
Oryza sativa L. Indica Seed Coat Ameliorated Concanavalin A—Induced Acute Hepatitis in Mice via MDM2/p53 and PKCα/MAPK1 Signaling Pathways
 and
and 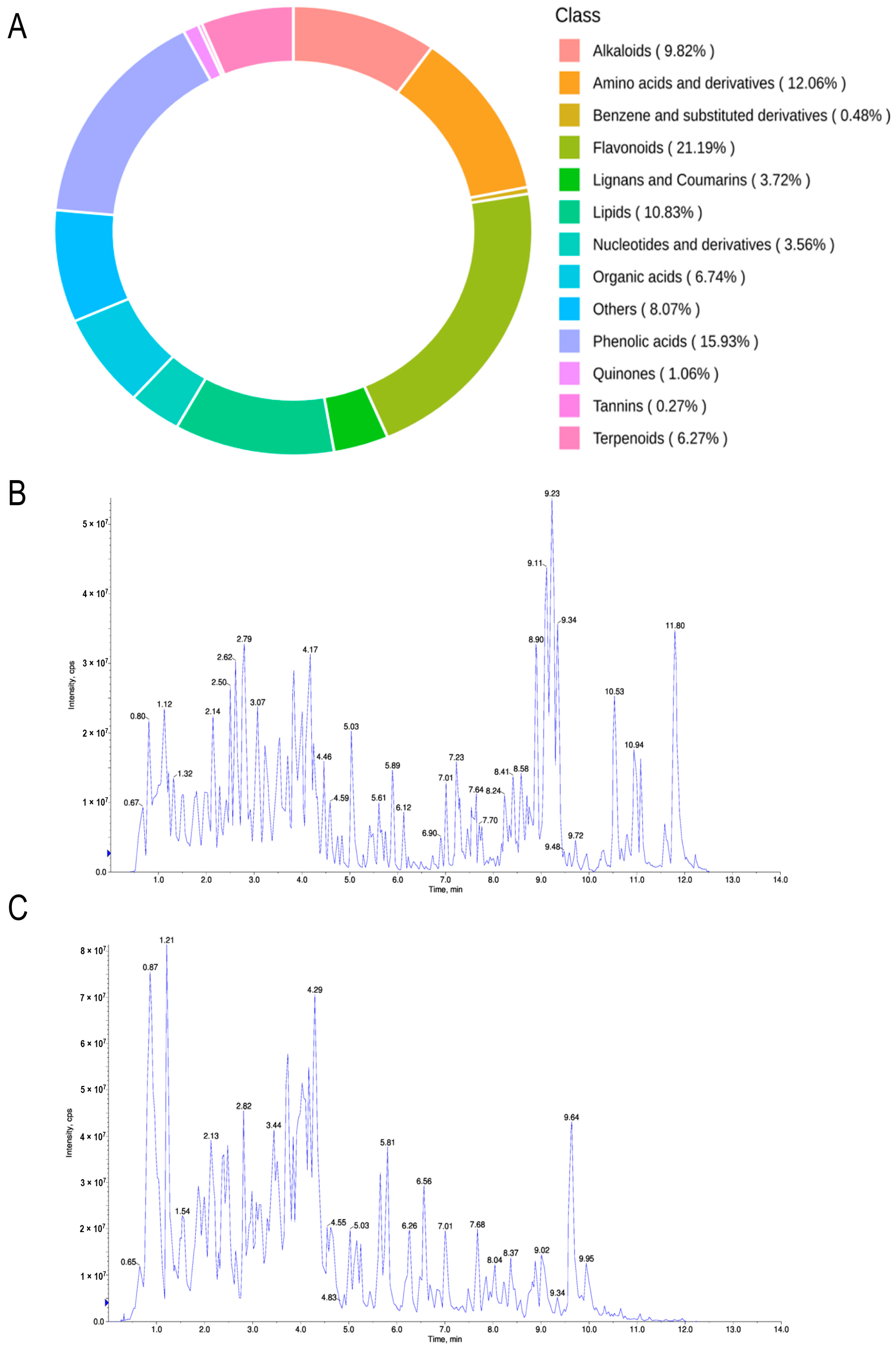{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Extraction and Identification of Seed Coat Components of Oryza sativa L. Indica
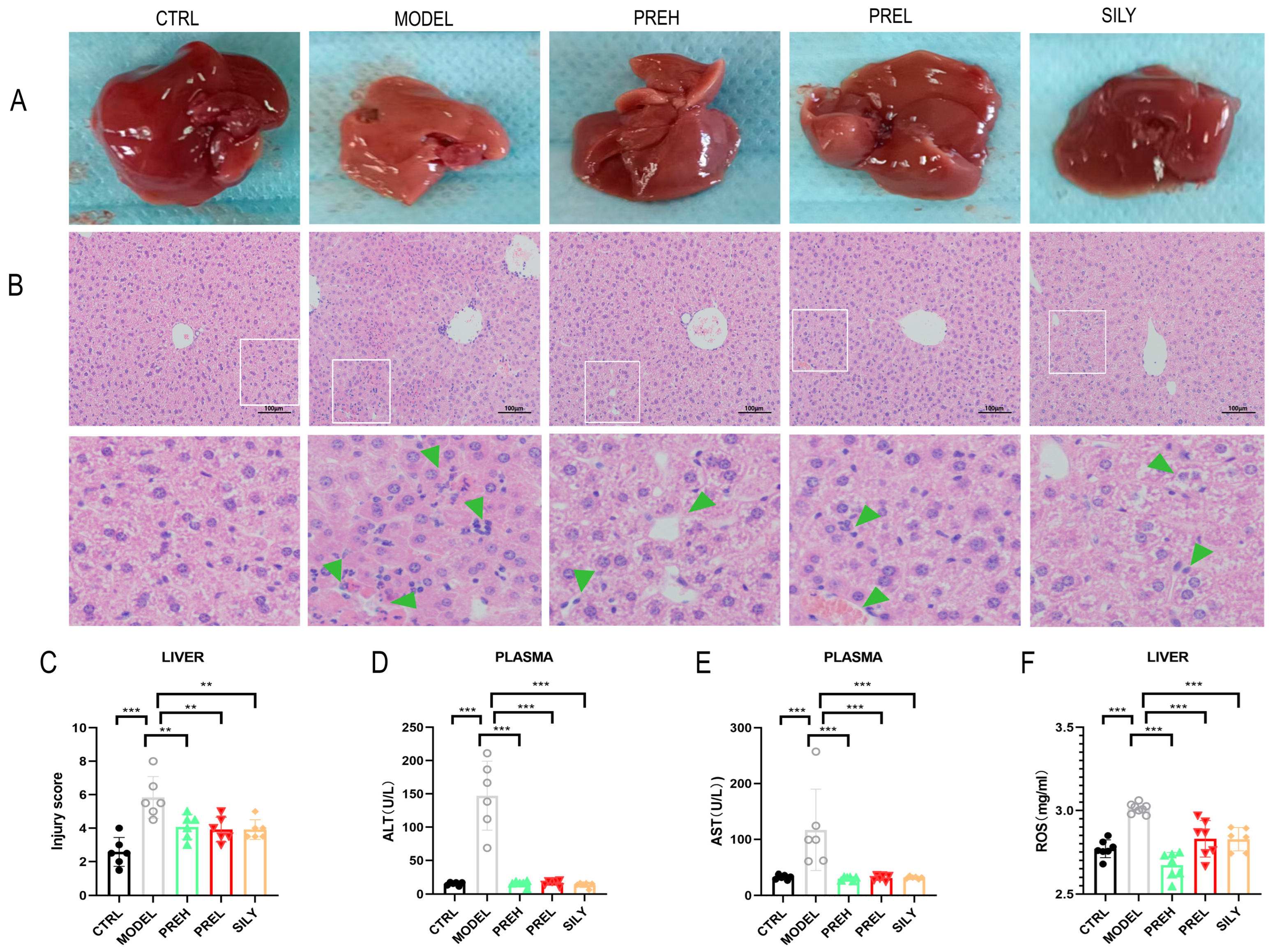
2.2. PRE Improved Pathological Liver Injury and Liver Function in AH Mice
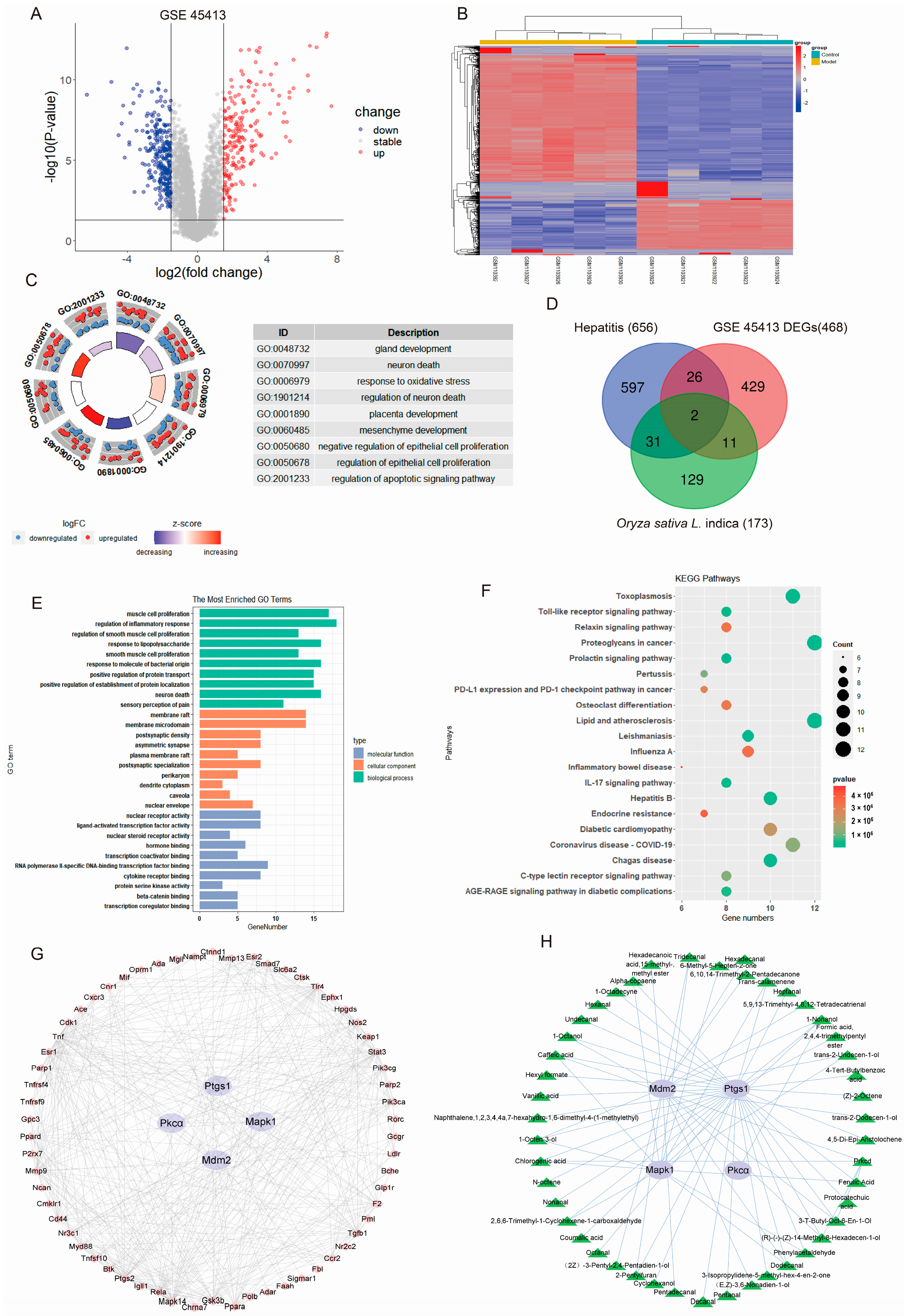
2.3. PRE May Ameliorate AH by Regulating Target Genes as Predicted by Bioinformatics
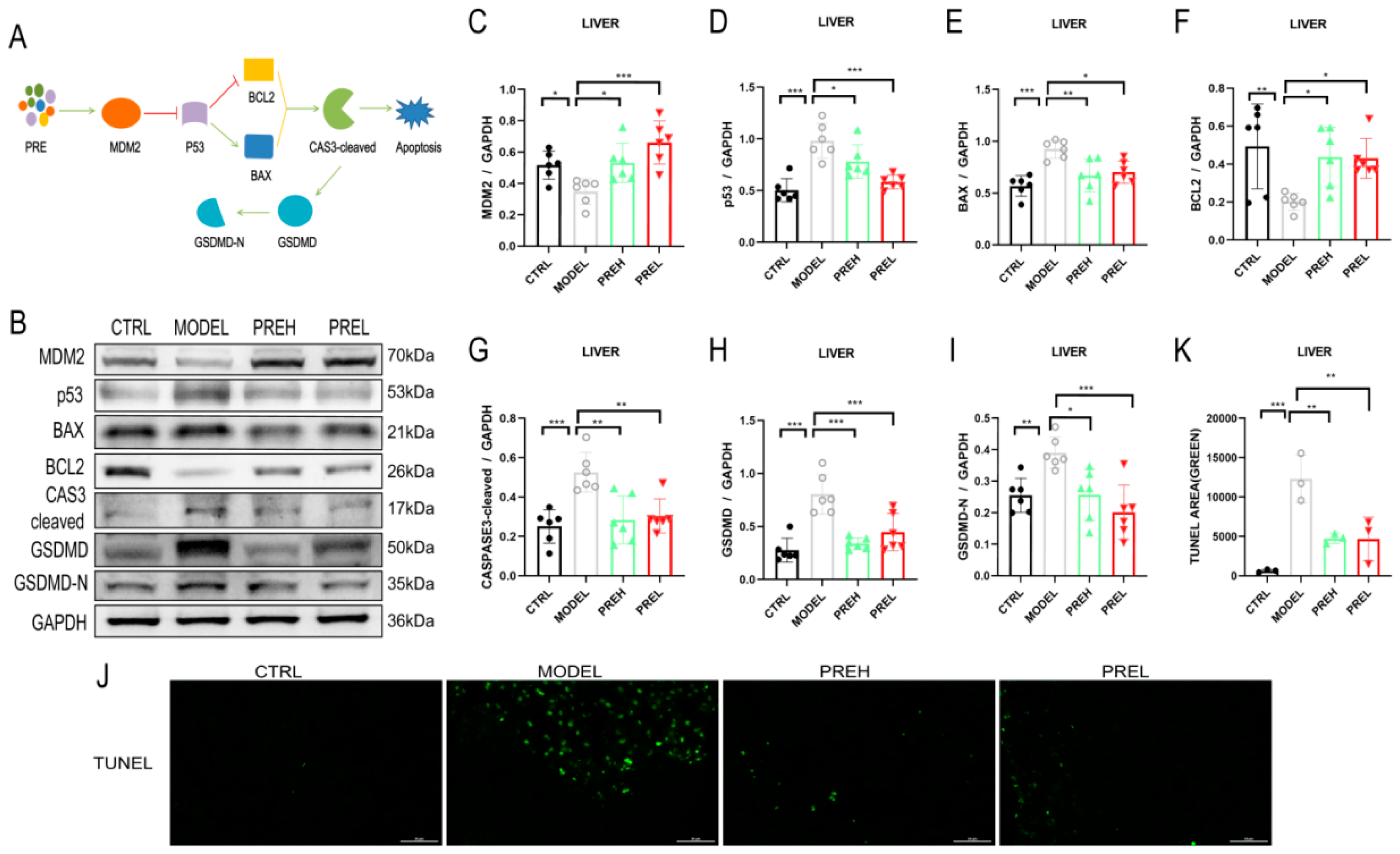
2.4. PRE Attenuated Liver Apoptosis in AH Mice through Upregulating MDM2
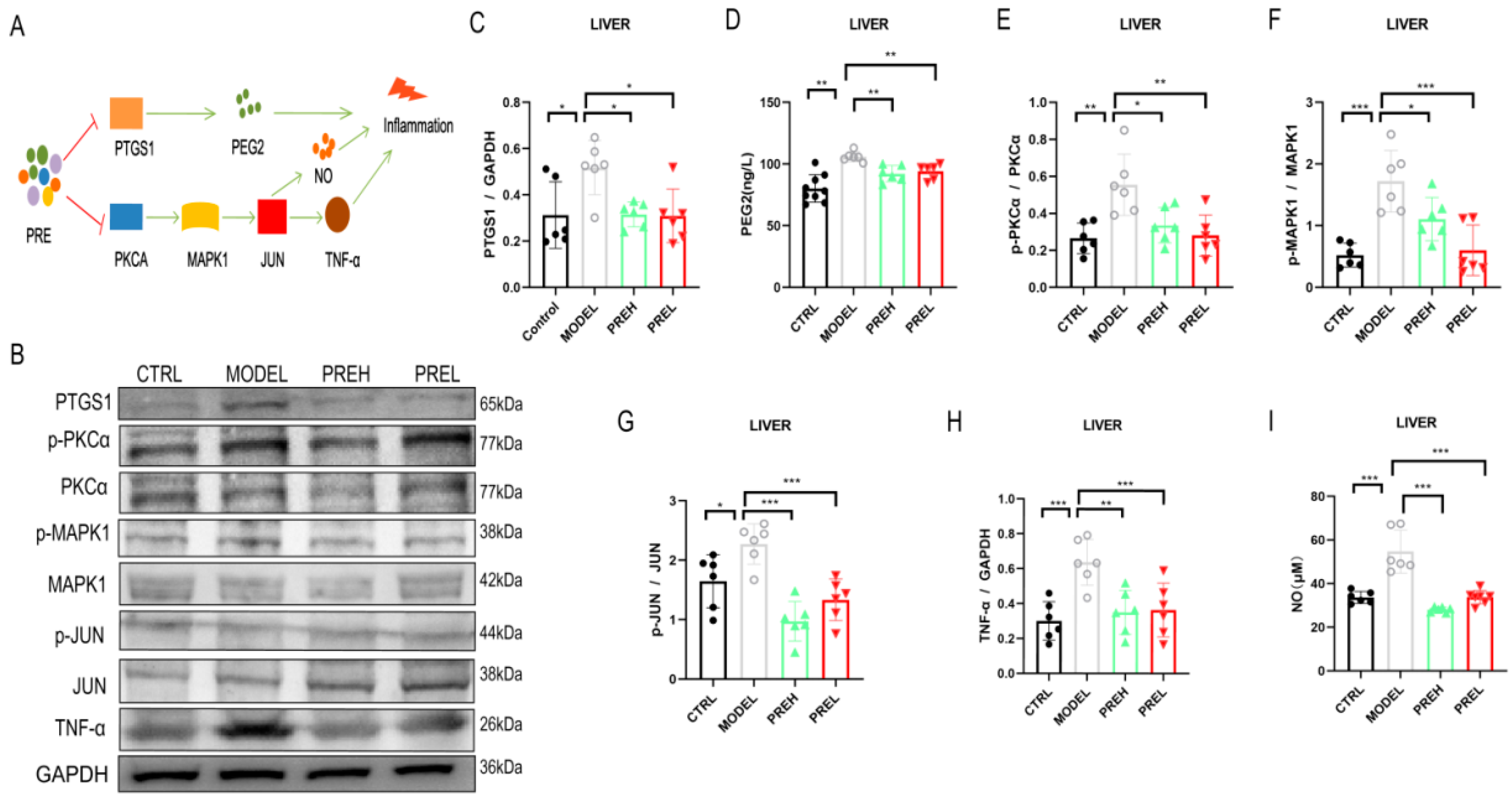
2.5. PRE Alleviated Inflammation in AH Mice through Downregulating PTGS1 and PKCα
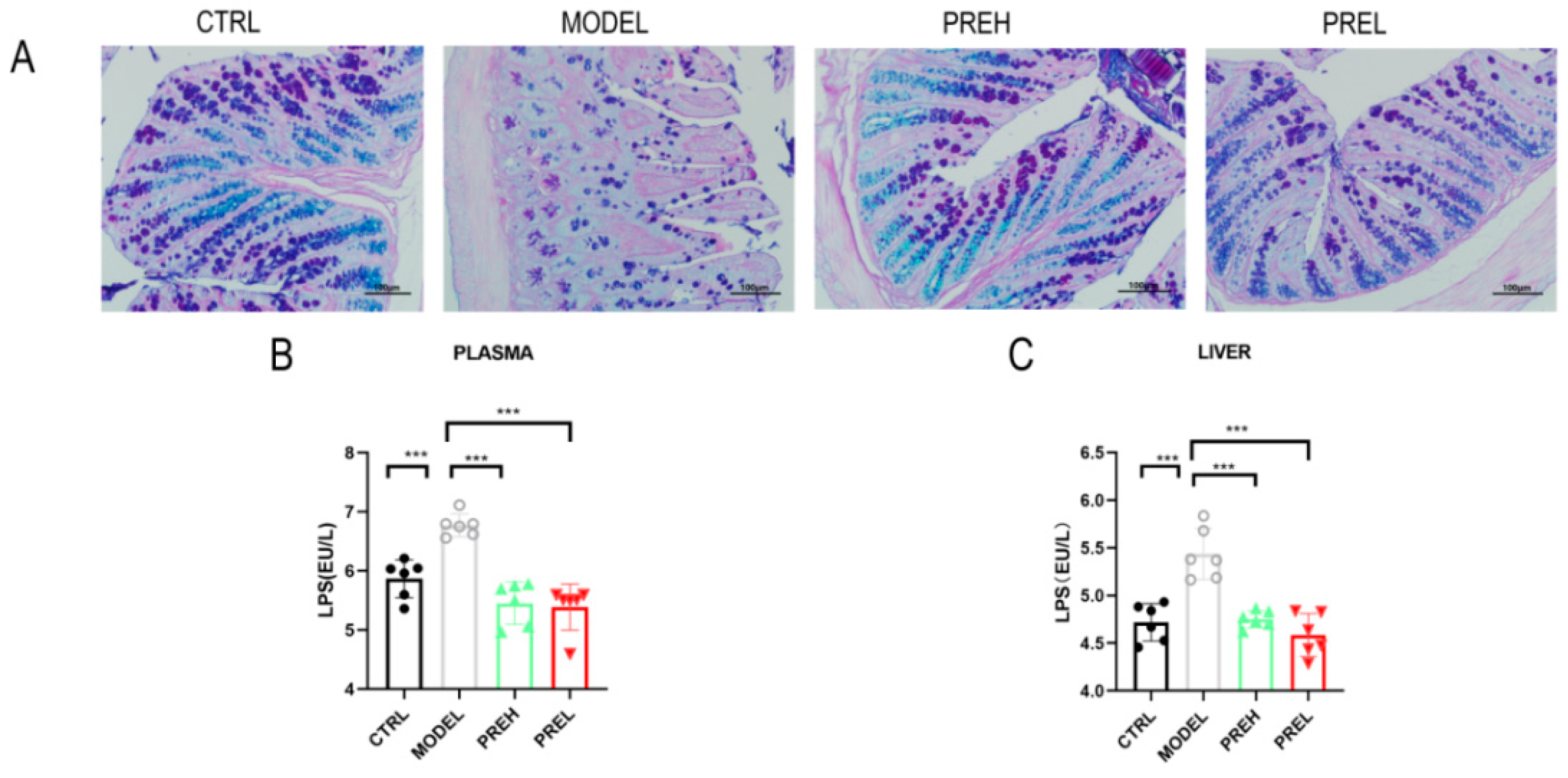
2.6. PRE Improved the Intestinal Barrier and Downregulated LPS Levels in AH Mice
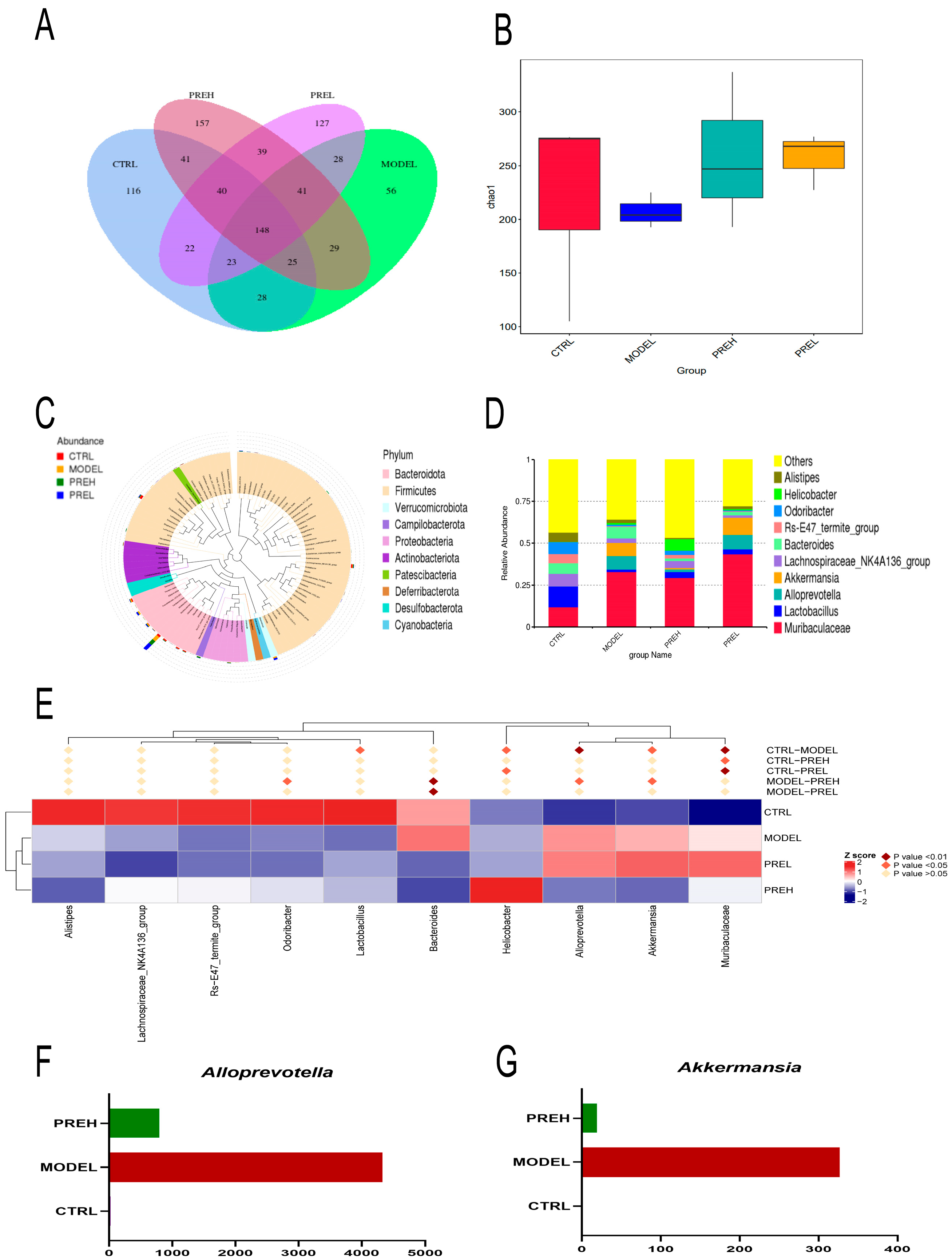
2.7. Effect of PRE on Gut Microbiota in AH Mice
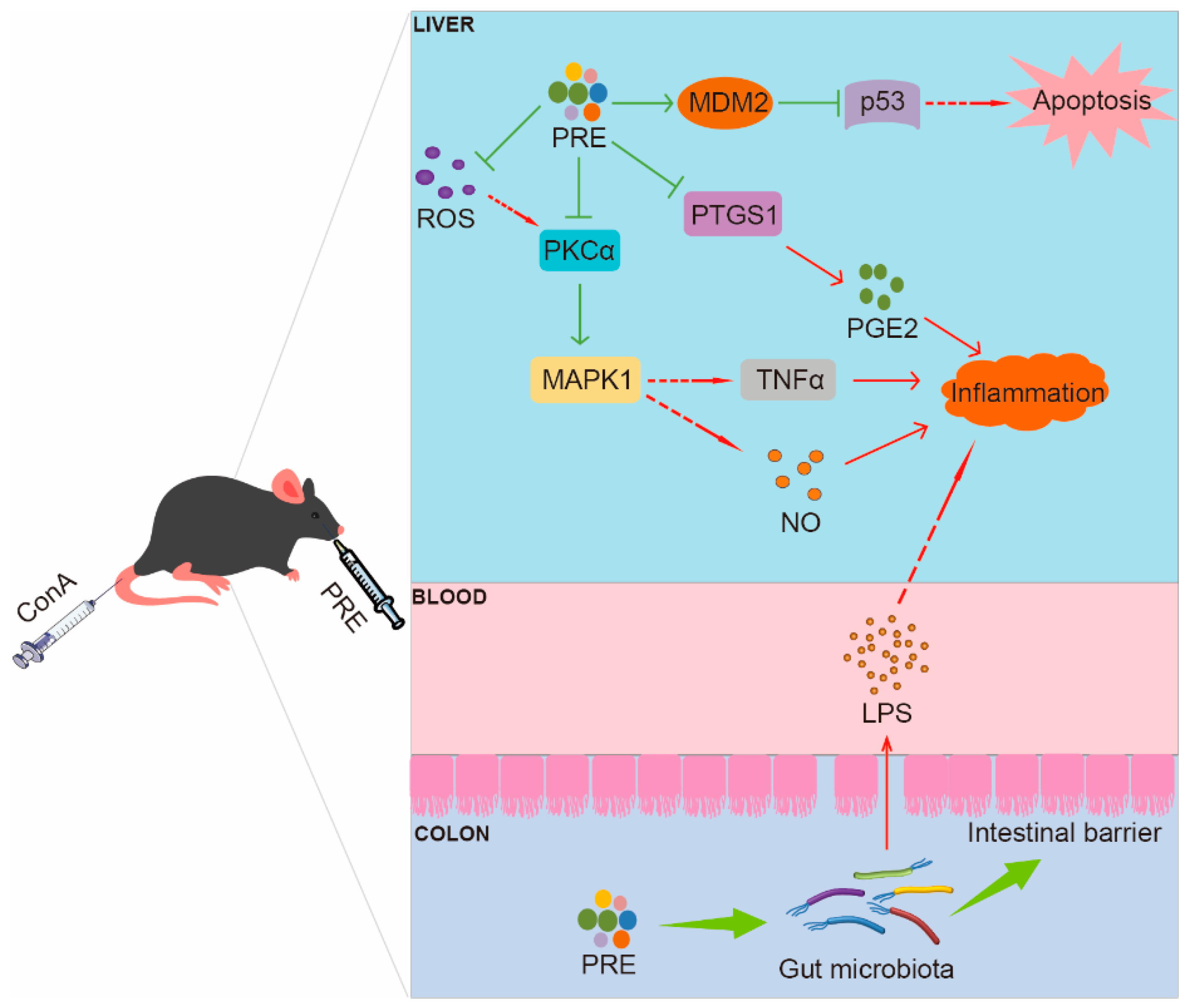
3. Discussion
4. Materials and Methods
4.1. Acquisition and Analysis of AH Differentially Expressed Genes (DEGs)
4.2. Potential Target Prediction of Oryza sativa L. Indica and AH
4.3. Extraction of Oryza sativa L. Indica Seed Coat
4.4. Widely Targeted Components Analysis of PRE
4.5. Animal Study
4.6. Histological Analysis
4.7. Detection of Cytokines
4.8. TUNEL Staining
4.9. 16S rRNA Sequencing of the Gut Microbiota
4.10. Western Blotting
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Muratori, P.; Granito, A.; Lenzi, M.; Muratori, L. Limitation of the simplified scoring system for the diagnosis of autoimmune Hepatitis with acute onset. Liver Int. 2021, 41, 529–534. [Google Scholar] [CrossRef]
- Liu, C.-H.; Kao, J.-H. Acute hepatitis C virus infection: Clinical update and remaining challenges. Clin. Mol. Hepatol. 2023, 29, 623–642. [Google Scholar] [CrossRef]
- Granito, A.; Muratori, P.; Muratori, L. Acute-on-chronic liver failure: A complex clinical entity in patients with autoimmune hepatitis. J. Hepatol. 2021, 75, 1503–1505. [Google Scholar] [CrossRef]
- Montrief, T.; Koyfman, A.; Long, B. Acute liver failure: A review for emergency physicians. Am. J. Emerg. Med. 2019, 37, 329–337. [Google Scholar] [CrossRef]
- Navarro, M.E.D.; Yao, C.C.; Whiteley, A.; Movahedi, B.; Devuni, D.; Barry, C.; Zacharias, I.; Theodoropoulos, N.M.; Bozorgzadeh, A.; Martins, P.N. Liver transplant evaluation for fulminant liver failure due to acute hepatitis A infection: Case series and literature review. Transpl. Infect. Dis. 2020, 23, e13476. [Google Scholar] [CrossRef]
- Bernal, W.; Wendon, J. Acute Liver Failure. New Engl. J. Med. 2013, 369, 2525–2534. [Google Scholar] [CrossRef]
- Heymann, F.; Hamesch, K.; Weiskirchen, R.; Tacke, F. The concanavalin A model of acute hepatitis in mice. Lab. Anim. 2015, 49 (Suppl. S1), 12–20. [Google Scholar] [CrossRef]
- Williams, R.; Riordan, S.M. Acute liver failure: Established and putative hepatitis viruses and therapeutic implications. J. Gastroenterol. Hepatol. 2002, 15, G17–G25. [Google Scholar] [CrossRef]
- Widjaja, A.A.; Singh, B.K.; Adami, E.; Viswanathan, S.; Dong, J.; D’Agostino, G.A.; Ng, B.; Lim, W.W.; Tan, J.; Paleja, B.S.; et al. Inhibiting Interleukin 11 Signaling Reduces Hepatocyte Death and Liver Fibrosis, Inflammation, and Steatosis in Mouse Models of Nonalcoholic Steatohepatitis. Gastroenterology 2019, 157, 777–792.e14. [Google Scholar] [CrossRef]
- Wang, H.; Feng, X.; Yan, W.; Tian, D. Regulatory T Cells in Autoimmune Hepatitis: Unveiling Their Roles in Mouse Models and Patients. Front. Immunol. 2020, 11, 575572. [Google Scholar] [CrossRef]
- Khan, H.A.; Ahmad, M.Z.; Khan, J.A.; Arshad, M.I. Crosstalk of liver immune cells and cell death mechanisms in different murine models of liver injury and its clinical relevance. Hepatobiliary Pancreat. Dis. Int. 2017, 16, 245–256. [Google Scholar] [CrossRef]
- Wang, X.; Wang, L.; Dong, R.; Huang, K.; Wang, C.; Gu, J.; Luo, H.; Liu, K.; Wu, J.; Sun, H.; et al. Luteolin ameliorates LPS-induced acute liver injury by inhibiting TXNIP-NLRP3 inflammasome in mice. Phytomedicine 2021, 87, 153586. [Google Scholar] [CrossRef]
- Zeng, Y.; Wang, L.; Du, J.; Liu, J.; Yang, S.; Pu, X.; Xiao, F. Elemental content in brown rice by inductively coupled plasma atomic emission spectroscopy reveals the evolution of asian cultivated rice. J. Integr. Plant Biol. 2009, 51, 466–475. [Google Scholar] [CrossRef]
- Yamuangmorn, S.; Prom, U.T.C. The Potential of High-Anthocyanin Purple Rice as a Functional Ingredient in Human Health. Antioxidants 2021, 10, 833. [Google Scholar] [CrossRef] [PubMed]
- Chariyakornkul, A.; Punvittayagul, C.; Taya, S.; Wongpoomchai, R. Inhibitory effect of purple rice husk extract on AFB(1)-induced micronucleus formation in rat liver through modulation of xenobiotic metabolizing enzymes. BMC Complement. Altern. Med. 2019, 19, 237. [Google Scholar] [CrossRef] [PubMed]
- Thipart, K.; Gruneck, L.; Phunikhom, K.; Sharpton, T.J.; Sattayasai, J.; Popluechai, S. Dark-purple rice extract modulates gut microbiota composition in acetic acid– and indomethacin-induced inflammatory bowel disease in rats. Int. Microbiol. 2022, 26, 423–434. [Google Scholar] [CrossRef]
- Feng, C.; Wan, H.; Zhang, Y.; Yu, L.; Shao, C.; He, Y.; Wan, H.; Jin, W. Neuroprotective Effect of Danhong Injection on Cerebral Ischemia-Reperfusion Injury in Rats by Activation of the PI3K-Akt Pathway. Front. Pharmacol. 2020, 11, 298. [Google Scholar] [CrossRef] [PubMed]
- Thomasova, D.; Mulay, S.R.; Bruns, H.; Anders, H.J. p53-independent roles of MDM2 in NF-kappaB signaling: Implications for cancer therapy, wound healing, and autoimmune diseases. Neoplasia 2012, 14, 1097–1101. [Google Scholar] [CrossRef] [PubMed]
- Overstreet, J.M.; Gifford, C.C.; Tang, J.; Higgins, P.J.; Samarakoon, R. Emerging role of tumor suppressor p53 in acute and chronic kidney diseases. Cell. Mol. Life Sci. CMLS 2022, 79, 474. [Google Scholar] [CrossRef]
- Liu, M.; Ai, W.; Sun, L.; Fang, F.; Wang, X.; Chen, S.; Wang, H. Triclosan-induced liver injury in zebrafish (Danio rerio) via regulating MAPK/p53 signaling pathway. Comp. Biochem. Physiology. Toxicol. Pharmacol. CBP 2019, 222, 108–117. [Google Scholar] [CrossRef]
- Al Humayed, S.; Al-Hashem, F.; Haidara, M.A.; El Karib, A.O.; Kamar, S.S.; Amin, S.N.; Al-Ani, B. Resveratrol Pretreatment Ameliorates p53-Bax Axis and Augments the Survival Biomarker B-Cell Lymphoma 2 Modulated by Paracetamol Overdose in a Rat Model of Acute Liver Injury. Pharmacology 2020, 105, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.N.; Tan, Y.; Xiao, X.C.; Li, Q.; Wu, Q.; Peng, Y.Y.; Ren, J.; Dong, M.L. Deletion of TLR4 attenuates lipopolysaccharide-induced acute liver injury by inhibiting inflammation and apoptosis. Acta Pharmacol. Sin. 2021, 42, 1610–1619. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.-W.; Li, W.; Zheng, X.-J.; Liu, J.-Y.; Yang, Y.-H.; Li, S.; Zhang, S.; Fu, W.-Q.; Xiao, B.; Wang, J.-H.; et al. Benzimidazoles induce concurrent apoptosis and pyroptosis of human glioblastoma cells via arresting cell cycle. Acta Pharmacol. Sin. 2021, 43, 194–208. [Google Scholar] [CrossRef]
- Zheng, Y.; Cui, B.; Sun, W.; Wang, S.; Huang, X.; Gao, H.; Gao, F.; Cheng, Q.; Lu, L.; An, Y.; et al. Potential Crosstalk between Liver and Extra-liver Organs in Mouse Models of Acute Liver Injury. Int. J. Biol. Sci. 2020, 16, 1166–1179. [Google Scholar] [CrossRef]
- Chen, K.; Li, J.; Li, S.; Feng, J.; Wu, L.; Liu, T.; Zhang, R.; Xu, S.; Cheng, K.; Zhou, Y.; et al. 15d-PGJ2 alleviates ConA-induced acute liver injury in mice by up-regulating HO-1 and reducing hepatic cell autophagy. Biomed. Pharmacother. Biomed. Pharmacother. 2016, 80, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kornienko, N. Electrocatalytic carbon dioxide reduction in acid. Chem Catal. 2022, 2, 29–38. [Google Scholar] [CrossRef]
- Tian, L.; Cao, J.; Zhao, T.; Liu, Y.; Khan, A.; Cheng, G. The Bioavailability, Extraction, Biosynthesis and Distribution of Natural Dihydrochalcone: Phloridzin. Int. J. Mol. Sci. 2021, 22, 962. [Google Scholar] [CrossRef] [PubMed]
- Bakrim, W.B.; Nurcahyanti, A.D.R.; Dmirieh, M.; Mahdi, I.; Elgamal, A.M.; El Raey, M.A.; Wink, M.; Sobeh, M. Phytochemical Profiling of the Leaf Extract of Ximenia americana var. caffra and Its Antioxidant, Antibacterial, and Antiaging Activities In Vitro and in Caenorhabditis elegans: A Cosmeceutical and Dermatological Approach. Oxidative Med. Cell. Longev. 2022, 2022, 3486257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lv, W.; Fu, Y.; Li, Y.; Wang, J.; Chen, D.; Han, X.; Li, Z. Hepatoprotective Activity of Ethanol Extract of Rice Solid-State Fermentation of Ganoderma tsugae against CCl(4)-Induced Acute Liver Injury in Mice. Molecules 2022, 27, 5347. [Google Scholar] [CrossRef]
- Hu, J.J.W.; Pan, C.W.H.; Lin, M.X. Isovitexin alleviates liver injury induced by lipopolysaccharide/d-galactosamine by activating Nrf2 and inhibiting NF-kappaB activation. Microb. Pathog. 2018, 119, 86–92. [Google Scholar] [CrossRef]
- Sreekanth, G.P.; Chuncharunee, A.; Cheunsuchon, B.; Noisakran, S.; Yenchitsomanus, P.T.; Limjindaporn, T. JNK1/2 inhibitor reduces dengue virus-induced liver injury. Antivir. Res. 2017, 141, 7–18. [Google Scholar] [CrossRef]
- Han, C.; Sheng, J.; Pei, H.; Sheng, Y.; Wang, J.; Zhou, X.; Li, W.; Cao, C.; Yang, Y. Environmental toxin chlorpyrifos induces liver injury by activating P53-mediated ferroptosis via GSDMD-mtROS. Ecotoxicol. Environ. Saf. 2023, 257, 114938. [Google Scholar] [CrossRef]
- Yan, H.; Wu, W.; Hu, Y.; Li, J.; Xu, J.; Chen, X.; Xu, Z.; Yang, X.; Yang, B.; He, Q.; et al. Regorafenib inhibits EphA2 phosphorylation and leads to liver damage via the ERK/MDM2/p53 axis. Nat. Commun. 2023, 14, 2756. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chin, L.-C.; Kimura, K.; Nakahata, Y. Human Placental Extract Delays In Vitro Cellular Senescence through the Activation of NRF2-Mediated Antioxidant Pathway. Antioxidants 2022, 11, 1545. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lv, Q.; Ma, J.; Liu, Y. PLCγ2 promotes apoptosis while inhibits proliferation in rat hepatocytes through PKCD/JNK MAPK and PKCD/p38 MAPK signalling. Cell Prolif. 2018, 51, e12437. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-J.; Yang, B.-C.; Liu, M.-Y. Lead Increases Lipopolysaccharide-Induced Liver Injury through Tumor Necrosis Factor-α Overexpression by Monocytes/Macrophages: Role of Protein Kinase C and p42/44 Mitogen-Activated Protein Kinase. Environ. Health Perspect. 2006, 114, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.P.; Huang, X.; Yang, M.W.; Fang, S.Y.; Hong, F.F.; Yang, S.L. Effects of non-drug treatment on liver cells apoptosis during hepatic ischemia-reperfusion injury. Life Sci. 2021, 275, 119321. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, Y.; Aguilar, A.; Bernard, D.; Yang, C.Y. Targeting the MDM2-p53 Protein-Protein Interaction for New Cancer Therapy: Progress and Challenges. Cold Spring Harb. Perspect. Med. 2017, 7, a026245. [Google Scholar] [CrossRef]
- Jimenez-Castro, M.B.; Cornide-Petronio, M.E.; Gracia-Sancho, J.; Peralta, C. Inflammasome-Mediated Inflammation in Liver Ischemia-Reperfusion Injury. Cells 2019, 8, 1131. [Google Scholar] [CrossRef]
- Ahn, M.; Park, J.S.; Chae, S.; Kim, S.; Moon, C.; Hyun, J.W.; Shin, T. Hepatoprotective effects of Lycium chinense Miller fruit and its constituent betaine in CCl4-induced hepatic damage in rats. Acta Histochem. 2014, 116, 1104–1112. [Google Scholar] [CrossRef]
- Guo, C.; Guo, D.; Fang, L.; Sang, T.; Wu, J.; Guo, C.; Wang, Y.; Wang, Y.; Chen, C.; Chen, J.; et al. Ganoderma lucidum polysaccharide modulates gut microbiota and immune cell function to inhibit inflammation and tumorigenesis in colon. Carbohydr. Polym. 2021, 267, 118231. [Google Scholar] [CrossRef]
- Ulger Toprak, N.; Duman, N.; Sacak, B.; Ozkan, M.C.; Sayın, E.; Mulazimoglu, L.; Soyletir, G. Alloprevotella rava isolated from a mixed infection of an elderly patient with chronic mandibular osteomyelitis mimicking oral squamous cell carcinoma. New Microbes New Infect. 2021, 42, 100880. [Google Scholar] [CrossRef]
- Gao, Y.; Ma, X.; Zhou, Y.; Li, Y.; Xiang, D. Dietary supplementation of squalene increases the growth performance of early-weaned piglets by improving gut microbiota, intestinal barrier, and blood antioxidant capacity. Front. Vet. Sci. 2022, 9, 995548. [Google Scholar] [CrossRef]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Susceptibility. Cell 2016, 167, 1339–1353.e21. [Google Scholar] [CrossRef]
- Seregin, S.S.; Golovchenko, N.; Schaf, B.; Chen, J.; Pudlo, N.A.; Mitchell, J.; Baxter, N.T.; Zhao, L.; Schloss, P.D.; Martens, E.C.; et al. NLRP6 Protects Il10 Mice from Colitis by Limiting Colonization of Akkermansia muciniphila. Cell Rep. 2017, 19, 733–745. [Google Scholar] [CrossRef]
- Song, Y.; Shan, B.; Zeng, S.; Zhang, J.; Jin, C.; Liao, Z.; Wang, T.; Zeng, Q.; He, H.; Wei, F.; et al. Raw and wine processed Schisandra chinensis attenuate anxiety like behavior via modulating gut microbiota and lipid metabolism pathway. J. Ethnopharmacol. 2021, 266, 113426. [Google Scholar] [CrossRef]
- Song, X.; Wang, W.; Ding, S.; Wang, Y.; Ye, L.; Chen, X.; Ma, H. Exploring the potential antidepressant mechanisms of puerarin: Anti-inflammatory response via the gut-brain axis. J. Affect. Disord. 2022, 310, 459–471. [Google Scholar] [CrossRef]
- Fujita, T.; Soontrapa, K.; Ito, Y.; Iwaisako, K.; Moniaga, C.S.; Asagiri, M.; Majima, M.; Narumiya, S. Hepatic Stellate Cells Relay Inflammation Signaling From Sinusoids to Parenchyma in Mouse Models of Immune-Mediated Hepatitis. Hepatology 2016, 63, 1325–1339. [Google Scholar] [CrossRef]
- Li, W.H.; Han, J.R.; Ren, P.P.; Xie, Y.; Jiang, D.Y. Exploration of the mechanism of Zisheng Shenqi decoction against gout arthritis using network pharmacology. Comput. Biol. Chem. 2021, 90, 107358. [Google Scholar] [CrossRef]
- Pinero, J.; Bravo, A.; Queralt-Rosinach, N.; Gutierrez-Sacristan, A.; Deu-Pons, J.; Centeno, E.; Garcia-Garcia, J.; Sanz, F.; Furlong, L.I. DisGeNET: A comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res. 2017, 45, D833–D839. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Bae, I.Y.; An, J.S.; Oh, I.K.; Lee, H.G. Optimized preparation of anthocyanin-rich extract from black rice and its effects on in vitro digestibility. Food Sci. Biotechnol. 2017, 26, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, M.; He, J.; Lv, K.; Wang, M.; Guan, W.; Liu, J.; Tao, Y.; Li, S.; Ho, C.T.; et al. Protective effect of pterostilbene on concanavalin A-induced acute liver injury. Food Funct. 2019, 10, 7308–7314. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Nonalcoholic Steatohepatitis Clinical Research, N., Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- He, S.; Gao, Q.; Wu, X.; Shi, J.; Zhang, Y.; Yang, J.; Li, X.; Du, S.; Zhang, Y.; Yu, J. NAD+ameliorates endotoxin-induced acute kidney injury in a sirtuin1–dependent manner via GSK-3β/Nrf2 signalling pathway. J. Cell. Mol. Med. 2022, 26, 1979–1993. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Li, Y.; Guo, S.; Chen, Y.; Yin, H.; Li, Y.; Cheng, G.; Tian, L. Oryza sativa L. Indica Seed Coat Ameliorated Concanavalin A—Induced Acute Hepatitis in Mice via MDM2/p53 and PKCα/MAPK1 Signaling Pathways. Int. J. Mol. Sci. 2023, 24, 14503. https://doi.org/10.3390/ijms241914503
Zhao Z, Li Y, Guo S, Chen Y, Yin H, Li Y, Cheng G, Tian L. Oryza sativa L. Indica Seed Coat Ameliorated Concanavalin A—Induced Acute Hepatitis in Mice via MDM2/p53 and PKCα/MAPK1 Signaling Pathways. International Journal of Molecular Sciences. 2023; 24(19):14503. https://doi.org/10.3390/ijms241914503
Chicago/Turabian StyleZhao, Zhiye, Ye Li, Shancheng Guo, Yuxu Chen, Haiaolong Yin, Yaxian Li, Guiguang Cheng, and Lei Tian. 2023. "Oryza sativa L. Indica Seed Coat Ameliorated Concanavalin A—Induced Acute Hepatitis in Mice via MDM2/p53 and PKCα/MAPK1 Signaling Pathways" International Journal of Molecular Sciences 24, no. 19: 14503. https://doi.org/10.3390/ijms241914503