

Integrated Analysis of Morphological, Physiological, Anatomical and Molecular Responses of Cassava Seedlings to Different Light Qualities
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
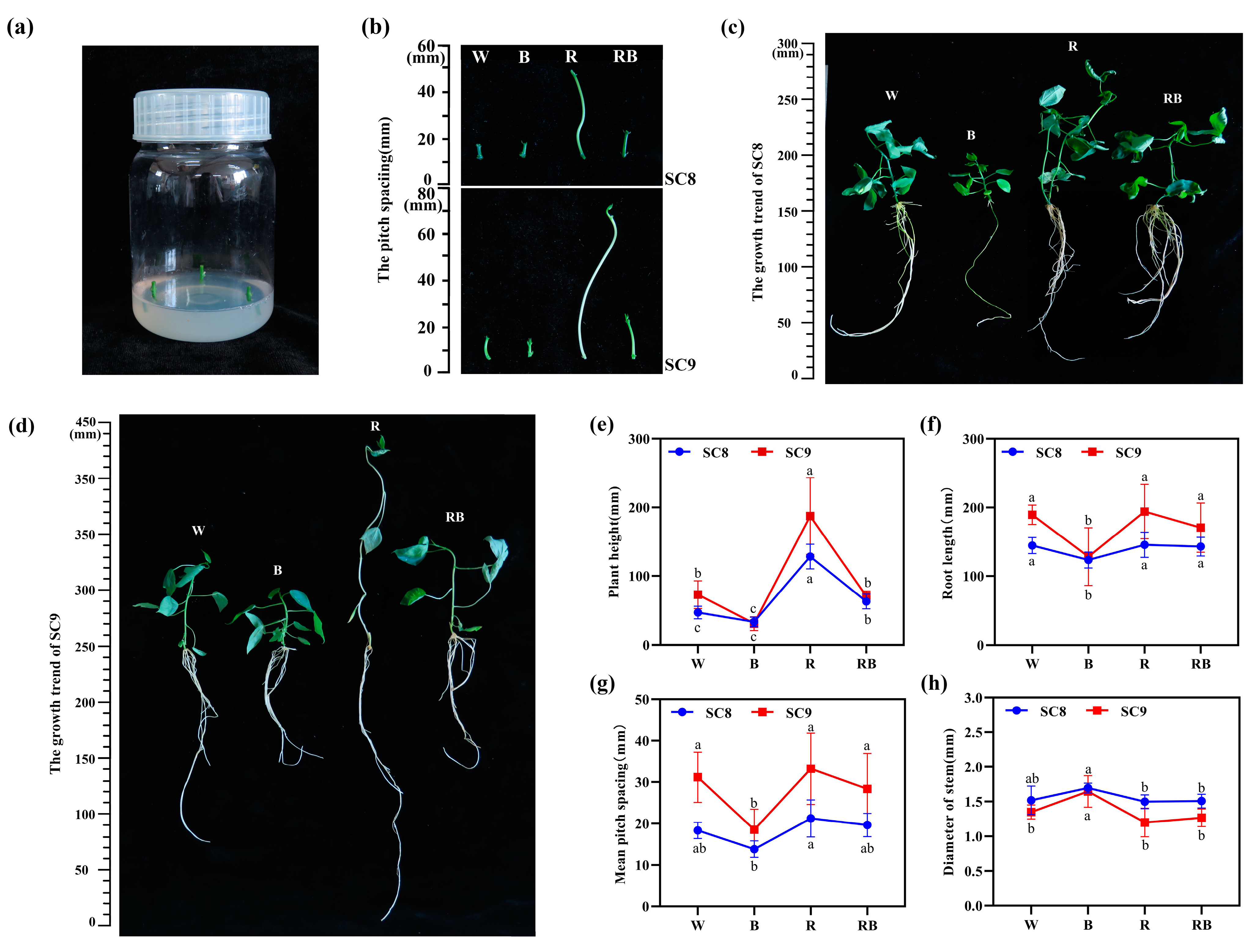
2.1. Effect of Light Quality on In Vitro Growth of Cassava
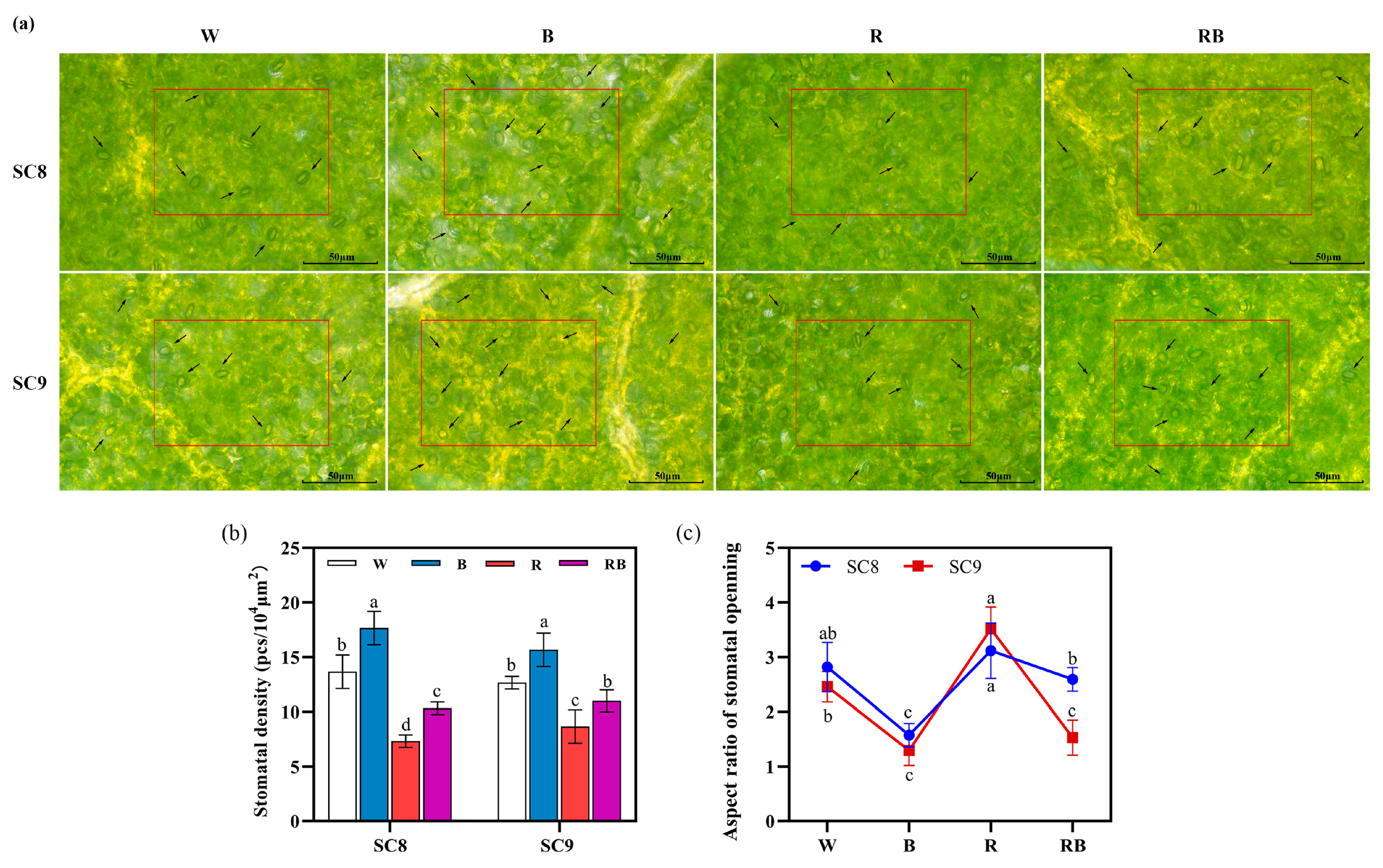
2.2. Effect of Light Quality on Stomatal Density and Stomatal Aperture of Cassava
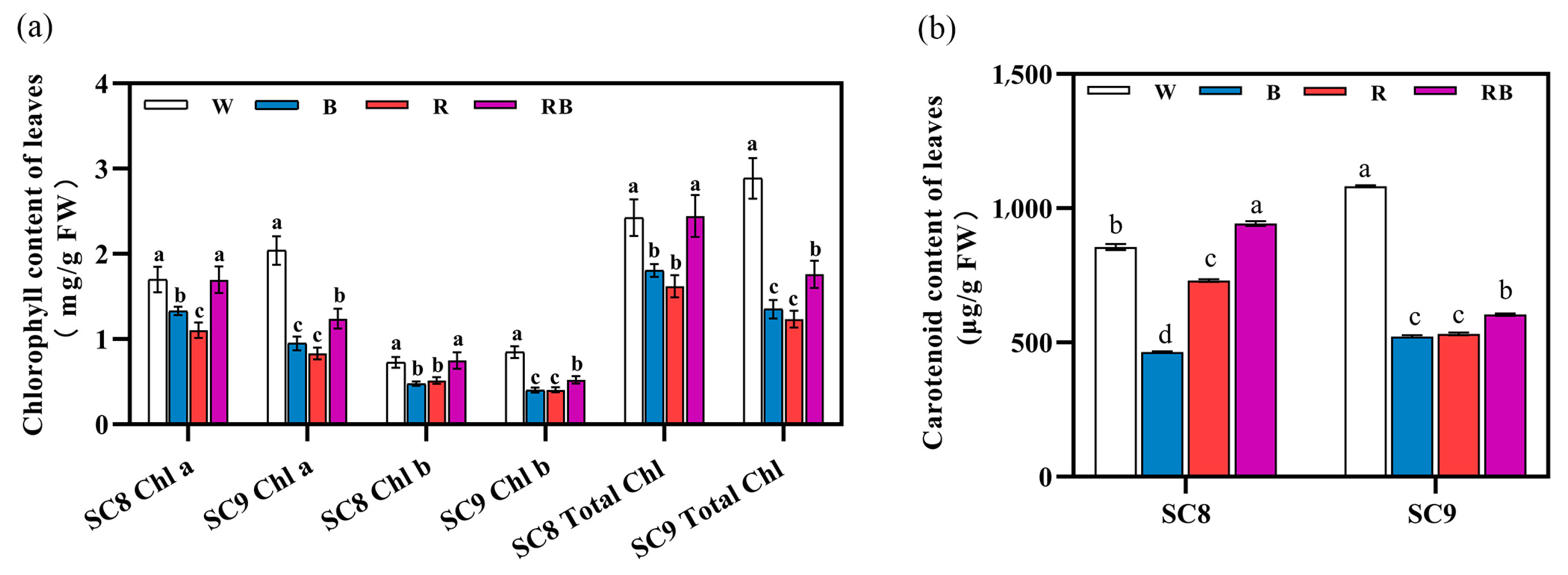
2.3. Effect of Light Quality on Photosynthetic Pigment Content
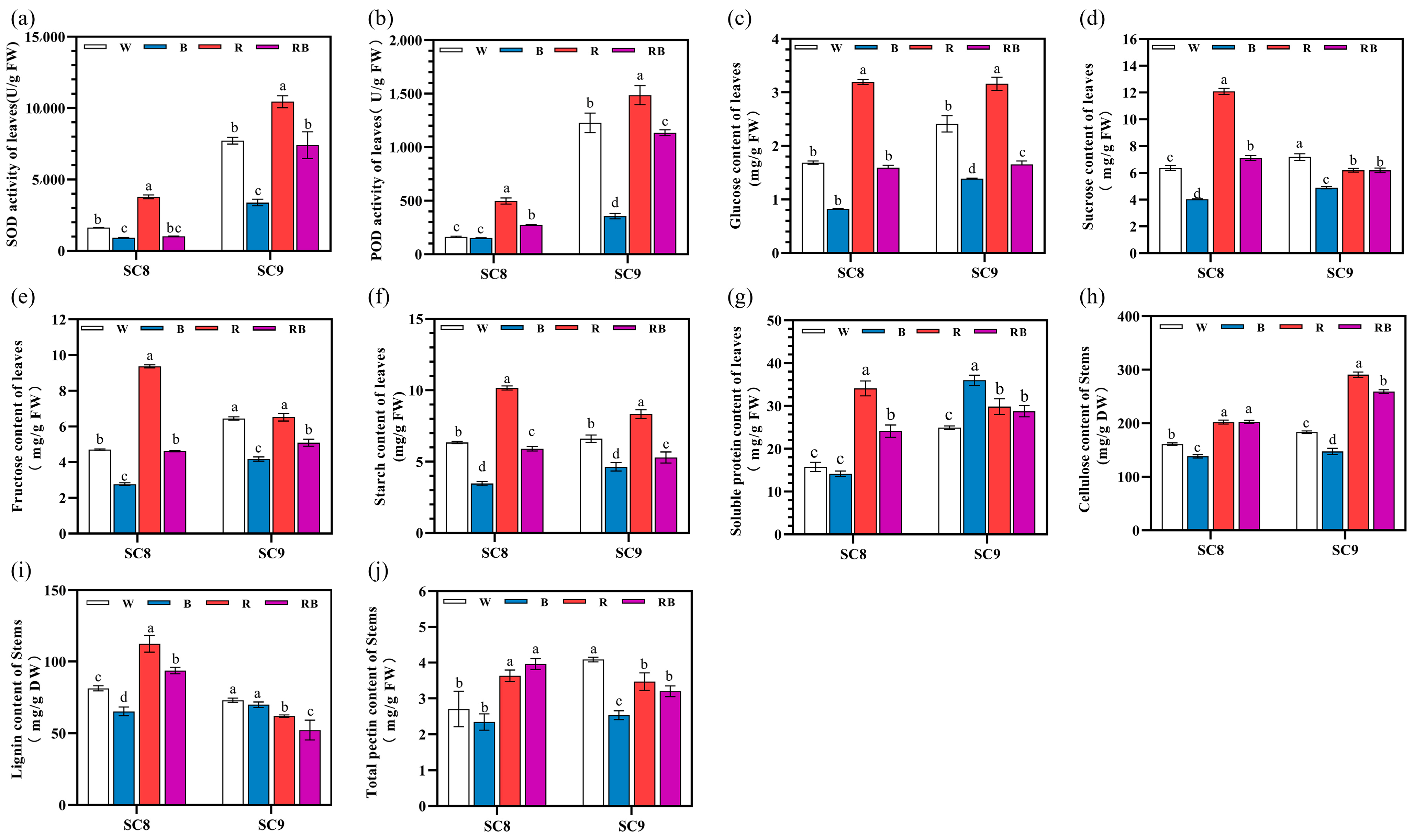
2.4. Effects of Different Light Qualities on Antioxidant Enzyme Activities, Carbohydrate and Soluble Protein Contents
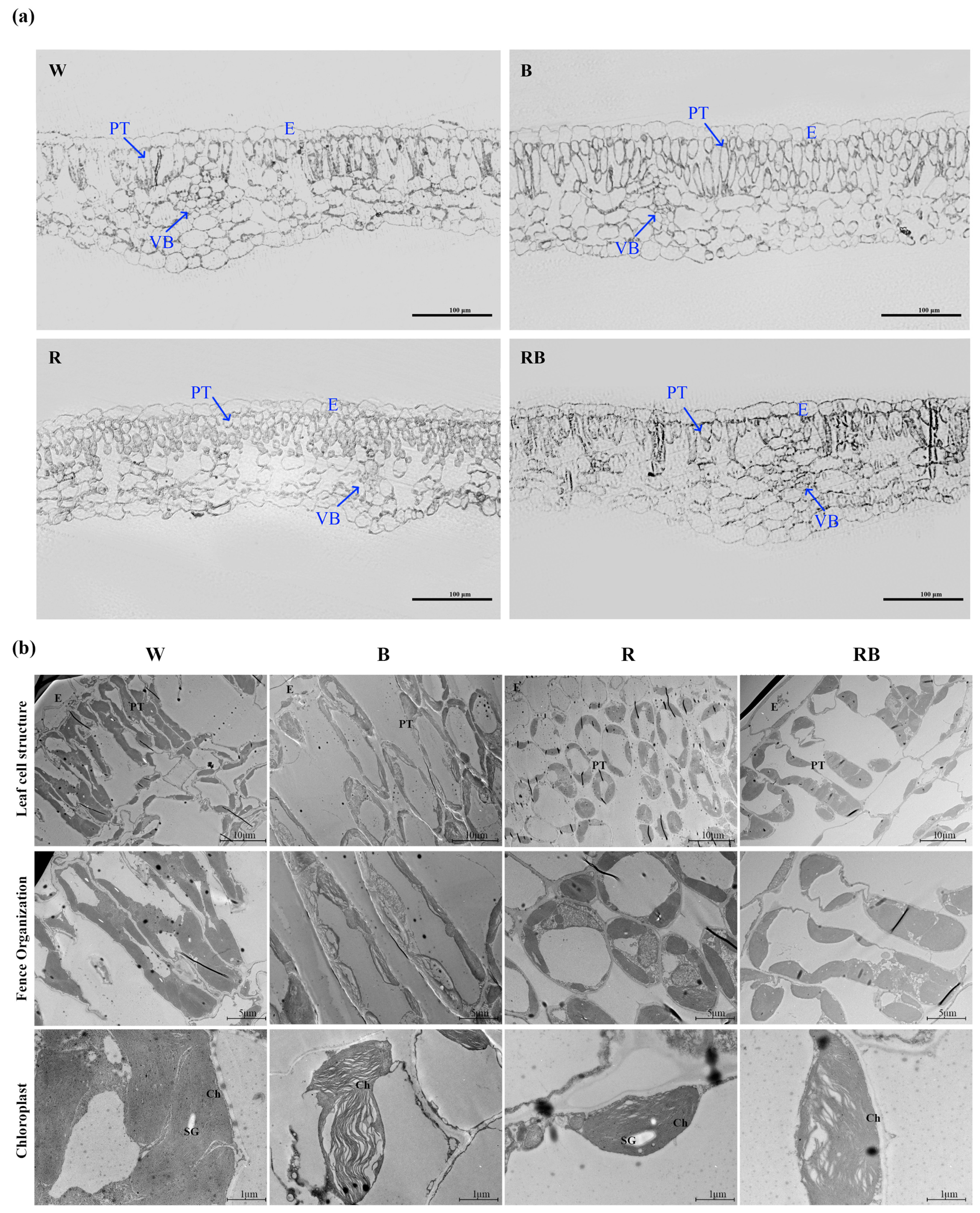
2.5. Effects of Different Light Qualities on Cell Structure of Cassava
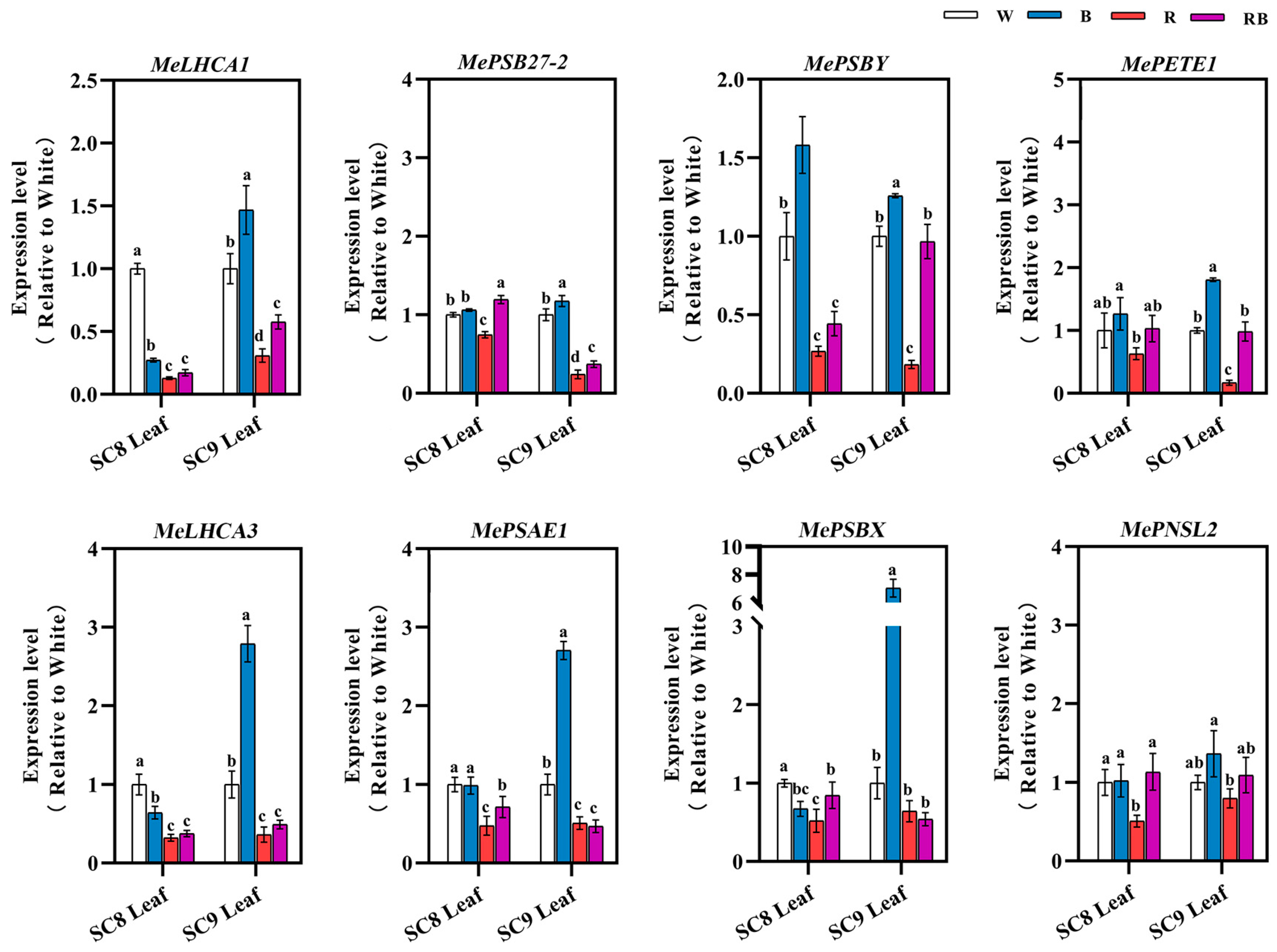
2.6. Effects of Different Light Qualities on the Expression of Photosynthesis-Related Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Light Treatments
4.2. Observation of Growth and Stomatal Characteristics of Cassava
4.3. Determination of Photosynthetic Pigment Content
4.4. Determination of SOD and POD Activities
4.5. Determination of Soluble Protein and Carbohydrate Content
4.6. Microstructure Measurements
4.7. Expression Analysis of Photosynthesis-Related Genes
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sonnewald, U.; Fernie, A.R.; Gruissem, W.; Schläpfer, P.; Anjanappa, R.B.; Chang, S.H.; Ludewig, F.; Rascher, U.; Muller, O.; van Doorn, A.M. The Cassava Source–Sink project: Opportunities and challenges for crop improvement by metabolic engineering. Plant J. 2020, 103, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Li, R.M.; Yuan, S.; Zhou, Y.J.; Wang, S.J.; Zhou, Q.; Ding, Z.P.; Wang, Y.J.; Yao, Y.; Liu, J.; Guo, J.C. Comparative Transcriptome Profiling of Cassava Tuberous Roots in Response to Postharvest Physiological Deterioration. Int. J. Mol. Sci. 2023, 24, 246. [Google Scholar] [CrossRef] [PubMed]
- Sessou, A.F.; Kahia, J.W.; Houngue, J.A.; Ateka, E.M.; Dadjo, C.; Ahanhanzo, C. In vitro propagation of three mosaic disease resistant cassava cultivars. BMC Biotechnol. 2020, 20, 51. [Google Scholar] [CrossRef]
- Eni, A.O.; Efekemo, O.P.; Onile-ere, O.A.; Pita, J.S. South West and North Central Nigeria: Assessment of cassava mosaic disease and field status of African cassava mosaic virus and East African cassava mosaic virus. Ann. Appl. Biol. 2021, 178, 466–479. [Google Scholar] [CrossRef] [PubMed]
- Cacaï, G.; Adoukonou-Sagbadja, H.; Kumulugui, B.; Ovono, P.; Houngue, J.; Ahanhanzo, C. Eradication of cassava (Manihot esculenta) mosaic symptoms through thermotherapy and meristems cultured in vitro. Int. J. Agron. Plant Prod. 2013, 4, 3697–3701. [Google Scholar]
- Tuo, D.C.; Zhao, G.Y.; Yan, P.; Li, R.M.; Chen, X.; Wang, W.Q.; Li, X.Y.; Yao, Y.; Cui, H.G.; Shen, W.T.; et al. First Report of Cassava common mosaic virus Infecting Cassava in Mainland China. Plant Dis. 2020, 104, 997. [Google Scholar] [CrossRef]
- Lin, X.; Li, R.; Zhou, Y.; Tang, F.; Wang, Y.; Lu, X.; Wang, S.; Yao, Y.; Liu, J.; Hu, X. Overexpression of cassava MeAnn2 enhances the salt and IAA tolerance of transgenic Arabidopsis. Plants 2021, 10, 941. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, R.; Zhou, Y.; Fernie, A.R.; Ding, Z.; Zhou, Q.; Che, Y.; Yao, Y.; Liu, J.; Wang, Y. Integrated Characterization of Cassava (Manihot esculenta) Pectin Methylesterase (MePME) Genes to Filter Candidate Gene Responses to Multiple Abiotic Stresses. Plants 2023, 12, 2529. [Google Scholar] [CrossRef] [PubMed]
- Omongo, C.A.; Opio, S.M.; Bayiyana, I.; Otim, M.H.; Omara, T.; Wamani, S.; Ocitti, P.; Bua, A.; Macfadyen, S.; Colvin, J. African cassava whitefly and viral disease management through timed application of imidacloprid. Crop Prot. 2022, 158, 106015. [Google Scholar] [CrossRef]
- Silvestri, C.; Caceres, M.E.; Ceccarelli, M.; Pica, A.L.; Rugini, E.; Cristofori, V. Influence of Continuous Spectrum Light on Morphological Traits and Leaf Anatomy of Hazelnut Plantlets. Front. Plant Sci. 2019, 10, 1318. [Google Scholar] [CrossRef]
- Zheng, L.; Van Labeke, M.-C. Long-Term Effects of Red- and Blue-Light Emitting Diodes on Leaf Anatomy and Photosynthetic Efficiency of Three Ornamental Pot Plants. Front. Plant Sci. 2017, 8, 917. [Google Scholar] [CrossRef]
- Fan, C.; Manivannan, A.; Wei, H. Light Quality-Mediated Influence of Morphogenesis in Micropropagated Horticultural Crops: A Comprehensive Overview. BioMed Res. Int. 2022, 2022, 4615079. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.; Yin, Y.; Chen, X.; Qiu, B. Chlorophylls. In Research Methods of Environmental Physiology in Aquatic Sciences; Gao, K., Hutchins, D.A., Beardall, J., Eds.; Springer: Singapore, 2021; pp. 95–106. [Google Scholar]
- Karimi, M.; Ahmadi, N.; Ebrahimi, M. Red LED light promotes biomass, flowering and secondary metabolites accumulation in hydroponically grown Hypericum perforatum L.(cv. Topas). Ind. Crops Prod. 2022, 175, 114239. [Google Scholar] [CrossRef]
- Yap, E.S.P.; Uthairatanakij, A.; Laohakunjit, N.; Jitareerat, P.; Vaswani, A.; Magana, A.A.; Morre, J.; Maier, C.S. Plant growth and metabolic changes in ‘Super Hot’chili fruit (Capsicum annuum) exposed to supplemental LED lights. Plant Sci. 2021, 305, 110826. [Google Scholar] [CrossRef] [PubMed]
- Lazzarin, M.; Meisenburg, M.; Meijer, D.; Van Ieperen, W.; Marcelis, L.; Kappers, I.; Van der Krol, A.; van Loon, J.; Dicke, M. LEDs make it resilient: Effects on plant growth and defense. Trends Plant Sci. 2021, 26, 496–508. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Yang, G.; Liu, D.; Li, Q.; Zheng, L.; Ma, J. Metabolomics and transcriptomics analysis of vitro growth in pitaya plantlets with different LED Light spectra treatment. Ind. Crops Prod. 2022, 186, 115237. [Google Scholar] [CrossRef]
- Chen, L.-L.; Zhang, K.; Gong, X.-C.; Wang, H.-Y.; Gao, Y.-H.; Wang, X.-Q.; Zeng, Z.-H.; Hu, Y.-G. Effects of different LEDs light spectrum on the growth, leaf anatomy, and chloroplast ultrastructure of potato plantlets in vitro and minituber production after transplanting in the greenhouse. J. Integr. Agric. 2020, 19, 108–119. [Google Scholar] [CrossRef]
- Vivanco-Galván, O.; Jiménez-Gaona, Y.; Castillo, D.; Lucero, H. Blue LED light enhances the growth of Cinchona officinalis L. cultured in vitro. In Organic and Hybrid Sensors and Bioelectronics XV, Proceedings of the SPIE Organic Photonics + Electronics, San Diego, CA, USA, 21–26 August 2022; SPIE: Bellingham, WA, USA, 2010; Volume 12210, p. 12210. [Google Scholar]
- Vialet-Chabrand, S.; Matthews, J.S.A.; Lawson, T. Light, power, action! Interaction of respiratory energy- and blue light-induced stomatal movements. New Phytol. 2021, 231, 2231–2246. [Google Scholar] [CrossRef]
- Matthews, J.S.A.; Vialet-Chabrand, S.; Lawson, T. Role of blue and red light in stomatal dynamic behaviour. J. Exp. Bot. 2019, 71, 2253–2269. [Google Scholar] [CrossRef] [PubMed]
- Morańska, E.; Simlat, M.; Warchoł, M.; Skrzypek, E.; Waligórski, P.; Laurain-Mattar, D.; Spina, R.; Ptak, A. Phenolic Acids and Amaryllidaceae Alkaloids Profiles in Leucojum aestivum L. In Vitro Plants Grown under Different Light Conditions. Molecules 2023, 28, 1525. [Google Scholar] [CrossRef]
- Gupta, R.; Sood, H. Emerging Technologies for the Production of In Vitro Raised Quality Rich Swertia chirayita by Using LED Lights. Sustainability 2023, 15, 1714. [Google Scholar] [CrossRef]
- Lupo, M.; Bashir, M.A.; Silvestri, C.; Brunori, E.; Pica, A.L.; Cristofori, V. LED Lighting Effects on Plant Growth and Quality of Pyrus communis L. Propagated In Vitro. Agronomy 2022, 12, 2531. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.; Kitaya, Y.; Seoka, M.; Kudaka, R.; Yahata, M.; Yamawaki, K.; Shimada, T.; Fujii, H.; Endo, T.; et al. Blue LED light induces regreening in the flavedo of Valencia orange in vitro. Food Chem. 2021, 335, 127621. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, P.H.V.; Oliveira, E.L.; Demetrio, C.A.; Ambrosano, G.B.; Piedade, S.M.S. Effects of different light spectra on the slow-grown in vitro storage and quality of banana plantlets cv. Prata Catarina (AAB). Plant Cell Tissue Organ Cult. (PCTOC) 2022, 150, 479–485. [Google Scholar] [CrossRef]
- Kalaitzoglou, P.; Taylor, C.; Calders, K.; Hogervorst, M.; van Ieperen, W.; Harbinson, J.; de Visser, P.; Nicole, C.C.S.; Marcelis, L.F.M. Unraveling the effects of blue light in an artificial solar background light on growth of tomato plants. Environ. Exp. Bot. 2021, 184, 104377. [Google Scholar] [CrossRef]
- Kaiser, E.; Ouzounis, T.; Giday, H.; Schipper, R.; Heuvelink, E.; Marcelis, L.F.M. Adding Blue to Red Supplemental Light Increases Biomass and Yield of Greenhouse-Grown Tomatoes, but Only to an Optimum. Front. Plant Sci. 2019, 9. [Google Scholar] [CrossRef]
- Izzo, L.G.; Hay Mele, B.; Vitale, L.; Vitale, E.; Arena, C. The role of monochromatic red and blue light in tomato early photomorphogenesis and photosynthetic traits. Environ. Exp. Bot. 2020, 179, 104195. [Google Scholar] [CrossRef]
- Fang, L.; Ma, Z.; Wang, Q.; Nian, H.; Ma, Q.; Huang, Q.; Mu, Y. Plant Growth and Photosynthetic Characteristics of Soybean Seedlings Under Different LED Lighting Quality Conditions. J. Plant Growth Regul. 2021, 40, 668–678. [Google Scholar] [CrossRef]
- Jin, D.Z.; Su, X.F.; Li, Y.F.; Shi, M.M.; Yang, B.B.; Wan, W.C.; Wen, X.; Yang, S.J.; Ding, X.T.; Zou, J. Effect of Red and Blue Light on Cucumber Seedlings Grown in a Plant Factory. Horticulturae 2023, 9, 124. [Google Scholar] [CrossRef]
- Tang, D.F.; Huang, Q.F.; Wei, K.H.; Yang, X.A.; Wei, F.; Miao, J.H. Identification of Differentially Expressed Genes and Pathways Involved in Growth and Development of Mesona chinensis Benth Under Red- and Blue-Light Conditions. Front. Plant Sci. 2021, 12, 761068. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.E.; Kong, Y.; Zheng, Y. Elongation growth mediated by blue light varies with light intensities and plant species: A comparison with red light in arugula and mustard seedlings. Environ. Exp. Bot. 2020, 169, 103898. [Google Scholar] [CrossRef]
- Nair, I.; Mazumdar, P.; Singh, P.; Rengasamy, N.; Harikrishna, J.A. Increasing the Ratio of Blue to Red Light Improves Growth and Phytochemical Content in Hydrocotyle bonariensis. Russ. J. Plant Physiol. 2021, 68, 337–346. [Google Scholar] [CrossRef]
- Di, Q.H.; Li, J.; Du, Y.F.; Wei, M.; Shi, Q.H.; Li, Y.; Yang, F.J. Combination of Red and Blue Lights Improved the Growth and Development of Eggplant (Solanum melongena L.) Seedlings by Regulating Photosynthesis. J. Plant Growth Regul. 2021, 40, 1477–1492. [Google Scholar] [CrossRef]
- Yang, J.; Li, C.; Kong, D.; Guo, F.; Wei, H. Light-Mediated Signaling and Metabolic Changes Coordinate Stomatal Opening and Closure. Front. Plant Sci. 2020, 11, 601478. [Google Scholar] [CrossRef] [PubMed]
- Shafiq, I.; Hussain, S.; Raza, M.A.; Iqbal, N.; Asghar, M.A.; Raza, A.; Fan, Y.-F.; Mumtaz, M.; Shoaib, M.; Ansar, M.; et al. Crop photosynthetic response to light quality and light intensity. J. Integr. Agric. 2021, 20, 4–23. [Google Scholar] [CrossRef]
- Zhang, T.; Shi, Y.; Piao, F.; Sun, Z. Effects of different LED sources on the growth and nitrogen metabolism of lettuce. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 134, 231–240. [Google Scholar] [CrossRef]
- Xiaoying, L.; Mingjuan, Y.; Xiaodong, X.; Abm, K.; Atak, A.; Caihong, Z.; Dawei, L. Effect of light on growth and chlorophyll development in kiwifruit ex vitro and in vitro. Sci. Hortic. 2022, 291, 110599. [Google Scholar] [CrossRef]
- Oh, H.E.; Yoon, A.; Park, Y.G. Red Light Enhances the Antioxidant Properties and Growth of Rubus hongnoensis. Plants 2021, 10, 2589. [Google Scholar] [CrossRef]
- Zhang, S.; Guo, X.; Li, J.; Zhang, Y.; Yang, Y.; Zheng, W.; Xue, X. Effects of light-emitting diode spectral combinations on growth and quality of pea sprouts under long photoperiod. Front. Plant Sci. 2022, 13, 978462. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Guo, X.; Zhang, S.; Zhang, Y.; Chen, L.; Zheng, W.; Xue, X. Effects of light quality on growth, nutritional characteristics, and antioxidant properties of winter wheat seedlings (Triticum aestivum L.). Front. Plant Sci. 2022, 13, 978468. [Google Scholar] [CrossRef]
- Li, Z.; Chen, Q.; Xin, Y.; Mei, Z.; Gao, A.; Liu, W.; Yu, L.; Chen, X.; Chen, Z.; Wang, N. Analyses of the photosynthetic characteristics, chloroplast ultrastructure, and transcriptome of apple (Malus domestica) grown under red and blue lights. BMC Plant Biol. 2021, 21, 483. [Google Scholar] [CrossRef]
- Frede, K.; Schreiner, M.; Zrenner, R.; Graefe, J.; Baldermann, S. Carotenoid biosynthesis of pak choi (Brassica rapa ssp. chinensis) sprouts grown under different light-emitting diodes during the diurnal course. Photochem. Photobiol. Sci. 2018, 17, 1289–1300. [Google Scholar] [CrossRef] [PubMed]
- Frede, K.; Baldermann, S. Accumulation of carotenoids in Brassica rapa ssp. chinensis by a high proportion of blue in the light spectrum. Photochem. Photobiol. Sci. 2022, 21, 1947–1959. [Google Scholar] [CrossRef]
- Gotoh, E.; Suetsugu, N.; Higa, T.; Matsushita, T.; Tsukaya, H.; Wada, M. Palisade cell shape affects the light-induced chloroplast movements and leaf photosynthesis. Sci. Rep. 2018, 8, 1472. [Google Scholar] [CrossRef]
- Chen, L.; Yang, Y.; Jiang, Y.; Zhao, J.; Zang, H.; Wang, X.; Hu, Y.; Xue, X. RNA-Seq Analysis Reveals Differential Responses of Potato (Solanum tuberosum L.) Plantlets Cultured in vitro to Red, Blue, Green, and White Light-emitting Diodes (LEDs). J. Plant Growth Regul. 2019, 38, 1412–1427. [Google Scholar] [CrossRef]
- Kasperbauer, M.J.; Hamilton, J.L. Chloroplast Structure and Starch Grain Accumulation in Leaves That Received Different Red and Far-Red Levels during Development. Plant Physiol. 1984, 74, 967–970. [Google Scholar] [CrossRef]
- Bondada, B.R.; Syvertsen, J.P. Concurrent changes in net CO2 assimilation and chloroplast ultrastructure in nitrogen deficient citrus leaves. Environ. Exp. Bot. 2005, 54, 41–48. [Google Scholar] [CrossRef]
- Pashkovskiy, P.; Kreslavski, V.D.; Ivanov, Y.; Ivanova, A.; Kartashov, A.; Shmarev, A.; Strokina, V.; Kuznetsov, V.V.; Allakhverdiev, S.I. Influence of Light of Different Spectral Compositions on the Growth, Photosynthesis, and Expression of Light-Dependent Genes of Scots Pine Seedlings. Cells 2021, 10, 3284. [Google Scholar] [CrossRef]
- Luo, J.; Abid, M.; Tu, J.; Gao, P.; Wang, Z.; Huang, H. Genome-Wide Identification of the LHC Gene Family in Kiwifruit and Regulatory Role of AcLhcb3.1/3.2 for Chlorophyll a Content. Int. J. Mol. Sci. 2022, 23, 6528. [Google Scholar] [CrossRef]
- Castelletti, S.; Morosinotto, T.; Robert, B.; Caffarri, S.; Bassi, R.; Croce, R. Recombinant Lhca2 and Lhca3 Subunits of the Photosystem I Antenna System. Biochemistry 2003, 42, 4226–4234. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-H.; Li, X.-P.; Razeghifard, R.; Anderson, J.M.; Niyogi, K.K.; Pogson, B.J.; Chow, W.S. The multiple roles of light-harvesting chlorophyll a/b-protein complexes define structure and optimize function of Arabidopsis chloroplasts: A study using two chlorophyll b-less mutants. Biochim. Biophys. Acta (BBA)-Bioenerg. 2009, 1787, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Xiao, Y.H.; Zhang, L.; Zhao, Y.G. Analysis of Lhc family genes reveals development regulation and diurnal fluctuation expression patterns in Cyperus esculentus, a Cyperaceae plant. Planta 2023, 257, 59. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Q.; Li, R.; Fernie, A.R.; Che, Y.; Ding, Z.; Yao, Y.; Liu, J.; Wang, Y.; Hu, X.; Guo, J. Integrated Analysis of Morphological, Physiological, Anatomical and Molecular Responses of Cassava Seedlings to Different Light Qualities. Int. J. Mol. Sci. 2023, 24, 14224. https://doi.org/10.3390/ijms241814224
Zhou Q, Li R, Fernie AR, Che Y, Ding Z, Yao Y, Liu J, Wang Y, Hu X, Guo J. Integrated Analysis of Morphological, Physiological, Anatomical and Molecular Responses of Cassava Seedlings to Different Light Qualities. International Journal of Molecular Sciences. 2023; 24(18):14224. https://doi.org/10.3390/ijms241814224
Chicago/Turabian StyleZhou, Qin, Ruimei Li, Alisdair R. Fernie, Yannian Che, Zhongping Ding, Yuan Yao, Jiao Liu, Yajie Wang, Xinwen Hu, and Jianchun Guo. 2023. "Integrated Analysis of Morphological, Physiological, Anatomical and Molecular Responses of Cassava Seedlings to Different Light Qualities" International Journal of Molecular Sciences 24, no. 18: 14224. https://doi.org/10.3390/ijms241814224