
Scopolamine-Induced Memory Impairment in Mice: Effects of PEA-OXA on Memory Retrieval and Hippocampal LTP

 , ,
, , 
 , and
, and 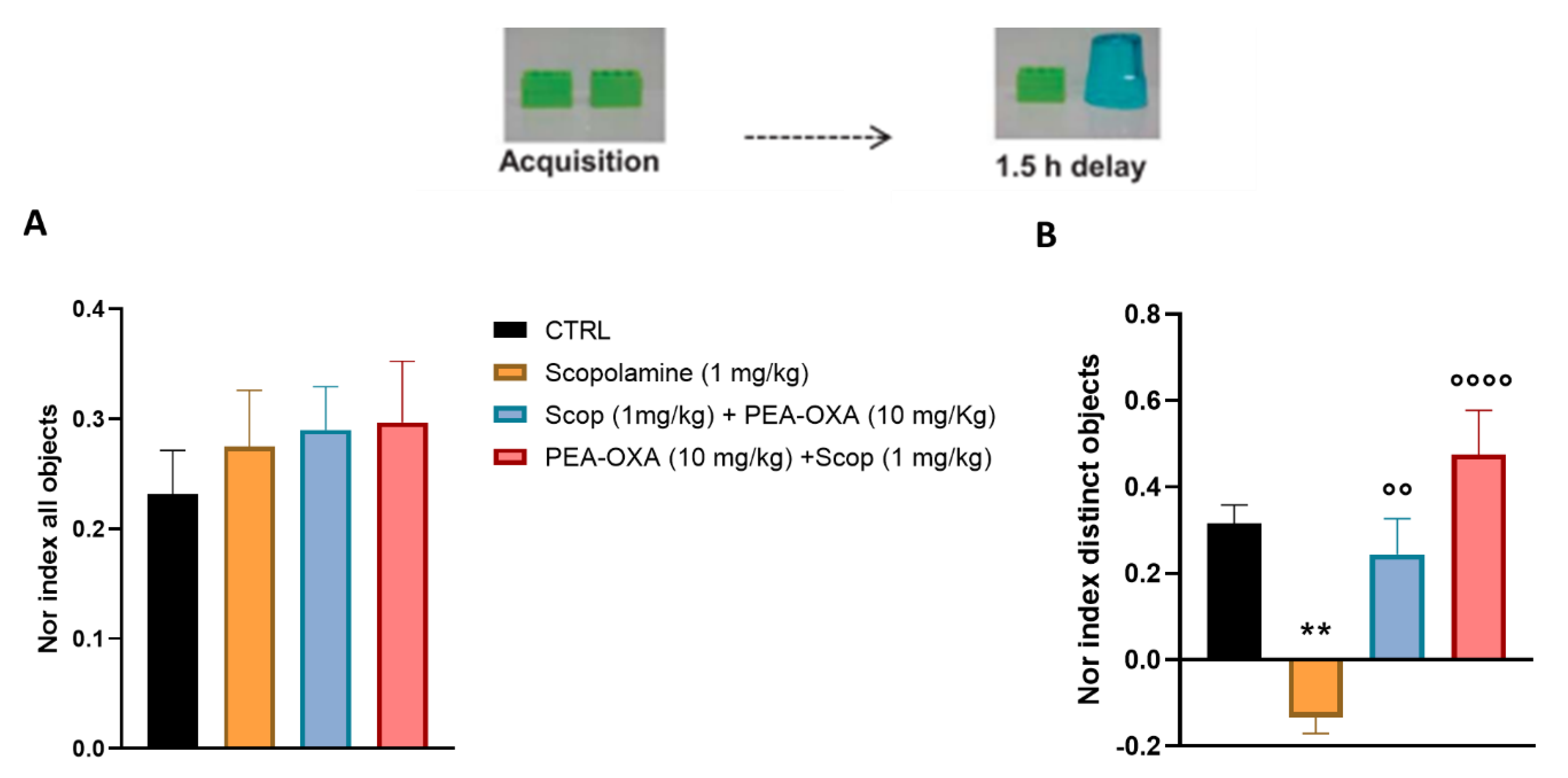{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. PEA-OXA Prevents Scopolamine-Induced Learning and Cognitive Damage
2.2. PEA-OXA Restores the Impaired LTP in the LEC-DG Circuit in Scopolamine-Injected Mice
2.3. Effect of PEA-OXA before and after Ccopolamine on the Expression Levels of Ach and ChAT
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Statistical Analysis
4.3. Experimental Design
4.4. Y-Maze Forced Alternation Test and Scopolamine-Induced Amnesia Protocol
4.5. Novel Object Recognition Test and Scopolamine-Induced Amnesia Protocol
4.6. In Vivo Recording of Long-Term Potentiation (LTP) in the LEC-DG Pathway
4.7. Immunoassays (ELISA)
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef] [PubMed]
- The Lancet Public Health Reinvigorating the public health response to dementia. Lancet Public Heal. 2021, 6, e696. [CrossRef] [PubMed]
- Pantoni, L. Transient global amnesia: An intriguing yet benign disturbance. Arq. Neuro-Psiquiatr. 2019, 77, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Agosti, C.; Borroni, B.; Akkawi, N.; Padovani, A. Cerebrovascular Risk Factors and Triggers in Transient Global Amnesia Patients with and without Jugular Valve Incompetence: Results from a Sample of 243 Patients. Eur. Neurol. 2010, 63, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Blake, M.; Krawczyk, M.; Baratti, C.; Boccia, M. Neuropharmacology of memory consolidation and reconsolidation: Insights on central cholinergic mechanisms. J. Physiol. 2014, 108, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Haroutunian, V.; Santucci, A.C.; Davis, K.L. Chapter 35 Implications of Multiple Transmitter System Lesions for Cholinomi-metic Therapy in Alzheimer’s Disease. Progress Brain Res. 1990, 84, 333–346. [Google Scholar]
- Tellez, S.; Colpaert, F.; Marien, M. α2-Adrenoceptor modulation of cortical acetylcholine release in vivo. Neuroscience 1999, 89, 1041–1050. [Google Scholar] [CrossRef]
- Agency for Healthcare Research and Quality (US). Eisenberg Center at Oregon Health & Science University Off -Label Use of Atypical Antipsychotic Drugs: A Summary for Clinicians and Policymakers. In Comparative Effectiveness Review Summary Guides for Clinicians; AHRQ Comparative Effec-tiveness Reviews; Agency for Healthcare Research and Quality (US): Rockville, MD, USA, 2007. [Google Scholar]
- Neyama, H.; Dozono, N.; Uchida, H.; Ueda, H. Mirtazapine, an α2 Antagonist-Type Antidepressant, Reverses Pain and Lack of Morphine Analgesia in Fibromyalgia-Like Mouse Models. Experiment 2020, 375, 1–9. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Siracusa, R.; Cordaro, M.; Crupi, R.; Peritore, A.F.; Gugliandolo, E.; D’Amico, R.; Petrosino, S.; Evangelista, M.; Di Paola, R.; et al. N-Palmitoylethanolamine-oxazoline (PEA-OXA): A new therapeutic strategy to reduce neuroinflammation, oxidative stress associated to vascular dementia in an experimental model of repeated bilateral common carotid arteries occlusion. Neurobiol. Dis. 2019, 125, 77–91. [Google Scholar] [CrossRef]
- Kolahdouzan, M.; Hamadeh, M.J. The neuroprotective effects of caffeine in neurodegenerative diseases. CNS Neurosci. Ther. 2017, 23, 272–290. [Google Scholar] [CrossRef]
- Zhou, Y.; Fang, S.-H.; Ye, Y.-L.; Chu, L.-S.; Zhang, W.-P.; Wang, M.-L.; Wei, E.-Q. Caffeic acid ameliorates early and delayed brain injuries after focal cerebral ischemia in rats. Acta Pharmacol. Sin. 2006, 27, 1103–1110. [Google Scholar] [CrossRef]
- Trinh, K.; Andrews, L.; Krause, J.; Hanak, T.; Lee, D.; Gelb, M.; Pallanck, L. Decaffeinated Coffee and Nicotine-Free Tobacco Provide Neuroprotection in Drosophila Models of Parkinson’s Disease through an NRF2-Dependent Mechanism. J. Neurosci. 2010, 30, 5525–5532. [Google Scholar] [CrossRef]
- Hwang, Y.P.; Jeong, H.G. The coffee diterpene kahweol induces heme oxygenase-1 via the PI3K and p38/Nrf2 pathway to protect human dopaminergic neurons from 6-hydroxydopamine-derived oxidative stress. FEBS Lett. 2008, 582, 2655–2662. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Cordaro, M.; Bruschetta, G.; Crupi, R.; Pascali, J.; Alfonsi, D.; Marcolongo, G.; Cuzzocrea, S. 2-pentadecyl-2-oxazoline: Identification in coffee, synthesis and activity in a rat model of carrageenan-induced hindpaw inflammation. Pharmacol. Res. 2016, 108, 23–30. [Google Scholar] [CrossRef]
- Boccella, S.; Guida, F.; Iannotta, M.; Iannotti, F.A.; Infantino, R.; Ricciardi, F.; Cristiano, C.; Vitale, R.M.; Amodeo, P.; Marabese, I.; et al. 2-Pentadecyl-2-oxazoline ameliorates memory impairment and depression-like behaviour in neuropathic mice: Possible role of adrenergic alpha2- and H3 histamine autoreceptors. Mol. Brain 2021, 14, 1–24. [Google Scholar] [CrossRef]
- Boccella, S.; Iannotta, M.; Cristiano, C.; Iannotti, F.A.; Del Bello, F.; Guida, F.; Belardo, C.; Infantino, R.; Ricciardi, F.; Giannella, M.; et al. Treatment With 2-Pentadecyl-2-Oxazoline Restores Mild Traumatic Brain Injury-Induced Sensorial and Neuropsychiatric Dysfunctions. Front. Pharmacol. 2020, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Infantino, R.; Boccella, S.; Scuteri, D.; Perrone, M.; Ricciardi, F.; Vitale, R.; Bonsale, R.; Parente, A.; Allocca, I.; Virtuoso, A.; et al. 2-pentadecyl-2-oxazoline prevents cognitive and social behaviour impairments in the Amyloid β-induced Alzheimer-like mice model: Bring the α2 adrenergic receptor back into play. BioMedicine 2022, 156, 113844. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.J.; Kim, J.; Shim, J.; Kim, C.-Y.; Jang, J.-H.; Lee, K.W.; Lee, H.J. Decaffeinated coffee prevents scopolamine-induced memory impairment in rats. Behav. Brain Res. 2013, 245, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Ardila, A.; Moreno, C. Scopolamine intoxication as a model of transient global amnesia. Brain Cogn. 1991, 15, 236–245. [Google Scholar] [CrossRef]
- Farmer, P.; Pugin, J. β-Adrenergic agonists exert their “anti-inflammatory” effects in monocytic cells through the IκB/NF-κB pathway. Am. J. Physiol. Cell. Mol. Physiol. 2000, 279, L675–L682. [Google Scholar] [CrossRef]
- Cheon, S.Y.; Koo, B.-N.; Kim, S.Y.; Kam, E.H.; Nam, J.; Kim, E.J. Scopolamine promotes neuroinflammation and delirium-like neuropsychiatric disorder in mice. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bonaz, B.; Sinniger, V.; Pellissier, S. Anti-inflammatory properties of the vagus nerve: Potential therapeutic implications of vagus nerve stimulation. J. Physiol. 2016, 594, 5781–5790. [Google Scholar] [CrossRef] [PubMed]
- Frinchi, M.; Nuzzo, D.; Scaduto, P.; Di Carlo, M.; Massenti, M.F.; Belluardo, N.; Mudò, G. Anti-inflammatory and antioxidant effects of muscarinic acetylcholine receptor (mAChR) activation in the rat hippocampus. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef]
- Minett, T.; Cfas, M.; Classey, J.; Matthews, F.E.; Fahrenhold, M.; Taga, M.; Brayne, C.; Ince, P.G.; Nicoll, J.A.R.; Boche, D. Microglial immunophenotype in dementia with Alzheimer’s pathology. J. Neuroinflammation 2016, 13, 135. [Google Scholar] [CrossRef] [PubMed]
- Ennerfelt, H.; Holliday, C.; Shapiro, D.A.; Zengeler, K.E.; Bolte, A.C.; Ulland, T.K.; Lukens, J.R. CARD9 attenuates Aβ pathology and modifies microglial responses in an Alzheimer’s disease mouse model. Proc. Natl. Acad. Sci. USA 2023, 120, e2303760120. [Google Scholar] [CrossRef]
- Boccella, S.; Cristiano, C.; Romano, R.; Iannotta, M.; Belardo, C.; Farina, A.; Guida, F.; Piscitelli, F.; Palazzo, E.; Mazzitelli, M.; et al. Ultra-micronized palmitoylethanolamide rescues the cognitive decline-associated loss of neural plasticity in the neuropathic mouse entorhinal cortex-dentate gyrus pathway. Neurobiol. Dis. 2018, 121, 106–119. [Google Scholar] [CrossRef]
- Kodama, D.; Ono, H.; Tanabe, M. Altered hippocampal long-term potentiation after peripheral nerve injury in mice. Eur. J. Pharmacol. 2007, 574, 127–132. [Google Scholar] [CrossRef]
- Wohleb, E.S.; Wu, M.; Gerhard, D.M.; Taylor, S.R.; Picciotto, M.R.; Alreja, M.; Duman, R.S. GABA interneurons mediate the rapid antidepressant-like effects of scopolamine. J. Clin. Investig. 2016, 126, 2482–2494. [Google Scholar] [CrossRef]
- Toide, K. Effects of scopolamine on extracellular acetylcholine and choline levels and on spontaneous motor activity in freely moving rats measured by brain dialysis. Pharmacol. Biochem. Behav. 1989, 33, 109–113. [Google Scholar] [CrossRef]
- Lim, D.W.; Son, H.J.; Um, M.Y.; Kim, I.-H.; Han, D.; Cho, S.; Lee, C.-H. Enhanced Cognitive Effects of Demethoxycurcumin, a Natural Derivative of Curcumin on Scopolamine-Induced Memory Impairment in Mice. Molecules 2016, 21, 1022. [Google Scholar] [CrossRef]
- Wolf, A.; Bauer, B.; Abner, E.L.; Ashkenazy-Frolinger, T.; Hartz, A.M.S. A Comprehensive Behavioral Test Battery to Assess Learning and Memory in 129S6/Tg2576 Mice. PLoS ONE 2016, 11, e0147733. [Google Scholar] [CrossRef] [PubMed]
- Anisman, H. Dissociation of disinhibitory effects of scopolamine: Strain and task factors. Pharmacol. Biochem. Behav. 1975, 3, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Busquet, P.; Capurro, V.; Cavalli, A.; Piomelli, D.; Reggiani, A.; Bertorelli, R. Synergistic Effects of Galantamine and Memantine in Attenuating Scopolamine-Induced Amnesia in Mice. J. Pharmacol. Sci. 2012, 120, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Bolz, L.; Heigele, S.; Bischofberger, J. Running Improves Pattern Separation during Novel Object Recognition. Brain Plast. 2015, 1, 129–141. [Google Scholar] [CrossRef]
- Wu, Y.; Kawakami, R.; Shinohara, Y.; Fukaya, M.; Sakimura, K.; Mishina, M.; Watanabe, M.; Ito, I.; Shigemoto, R. Target-Cell-Specific Left-Right Asymmetry of NMDA Receptor Content in Schaffer Collateral Synapses in ϵ1/NR2A Knock-Out Mice. J. Neurosci. 2005, 25, 9213–9226. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belardo, C.; Boccella, S.; Perrone, M.; Fusco, A.; Morace, A.M.; Ricciardi, F.; Bonsale, R.; ELBini-Dhouib, I.; Guida, F.; Luongo, L.; et al. Scopolamine-Induced Memory Impairment in Mice: Effects of PEA-OXA on Memory Retrieval and Hippocampal LTP. Int. J. Mol. Sci. 2023, 24, 14399. https://doi.org/10.3390/ijms241814399
Belardo C, Boccella S, Perrone M, Fusco A, Morace AM, Ricciardi F, Bonsale R, ELBini-Dhouib I, Guida F, Luongo L, et al. Scopolamine-Induced Memory Impairment in Mice: Effects of PEA-OXA on Memory Retrieval and Hippocampal LTP. International Journal of Molecular Sciences. 2023; 24(18):14399. https://doi.org/10.3390/ijms241814399
Chicago/Turabian StyleBelardo, Carmela, Serena Boccella, Michela Perrone, Antimo Fusco, Andrea Maria Morace, Federica Ricciardi, Roozbe Bonsale, Ines ELBini-Dhouib, Francesca Guida, Livio Luongo, and et al. 2023. "Scopolamine-Induced Memory Impairment in Mice: Effects of PEA-OXA on Memory Retrieval and Hippocampal LTP" International Journal of Molecular Sciences 24, no. 18: 14399. https://doi.org/10.3390/ijms241814399