Sci. Adv. 2022, 8, eabm7342. [Google Scholar] [CrossRef]
- Nygaard, U.; Hvid, M.; Johansen, C.; Buchner, M.; Fölster-Holst, R.; Deleuran, M.; Vestergaard, C. TSLP, IL-31, IL-33 and sST2 are new biomarkers in endophenotypic profiling of adult and childhood atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 1930–1938. [Google Scholar] [CrossRef]
- Guttman-Yassky, E.; Teixeira, H.D.; Simpson, E.L.; Papp, K.A.; Pangan, A.L.; Blauvelt, A.; Thaçi, D.; Chu, C.-Y.; Hong, H.C.-H.; Katoh, N.; et al. Once-daily upadacitinib versus placebo in adolescents and adults with moderate-to-severe atopic dermatitis (Measure Up 1 and Measure Up 2): Results from two replicate double-blind, randomised controlled phase 3 trials. Lancet 2021, 397, 2151–2168. [Google Scholar] [CrossRef]
- Napolitano, M.; Fabbrocini, G.; Genco, L.; Martora, F.; Potestio, L.; Patruno, C. Rapid improvement in pruritus in atopic dermatitis patients treated with upadacitinib: A real-life experience. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 1497–1498. [Google Scholar] [CrossRef]
- Xiao, S.; Lu, Z.; Steinhoff, M.; Li, Y.; Buhl, T.; Fischer, M.; Chen, W.; Cheng, W.; Zhu, R.; Yan, X.; et al. Innate immune regulates cutaneous sensory IL-13 receptor alpha 2 to promote atopic dermatitis. Brain Behav. Immun. 2021, 98, 28–39. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaczmarska, A.; Kwiatkowska, D.; Skrzypek, K.K.; Kowalewski, Z.T.; Jaworecka, K.; Reich, A. Pathomechanism of Pruritus in Psoriasis and Atopic Dermatitis: Novel Approaches, Similarities and Differences. Int. J. Mol. Sci. 2023, 24, 14734. https://doi.org/10.3390/ijms241914734
Kaczmarska A, Kwiatkowska D, Skrzypek KK, Kowalewski ZT, Jaworecka K, Reich A. Pathomechanism of Pruritus in Psoriasis and Atopic Dermatitis: Novel Approaches, Similarities and Differences. International Journal of Molecular Sciences. 2023; 24(19):14734. https://doi.org/10.3390/ijms241914734
Chicago/Turabian StyleKaczmarska, Agnieszka, Dominika Kwiatkowska, Katarzyna Konstancja Skrzypek, Zbigniew Tadeusz Kowalewski, Kamila Jaworecka, and Adam Reich. 2023. "Pathomechanism of Pruritus in Psoriasis and Atopic Dermatitis: Novel Approaches, Similarities and Differences" International Journal of Molecular Sciences 24, no. 19: 14734. https://doi.org/10.3390/ijms241914734