
Immune Activation and Inflammatory Response Mediated by the NOD/Toll-like Receptor Signaling Pathway—The Potential Mechanism of Bullfrog (Lithobates catesbeiana) Meningitis Caused by Elizabethkingia miricola
 ,
,
Abstract
:1. Introduction
2. Results
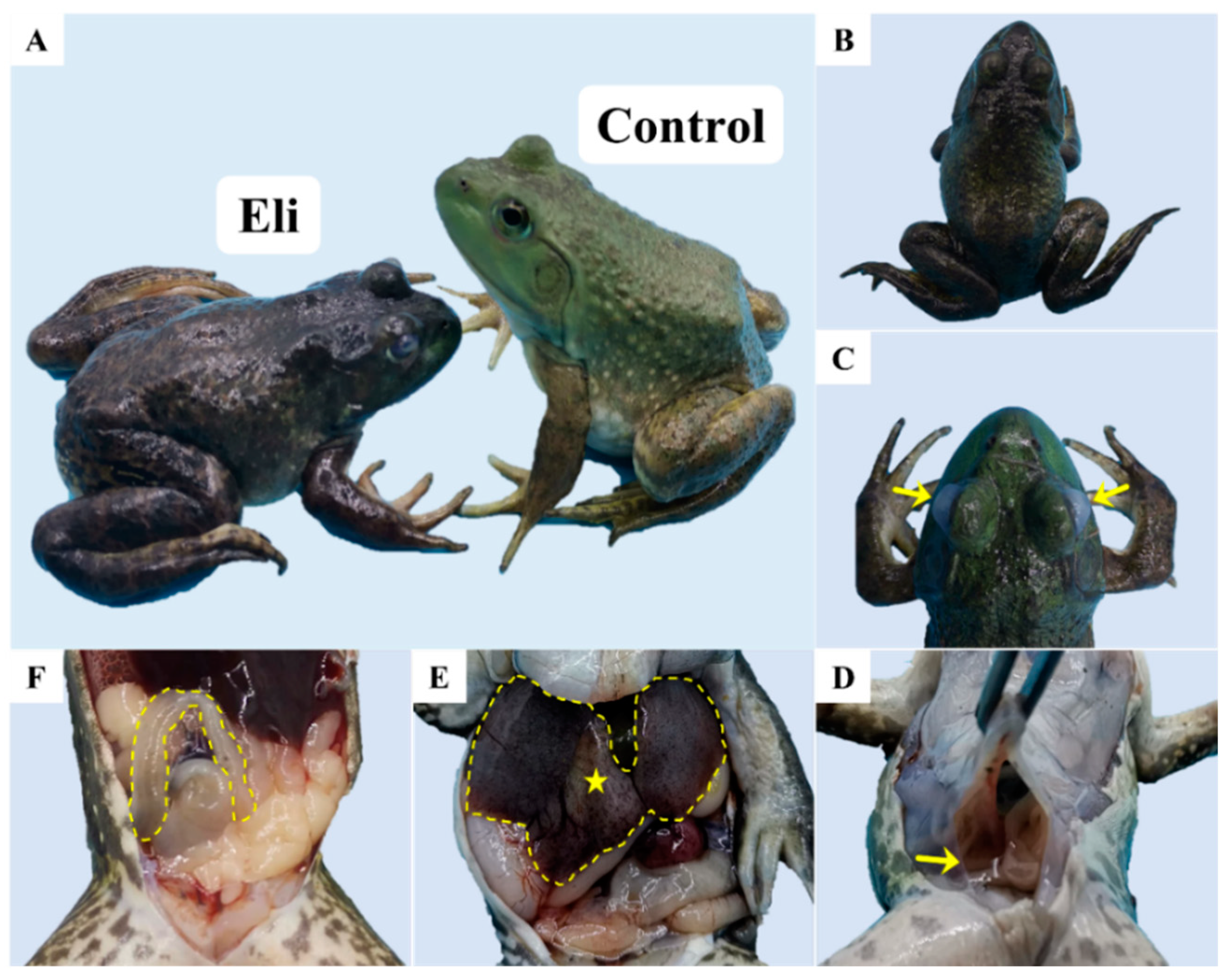
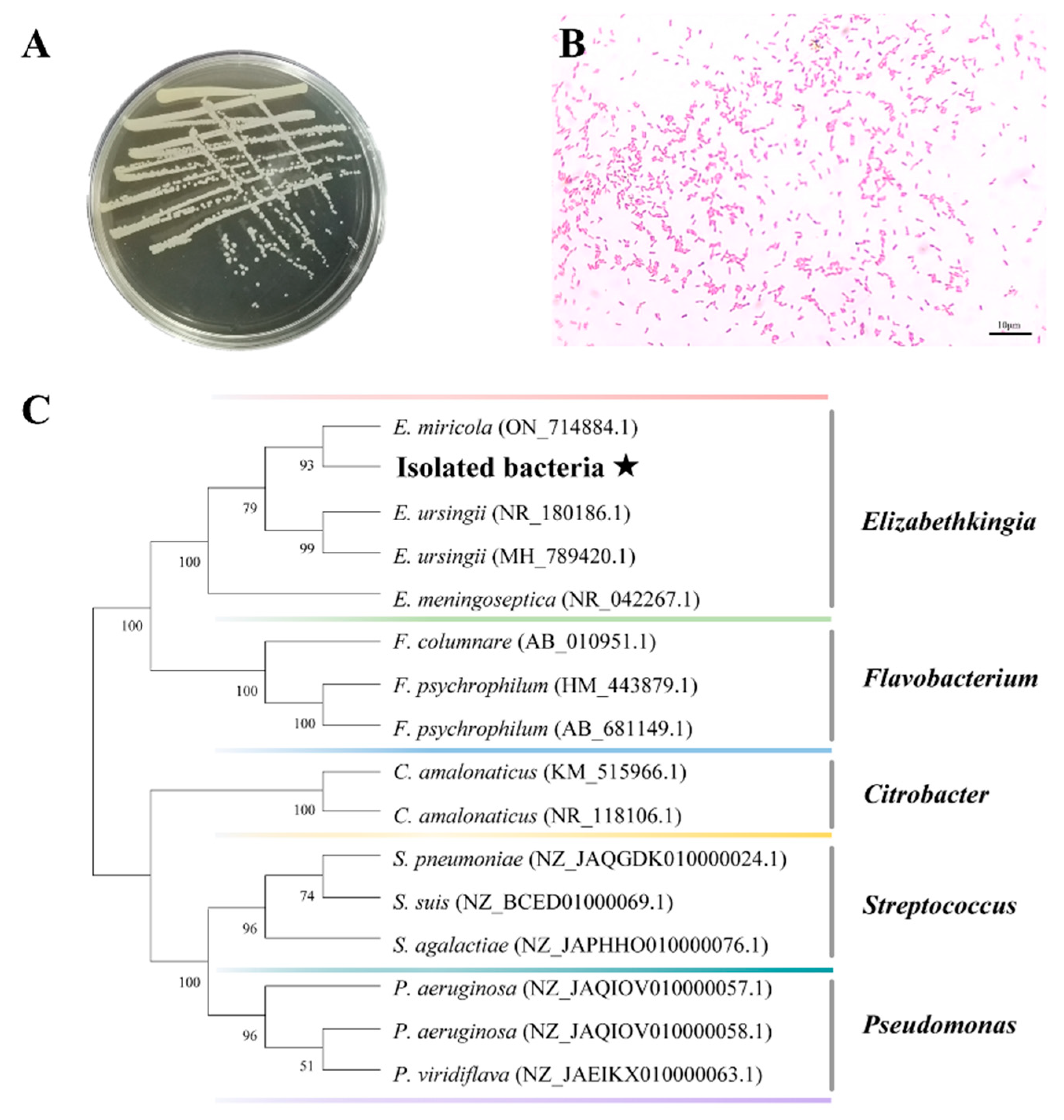
2.1. Isolation and Identification of Pathogenic Bacteria
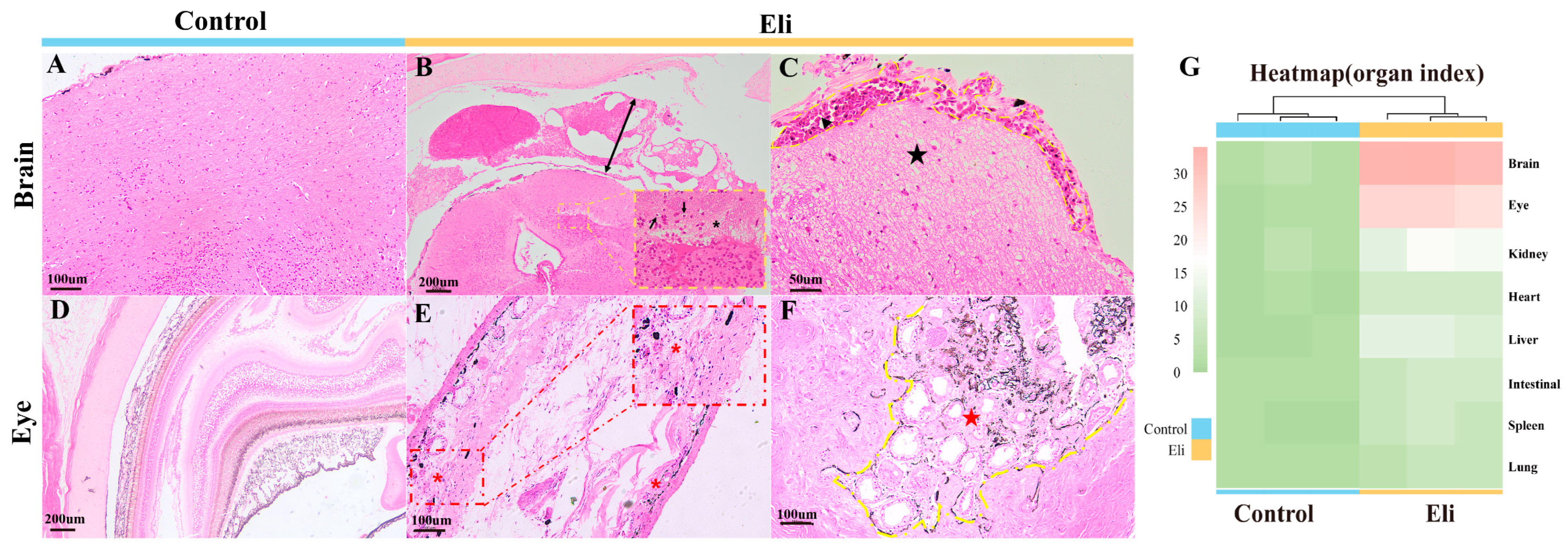
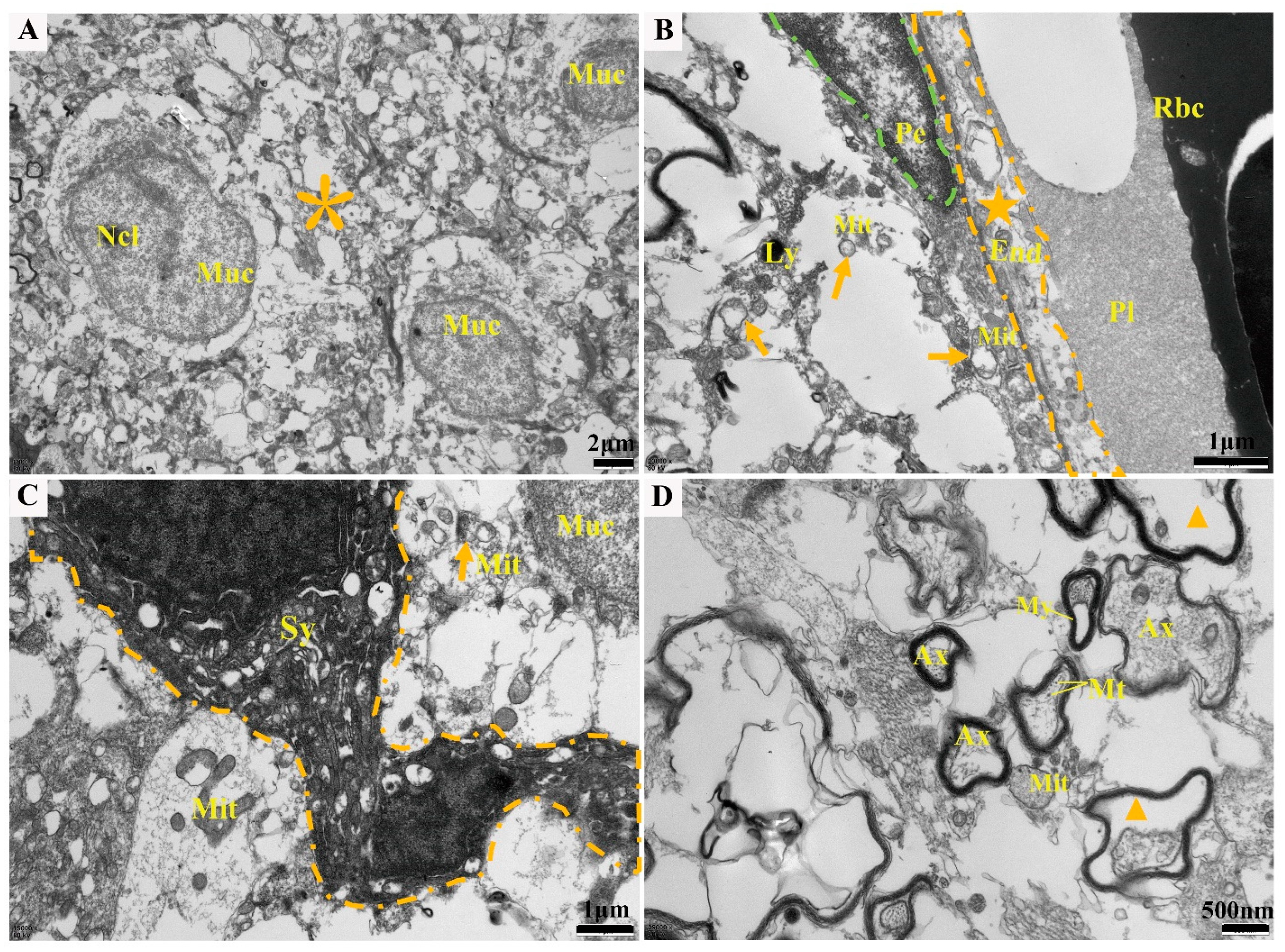
2.2. Histopathological Changes
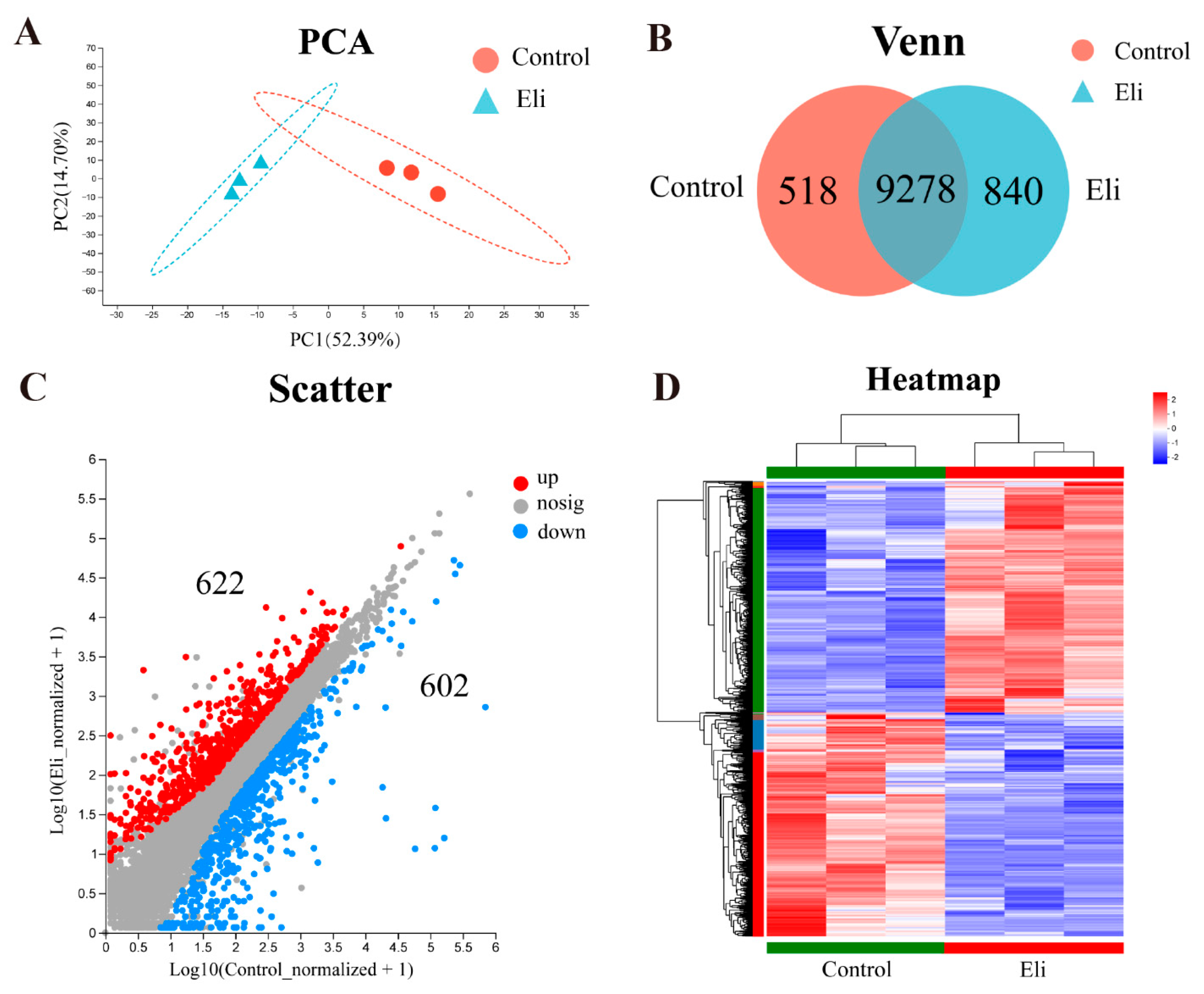
2.3. Elizabethkingia Miricola Infection Affects Gene Expression Profile in the Brain of Bullfrog
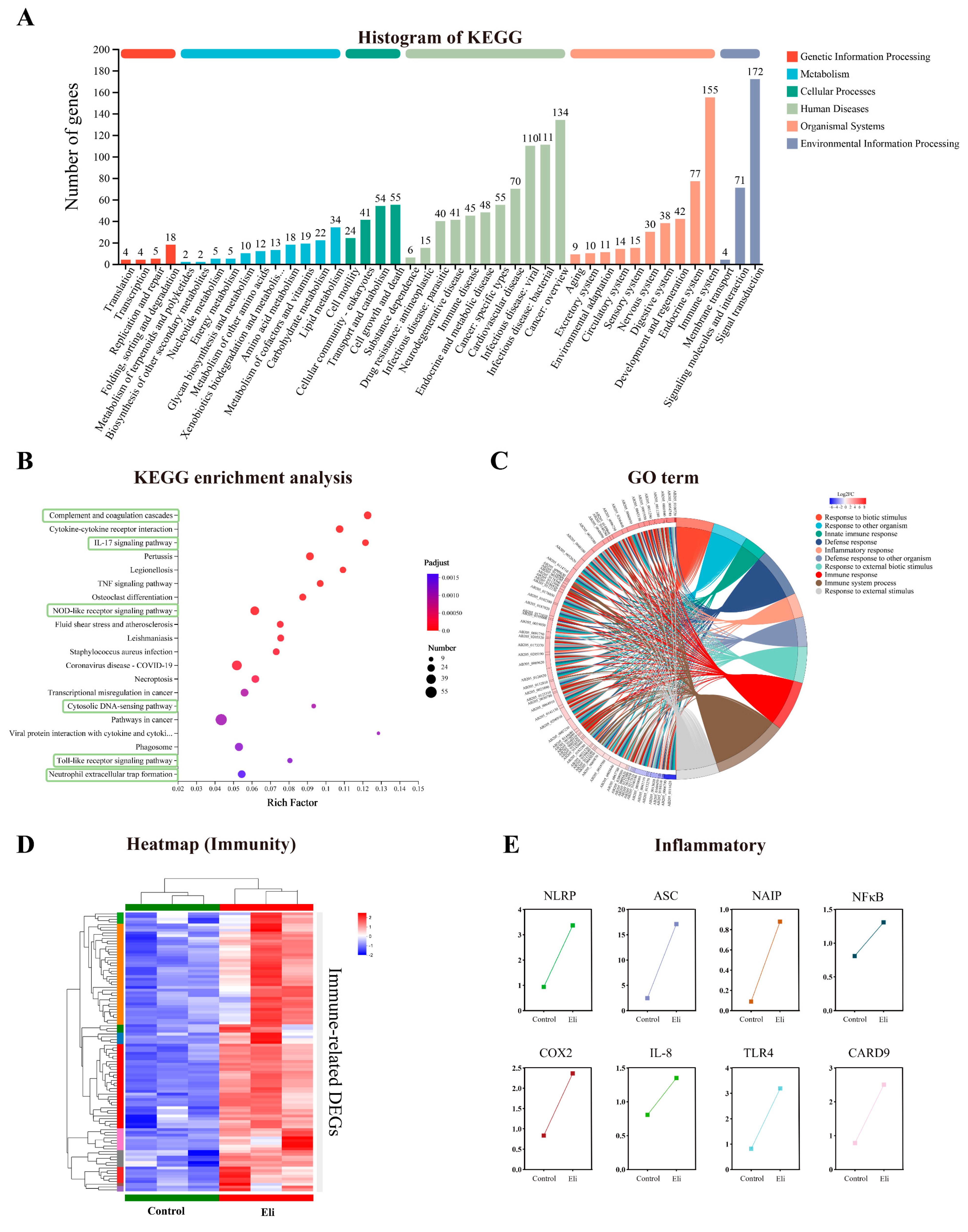
2.4. Immune Pathways Are Enriched in Brains after Infection with Elizabethkingia Miricola
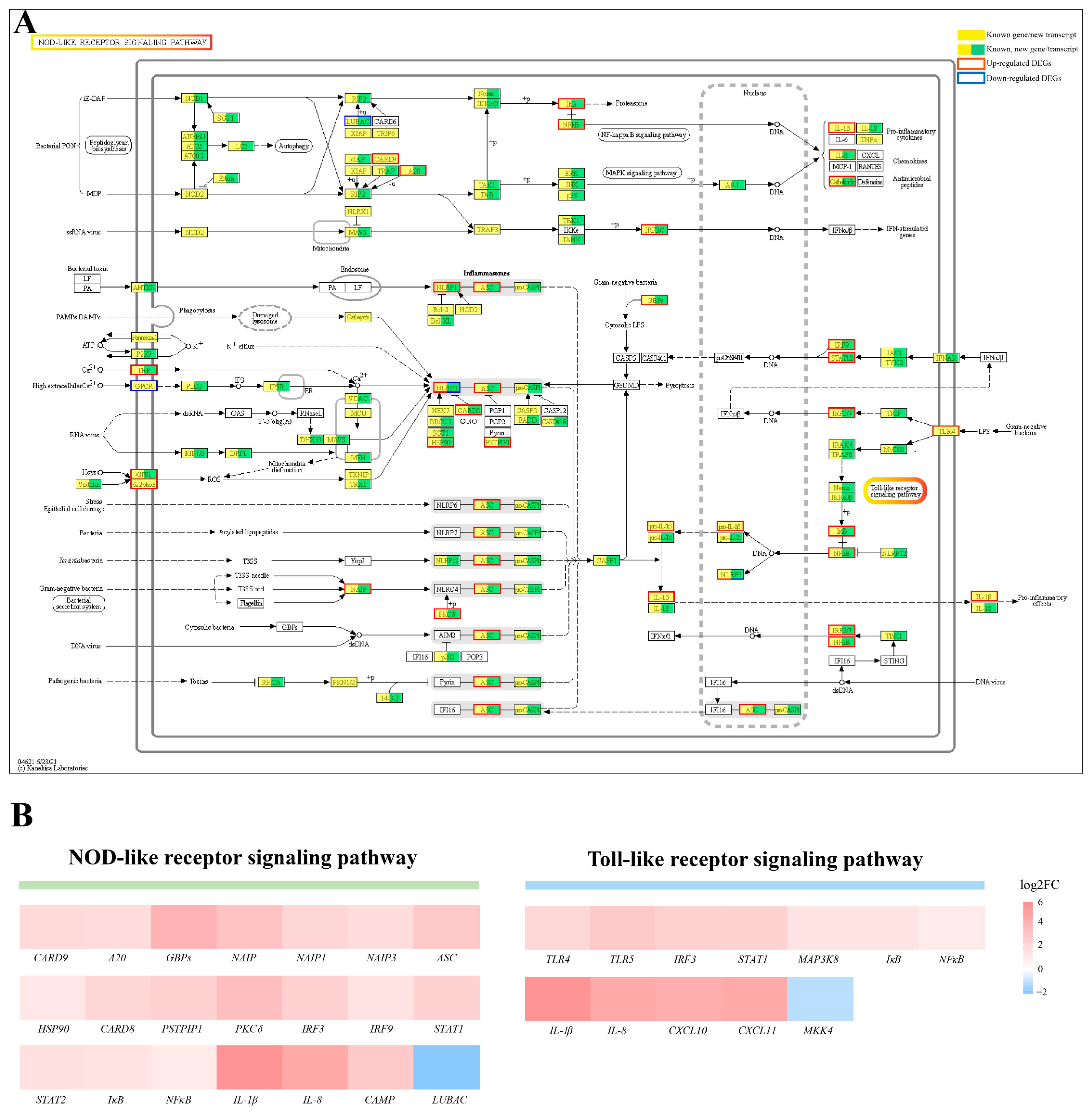
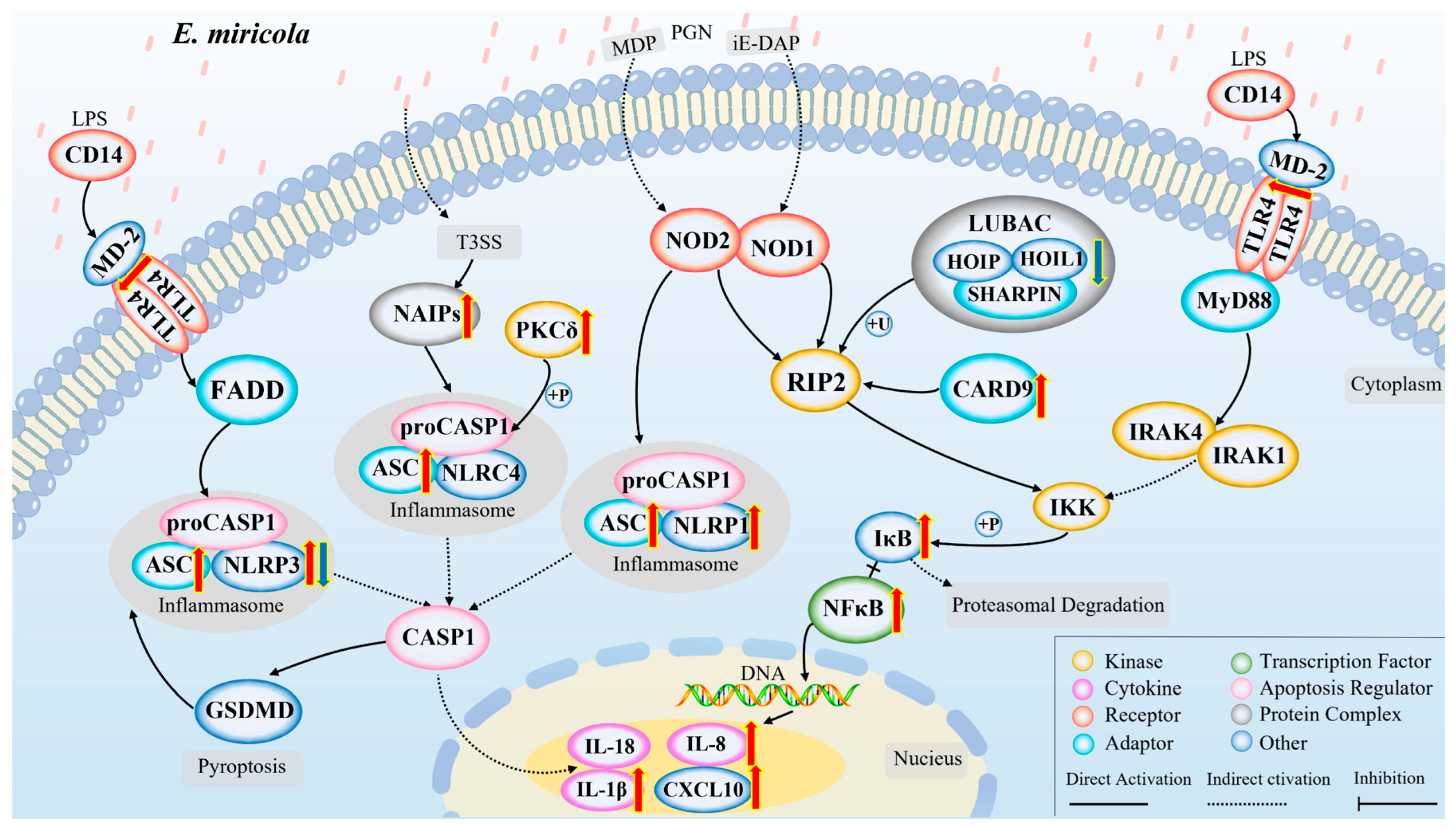
2.5. Activation of NOD/Toll-like Signaling Pathway after Infection with Elizabethkingia Miricola
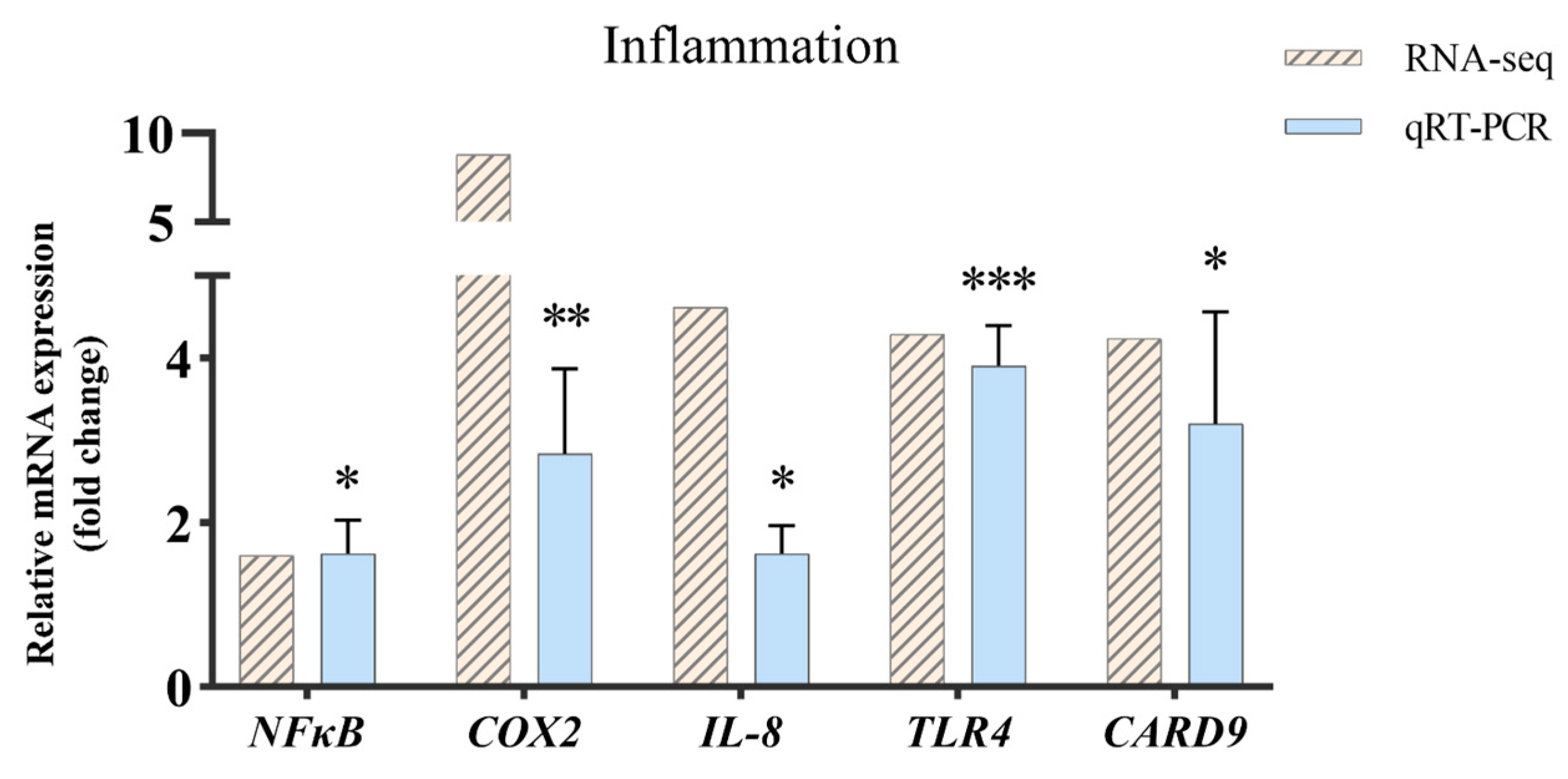
2.6. Validation of RNA-Seq Results via Quantitative PCR Analysis
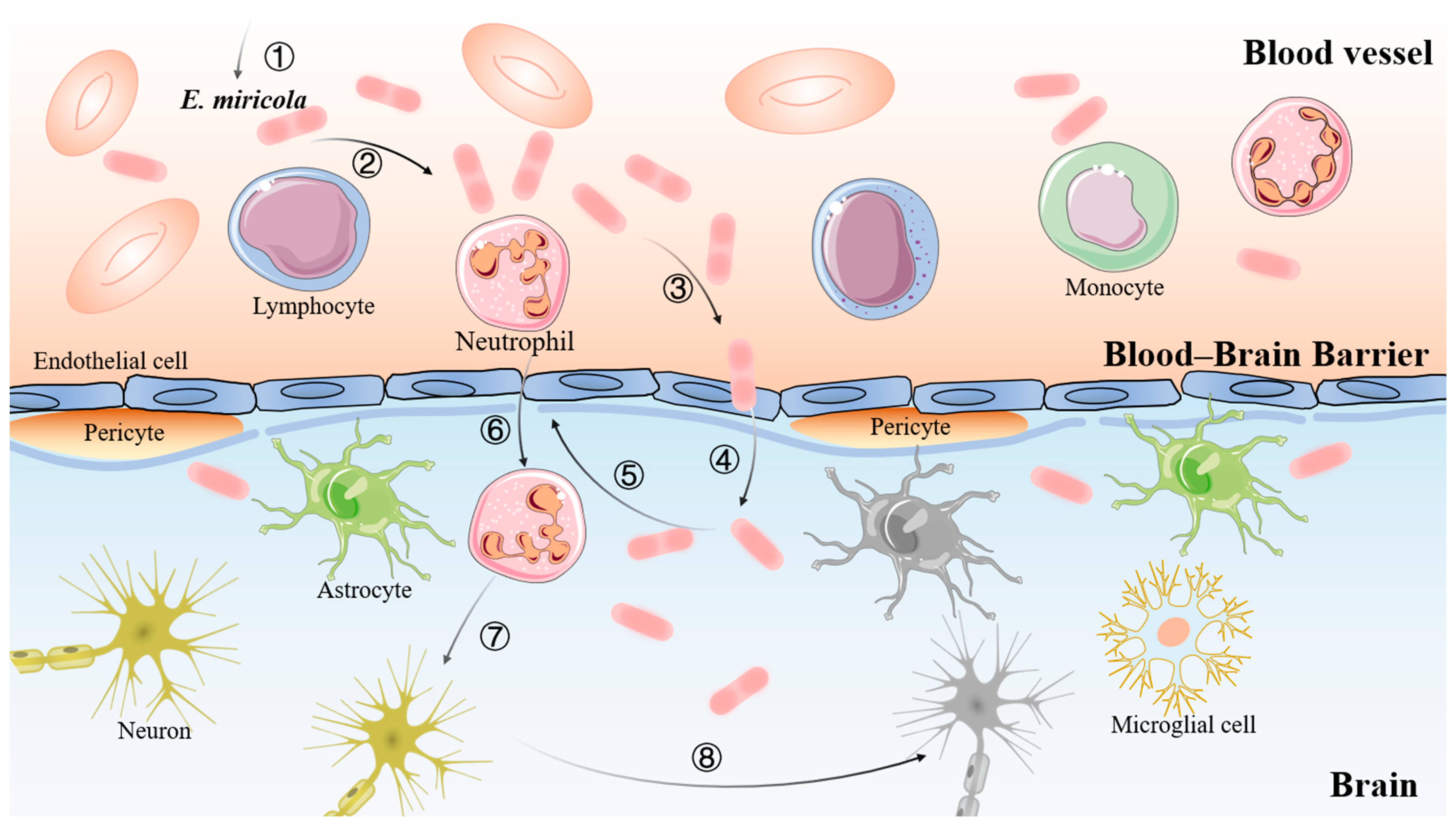
3. Discussion
4. Materials and Methods
4.1. Bullfrogs Sample Collection
4.2. Isolation and Sequencing of Pathogenic Bacteria
4.3. Histological Analysis of Bullfrogs
4.4. Transmission Electron Microscopy
4.5. RNA Isolation, cDNA Library Preparation, and Sequencing
4.6. Transcriptome Quality Control and Gene Annotation
4.7. Differential Gene Expression and Enrichment Analysis
4.8. qRT-PCR Validation of DEG Expression
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.; Kawamura, Y.; Fujiwara, N.; Naka, T.; Liu, H.; Huang, X.; Kobayashi, K.; Ezaki, T. Chryseobacterium miricola sp. nov., A Novel Species Isolated from Condensation Water of Space Station Mir. Syst. Appl. Microbiol. 2003, 26, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Kim, M.K.; Lim, J.H.; Park, H.Y.; Lee, S.-T. Transfer of Chryseobacterium meningosepticum and Chryseobacterium miricola to Elizabethkingia gen. nov. as Elizabethkingia meningoseptica comb. nov. and Elizabethkingia miricola comb. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Yuan, J.; Meng, Y.; Wang, Z.; Gu, Z. Pathogenic Elizabethkingia miricola infection in cultured black-spotted frogs, China 2016. Emerg. Infect. Dis. 2017, 23, 2055. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Li, T.; Feng, L.; Zhang, S. Elizabethkingia miricola Causes Intracranial Infection: A Case Study. Front. Med. 2021, 8, 761924. [Google Scholar] [CrossRef] [PubMed]
- Opota, O.; Diene, S.M.; Bertelli, C.; Prod’Hom, G.; Eckert, P.; Greub, G. Genome of the carbapenemase-producing clinical isolate Elizabethkingia miricola EM_CHUV and comparative genomics with Elizabethkingia meningoseptica and Elizabethkingia anophelis: Evidence for intrinsic multidrug resistance trait of emerging pathogens. Int. J. Antimicrob. Agents 2017, 49, 93–97. [Google Scholar] [CrossRef]
- Green, O.; Murray, P.; Gea-Banacloche, J.C. Sepsis caused by Elizabethkingia miricola successfully treated with tigecycline and levofloxacin. Diagn. Microbiol. Infect. Dis. 2008, 62, 430–432. [Google Scholar] [CrossRef]
- Gupta, P.; Zaman, K.; Mohan, B.; Taneja, N. Elizabethkingia miricola: A rare non-fermenter causing urinary tract infection. World J. Clin. Cases 2017, 5, 187–190. [Google Scholar] [CrossRef]
- Huang, X.; Feng, Y.; Tang, H.; Xiong, G.; Li, L.; Yang, Y.; Wang, K.; Ouyang, P.; Geng, Y.; Chen, D. Candidate animal disease model of Elizabethkingia spp. infection in humans, based on the systematic pathology and oxidative damage caused by E. miricola in pelophylax nigromaculatus. Oxidative Med. Cell. Longev. 2019, 2019, 6407524. [Google Scholar] [CrossRef]
- Rodrigues, C.M.; Maiden, M.C. A world without bacterial meningitis: How genomic epidemiology can inform vaccination strategy. F1000Research 2018, 7, 401. [Google Scholar] [CrossRef]
- Yang, R.; Wang, J.; Wang, F.; Zhang, H.; Tan, C.; Chen, H.; Wang, X. Blood–Brain Barrier Integrity Damage in Bacterial Meningitis: The Underlying Link, Mechanisms, and Therapeutic Targets. Int. J. Mol. Sci. 2023, 24, 2852. [Google Scholar] [CrossRef]
- de Vries, H.E.; Blom-Roosemalen, M.C.; van Oosten, M.; de Boer, A.G.; van Berkel, T.J.; Breimer, D.D.; Kuiper, J. The influence of cytokines on the integrity of the blood-brain barrier in vitro. J. Neuroimmunol. 1996, 64, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Prager, O.; Friedman, A.; Nebenzahl, Y.M. Role of neural barriers in the pathogenesis and outcome of Streptococcus pneumoniae meningitis. Exp. Ther. Med. 2017, 13, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Peng, L.; Gai, Z.; Zhang, L.; Jong, A.; Cao, H.; Huang, S.-H. Pathogenic Triad in Bacterial Meningitis: Pathogen Invasion, NF-κB Activation, and Leukocyte Transmigration that Occur at the Blood-Brain Barrier. Front. Microbiol. 2016, 7, 148. [Google Scholar] [CrossRef]
- Zhang, C.-X.; Huang, K.-K.; Wang, L.; Song, K.; Zhang, L.; Li, P. Apparent digestibility coefficients and amino acid availability of common protein ingredients in the diets of bullfrog, Rana (Lithobates) catesbeiana. Aquaculture 2014, 437, 38–45. [Google Scholar] [CrossRef]
- Lin, H.; Ma, J.; Sun, J.; Qin, Z.; Jiang, B.; Li, W.; Wang, Q.; Su, Y.; Lin, L.; Liu, C. Identification and Characterization of Klebsiella pneumoniae from Farmed American Bullfrogs (Rana catesbeiana). Microbiol. Spectr. 2023, 11, e0357922. [Google Scholar] [CrossRef]
- Pasteris, S.; Bühler, M.; Nader-Macías, M. Microbiological and histological studies of farmed-bullfrog (Rana catesbeiana) tissues displaying red-leg syndrome. Aquaculture 2006, 251, 11–18. [Google Scholar] [CrossRef]
- Wei, D.; Cheng, Y.; Liao, W.; Yu, Q.; Shi, J.; Xia, X.; Chen, M.; Han, S.; Li, P. Composition and function of the skin microbiota were altered of red leg syndrome in cultured bullfrog (Rana catesbeiana). Aquac. Rep. 2023, 29, 101487. [Google Scholar] [CrossRef]
- Wei, D.; Xiao, S.; Liao, W.; Yu, Q.; Han, S.; Shi, J.; He, J.; Li, P. The occurrence of Morganella morganii caused large death in cultured American bullfrog (Rana catebeiana). Aquaculture 2023, 568, 739343. [Google Scholar] [CrossRef]
- Lin, J.-N.; Lai, C.-H.; Yang, C.-H.; Huang, Y.-H. Elizabethkingia Infections in Humans: From Genomics to Clinics. Microorganisms 2019, 7, 295. [Google Scholar] [CrossRef]
- Lei, X.P.; Yi, G.; Wang, K.Y.; OuYang, P.; Chen, D.F.; Huang, X.L.; Huang, C.; Lai, W.M.; Zhong, Z.J.; Huo, C.L. Elizabethkingia miricola infection in Chinese spiny frog (Quasipaa spinosa). Transbound. Emerg. Dis. 2019, 66, 1049–1053. [Google Scholar] [CrossRef]
- Trimpert, J.; Eichhorn, I.; Vladimirova, D.; Haake, A.; Schink, A.; Klopfleisch, R.; Lübke-Becker, A. Elizabethkingia miricola infection in multiple anuran species. Transbound. Emerg. Dis. 2020, 68, 931–940. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Zhao, T.; Zhao, C.; Zhu, W.; Xu, L.; Liu, J.; Li, C.; Xie, F.; Jiang, J.; Zhu, W. Microbiomic and transcriptomic insight into the pathogenesis of meningitis-like disease in cultured Pelophylax nigromaculatus. Aquaculture 2020, 530, 735736. [Google Scholar] [CrossRef]
- Sun, B.; Li, X.; Ning, X.; Sun, L. Transcriptome Analysis of Paralichthys olivaceus Erythrocytes Reveals Profound Immune Responses Induced by Edwardsiella tarda Infection. Int. J. Mol. Sci. 2020, 21, 3094. [Google Scholar] [CrossRef]
- Meylan, E.; Tschopp, J.; Karin, M. Intracellular pattern recognition receptors in the host response. Nature 2006, 442, 39–44. [Google Scholar] [CrossRef]
- Philpott, D.J.; Girardin, E.S. Nod-like receptors: Sentinels at host membranes. Curr. Opin. Immunol. 2010, 22, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Rubino, S.J.; Selvanantham, T.; Girardin, E.S.; Philpott, D.J. Nod-like receptors in the control of intestinal inflammation. Curr. Opin. Immunol. 2012, 24, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2009, 21, 317–337. [Google Scholar] [CrossRef]
- Viala, J.; Chaput, C.; Boneca, I.G.; Cardona, A.; Girardin, S.E.; Moran, A.P.; Athman, R.; Mémet, S.; Huerre, M.R.; Coyle, A.J.; et al. Nod1 responds to peptidoglycan delivered by the Helicobacter pylori cag pathogenicity island. Nat. Immunol. 2004, 5, 1166–1174. [Google Scholar] [CrossRef]
- Kobayashi, K.S.; Chamaillard, M.; Ogura, Y.; Henegariu, O.; Inohara, N.; Nuñez, G.; Flavell, R.A. Nod2-Dependent Regulation of Innate and Adaptive Immunity in the Intestinal Tract. Science 2005, 307, 731–734. [Google Scholar] [CrossRef] [PubMed]
- Miyake, K. Innate immune sensing of pathogens and danger signals by cell surface Toll-like receptors. Semin. Immunol. 2007, 19, 3–10. [Google Scholar] [CrossRef]
- Kim, K.S. Pathogenesis of bacterial meningitis: From bacteraemia to neuronal injury. Nat. Rev. Neurosci. 2003, 4, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Le Guennec, L.; Coureuil, M.; Nassif, X.; Bourdoulous, S. Strategies used by bacterial pathogens to cross the blood–brain barrier. Cell. Microbiol. 2019, 22, e13132. [Google Scholar] [CrossRef] [PubMed]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in fish: Proposal for a protocol to assess aquatic pollution. J. Fish Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32 (Suppl. S1), D277–D280. [Google Scholar] [CrossRef]
- Rigden, D.J.; Fernández, X.M. The 2022 Nucleic Acids Research database issue and the online molecular biology database collection. Nucleic Acids Res. 2022, 50, D1–D10. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39 (Suppl. S2), W316–W322. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Raw Reads | Raw Bases | Clean Reads | Error Rate (%) | Q20 (%) |
|---|---|---|---|---|---|
| Control-1 | 55,240,800 | 8,341,360,800 | 54,645,342 | 0.0259 | 97.71 |
| Control-2 | 55,210,162 | 8,336,734,462 | 54,541,358 | 0.0265 | 97.51 |
| Control-3 | 63,303,662 | 9,558,852,962 | 62,634,710 | 0.0262 | 97.64 |
| Eli-1 | 66,743,592 | 10,078,282,392 | 65,771,358 | 0.0274 | 97.14 |
| Eli-2 | 60,743,954 | 9,172,337,054 | 59,997,100 | 0.0268 | 97.36 |
| Eli-3 | 62,318,058 | 9,410,026,758 | 61,569,246 | 0.027 | 97.29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Chen, B.; Xu, M.; Feng, Y.; Deng, Y.; Huang, X.; Geng, Y.; Ouyang, P.; Chen, D. Immune Activation and Inflammatory Response Mediated by the NOD/Toll-like Receptor Signaling Pathway—The Potential Mechanism of Bullfrog (Lithobates catesbeiana) Meningitis Caused by Elizabethkingia miricola. Int. J. Mol. Sci. 2023, 24, 14554. https://doi.org/10.3390/ijms241914554
Li F, Chen B, Xu M, Feng Y, Deng Y, Huang X, Geng Y, Ouyang P, Chen D. Immune Activation and Inflammatory Response Mediated by the NOD/Toll-like Receptor Signaling Pathway—The Potential Mechanism of Bullfrog (Lithobates catesbeiana) Meningitis Caused by Elizabethkingia miricola. International Journal of Molecular Sciences. 2023; 24(19):14554. https://doi.org/10.3390/ijms241914554
Chicago/Turabian StyleLi, Fulong, Baipeng Chen, Ming Xu, Yang Feng, Yongqiang Deng, Xiaoli Huang, Yi Geng, Ping Ouyang, and Defang Chen. 2023. "Immune Activation and Inflammatory Response Mediated by the NOD/Toll-like Receptor Signaling Pathway—The Potential Mechanism of Bullfrog (Lithobates catesbeiana) Meningitis Caused by Elizabethkingia miricola" International Journal of Molecular Sciences 24, no. 19: 14554. https://doi.org/10.3390/ijms241914554