
Recovering What Matters: High Protein Recovery after Endotoxin Removal from LPS-Contaminated Formulations Using Novel Anti-Lipid A Antibody Microparticle Conjugates
 , , , , , and
, , , , , and 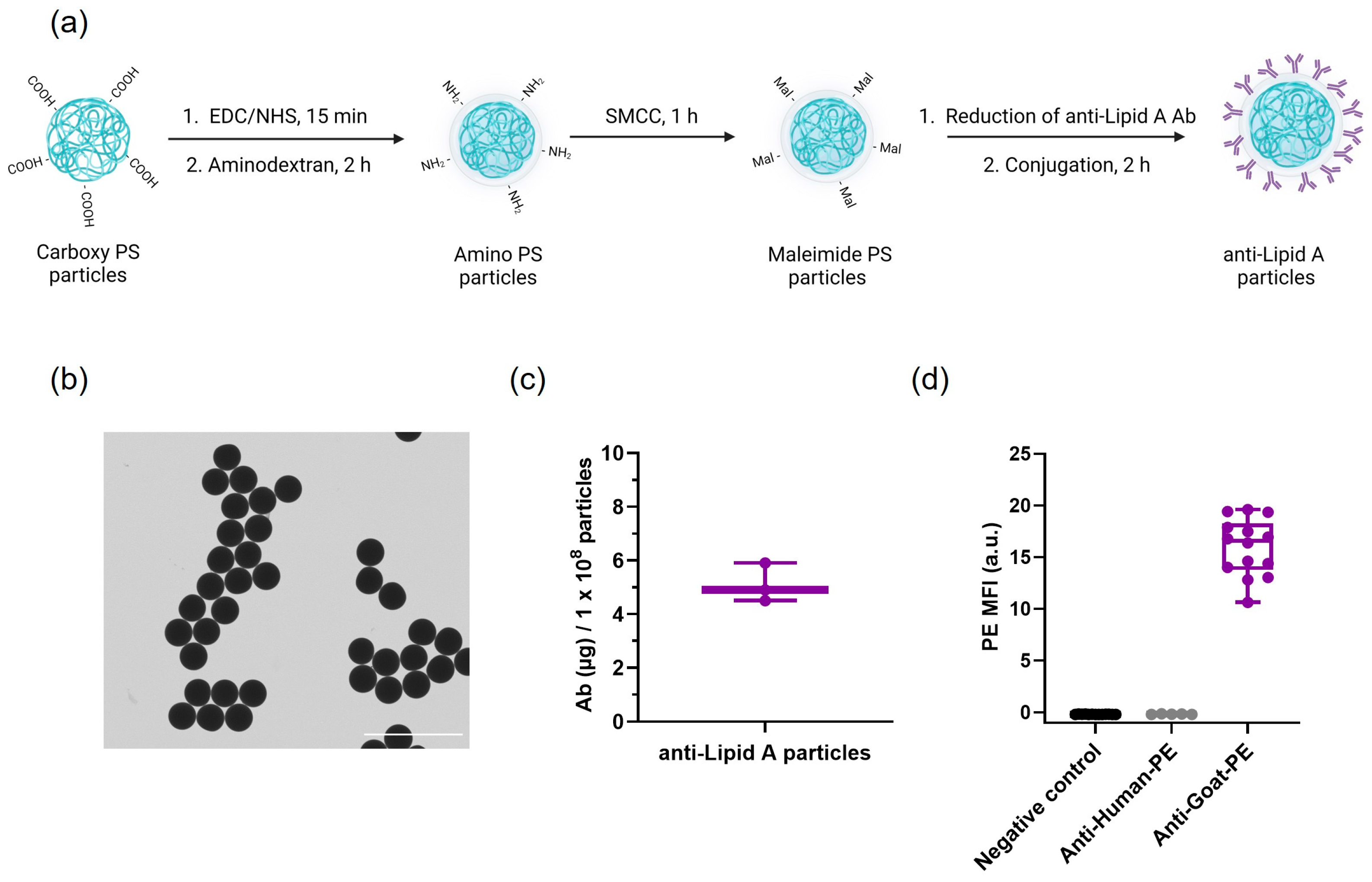{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Anti-Lipid A Particles: Conjugation and Characterization
2.2. Anti-Lipid A Particles’ Functionality and Stability
2.2.1. LPS Removal Capacity of Anti-Lipid A Particles
2.2.2. Longitudinal Stability
2.3. LPS Removal and Protein Recovery
2.3.1. LPS Removal and Protein Recovery in 1 mg/mL BSA in Water
2.3.2. LPS Removal and Protein Recovery in 1 mg/mL Insulin in HEPES 25 mM
2.3.3. LPS Removal and Protein Recovery in Naturally LPS-Contaminated BPE in DPBS
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Anti-Lipid A Particles’ Conjugation
3.2.2. Anti-Lipid A Particles’ Characterization
3.2.3. LPS Removal Capacity of Anti-Lipid A Particles
3.2.4. Longitudinal Stability
3.2.5. LPS Removal and Protein Recovery in 1 mg/mL BSA in Water
3.2.6. LPS Removal and Protein Recovery in 1 mg/mL Insulin Solution in HEPES 25 mM
3.2.7. LPS Removal and Protein Recovery in Naturally LPS-Contaminated BPE in DPBS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Silhavy, T.J.; Kahne, D.; Walker, S. The Bacterial Cell Envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000414. [Google Scholar] [CrossRef]
- Stromberg, L.R.; Mendez, H.M.; Mukundan, H. Detection Methods for Lipopolysaccharides: Past and Present. In Escherichia coli—Recent Advances on Physiology, Pathogenesis and Biotechnological Applications; Samie, A., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Sampath, V. Bacterial endotoxin-lipopolysaccharide; structure, function and its role in immunity in vertebrates and invertebrates. Agric. Nat. Resour. 2018, 52, 115–120. [Google Scholar] [CrossRef]
- Williams, K.L. Historical and Emerging Themes. In Endotoxins: Pyrogens, LAL Testing and Depyrogenation; Williams, K.L., Ed.; Informa Healthcare Inc.: Richmond, VA, USA, 2007. [Google Scholar]
- Fux, A.C.; Melo, C.C.; Michelini, S.; Swartzwelter, B.J.; Neusch, A.; Italiani, P.; Himly, M. Heterogeneity of Lipopolysaccharide as Source of Variability in Bioassays and LPS-Binding Proteins as Remedy. Int. J. Mol. Sci. 2023, 24, 8395. [Google Scholar] [CrossRef] [PubMed]
- Schneier, M.; Razdan, S.; Miller, A.M.; Briceno, M.E.; Barua, S. Current technologies to endotoxin detection and removal for biopharmaceutical purification. Biotechnol. Bioeng. 2020, 117, 2588–2609. [Google Scholar] [CrossRef]
- U.S. Food & Drug Administration. Bacterial Endotoxins/Pyrogens. Available online: https://www.fda.gov/inspections-compliance-enforcement-and-criminal-investigations/inspection-technical-guides/bacterial-endotoxinspyrogens (accessed on 17 December 2020).
- Dawson, M. Endotoxin Limits for Parenteral Drug Products; BET White Paper; Associates of Cape Cod, Inc.: East Falmouth, MA, USA, 2017; Volume 1. [Google Scholar]
- Sandle, T. A Comparative Study of Different Methods for Endotoxin Destruction. Am. Pharm. Rev. 2013, 16, 15–17. [Google Scholar]
- Bononi, I.; Balatti, V.; Gaeta, S.; Tognon, M. Gram-Negative Bacterial Lipopolysaccharide Retention by a Positively Charged New-Generation Filter. Appl. Environ. Microbiol. 2008, 74, 6470–6472. [Google Scholar] [CrossRef]
- Sandle, T. A Practical Approach to Depyrogenation Studies Using Bacterial Endotoxin. J. GXP Compliance 2011, 15, 90–96. [Google Scholar]
- de Oliveira Magalhães, P.O.; Lopes, A.M.; Mazzola, P.G.; Rangel-Yagui, C.; Penna, T.C.V.; Pessoa Júnior, A. Methods of endotoxin removal from biological preparations: A review. J. Pharm. Pharm. Sci. 2007, 10, 388–404. [Google Scholar]
- Razdan, S.; Wang, J.-C.; Barua, S. PolyBall: A new adsorbent for the efficient removal of endotoxin from biopharmaceuticals. Sci. Rep. 2019, 9, 8867. [Google Scholar] [CrossRef]
- Anspach, F.B.; Petsch, D. Membrane adsorbers for selective endotoxin removal from protein solutions. Process. Biochem. 2000, 35, 1005–1012. [Google Scholar] [CrossRef]
- Vagenende, V.; Ching, T.-J.; Chua, R.-J.; Thirumoorthi, N.; Gagnon, P. Amide-Mediated Hydrogen Bonding at Organic Crystal/Water Interfaces Enables Selective Endotoxin Binding with Picomolar Affinity. ACS Appl. Mater. Interfaces 2013, 5, 4472–4478. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shang, G.; You, M.; Peng, S.; Wang, Z.; Wu, H.; Chen, G.-Q. Endotoxin Removing Method Based on Lipopolysaccharide Binding Protein and Polyhydroxyalkanoate Binding Protein PhaP. Biomacromolecules 2011, 12, 602–608. [Google Scholar] [CrossRef]
- Chen, J.; Shi, Z.; Yang, X.; Zhang, X.; Wang, D.; Qian, S.; Sun, W.; Wang, C.; Li, Q.; Wang, Z.; et al. Broad-Spectrum Clearance of Lipopolysaccharides from Blood Based on a Hemocompatible Dihistidine Polymer. ACS Appl. Mater. Interfaces 2023, 15, 32251–32261. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, X.; Yang, X.; Zhang, X.; Ma, F.; Gan, H.; Chen, J.; Wang, D.; Sun, W.; Wang, J.; et al. Specific Clearance of Lipopolysaccharide from Blood Based on Peptide Bottlebrush Polymer for Sepsis Therapy. Adv. Mater. 2023, 35, 2302560. [Google Scholar] [CrossRef]
- Ribeiro, L.N.d.M.; Couto, V.M.; Fraceto, L.F.; de Paula, E. Use of nanoparticle concentration as a tool to understand the structural properties of colloids. Sci. Rep. 2018, 8, 982. [Google Scholar] [CrossRef]
- Filipe, V.; Hawe, A.; Jiskoot, W. Critical Evaluation of Nanoparticle Tracking Analysis (NTA) by NanoSight for the Measurement of Nanoparticles and Protein Aggregates. Pharm. Res. 2010, 27, 796–810. [Google Scholar] [CrossRef]
- Comfort, N.; Cai, K.; Bloomquist, T.R.; Strait, M.D.; Ferrante, A.W.J.; Baccarelli, A.A. Nanoparticle Tracking Analysis for the Quantification and Size Determination of Extracellular Vesicles. JoVE J. Vis. Exp. 2021, 28, e62447. [Google Scholar]
- Williams, K.L. Endotoxin Relevance and Control Overview. In Endotoxins: Pyrogens, LAL Testing and Depyrogenation; Informa Healthcare Inc.: Richmond, VA, USA, 2007. [Google Scholar]
- Kundu, N.; Banik, D.; Sarkar, N. Self-Assembly of Amphiphiles into Vesicles and Fibrils: Investigation of Structure and Dynamics Using Spectroscopy and Microscopy Techniques. Langmuir 2018, 34, 11637–11654. [Google Scholar] [CrossRef]
- Parikh, S.J.; Chorover, J. Infrared spectroscopy studies of cation effects on lipopolysaccharides in aqueous solution. Colloids Surf. B Biointerfaces 2007, 55, 241–250. [Google Scholar] [CrossRef]
- Santos, N.C.; Silva, A.C.; Castanho, M.A.R.B.; Martins-Silva, J.; Saldanha, C. Evaluation of Lipopolysaccharide Aggregation by Light Scattering Spectroscopy. ChemBioChem 2003, 4, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Miltenyi Biotec B.V. & Co. KG. Endotoxin Removal Beads. Available online: https://static.miltenyibiotec.com/asset/150655405641/document_505pfa7o0l78p0kmisccvm4f29?content-disposition=inline (accessed on 26 January 2022).
- Ionova, Y.; Wilson, L. Biologic excipients: Importance of clinical awareness of inactive ingredients. PLoS ONE 2020, 15, e0235076. [Google Scholar] [CrossRef] [PubMed]
- ThermoFisher Scientific. Bovine Serum Albumin (BSA Protein). Available online: https://www.thermofisher.com/be/en/home/life-science/cell-culture/mammalian-cell-culture/media-supplements/bovine-serum-albumin.html (accessed on 17 May 2023).
- Esparza, G.A.; Teghanemt, A.; Zhang, D.; Gioannini, T.L.; Weiss, J.P. Endotoxin·albumin complexes transfer endotoxin monomers to MD-2 resulting in activation of TLR4. Innate Immun. 2012, 18, 478–491. [Google Scholar] [CrossRef]
- Landgraf, W.; Sandow, J. Recombinant Human Insulins—Clinical Efficacy and Safety in Diabetes Therapy. Eur. Endocrinol. 2016, 12, 12–17. [Google Scholar] [CrossRef]
- Gupta, V.; Sengupta, M.; Prakash, J.; Tripathy, B.C. Production of Recombinant Pharmaceutical Proteins. Basic Appl. Asp. Biotechnol. 2016, 77–101. [Google Scholar] [CrossRef]
- Donner, T.; Sarkar, S. Insulin—Pharmacology, Therapeutic Regimens, and Principles of Intensive Insulin Therapy; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Biedermann, T.; Winther, L.; Till, S.J.; Panzner, P.; Knulst, A.; Valovirta, E. Birch pollen allergy in Europe. Allergy 2019, 74, 1237–1248. [Google Scholar] [CrossRef]
- Pablos, I.; Wildner, S.; Asam, C.; Wallner, M.; Gadermaier, G. Pollen Allergens for Molecular Diagnosis. Curr. Allergy Asthma Rep. 2016, 16, 31. [Google Scholar] [CrossRef] [PubMed]
- Pointner, L.; Bethanis, A.; Thaler, M.; Traidl-Hoffmann, C.; Gilles, S.; Ferreira, F.; Aglas, L. Initiating Pollen Sensitization—Complex Source, Complex Mechanisms. Clin. Transl. Allergy 2020, 10, 36. [Google Scholar] [CrossRef]
- Manirajan, B.A.; Hinrichs, A.-K.; Ratering, S.; Rusch, V.; Schwiertz, A.; Geissler-Plaum, R.; Eichner, G.; Cardinale, M.; Kuntz, S.; Schnell, S. Bacterial Species Associated with Highly Allergenic Plant Pollen Yield a High Level of Endotoxins and Induce Chemokine and Cytokine Release from Human A549 Cells. Inflammation 2022, 45, 2186–2201. [Google Scholar] [CrossRef]
- Di Lorenzo, F.; Duda, K.A.; Lanzetta, R.; Silipo, A.; De Castro, C.; Molinaro, A. A Journey from Structure to Function of Bacterial Lipopolysaccharides. Chem. Rev. 2021, 122, 15767–15821. [Google Scholar] [CrossRef] [PubMed]
- Jokerst, J.V.; Floriano, P.N.; Christodoulides, N.; Simmons, G.W.; McDevitt, J.T. Integration of semiconductor quantum dots into nano-bio-chip systems for enumeration of CD4+ T cell counts at the point-of-need. Lab A Chip 2008, 8, 2079–2090. [Google Scholar] [CrossRef]
- Tiwari, D.K.; Tanaka, S.-I.; Inouye, Y.; Yoshizawa, K.; Watanabe, T.M.; Jin, T. Synthesis and Characterization of Anti-HER2 Antibody Conjugated CdSe/CdZnS Quantum Dots for Fluorescence Imaging of Breast Cancer Cells. Sensors 2009, 9, 9332–9354. [Google Scholar] [CrossRef]
- Woythe, L.; Tholen, M.M.E.; Rosier, B.J.H.M.; Albertazzi, L. Single-Particle Functionality Imaging of Antibody-Conjugated Nanoparticles in Complex Media. ACS Appl. Bio Mater. 2023, 6, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Bio-Rad Doublet Discrimination . Available online: https://www.bio-rad-antibodies.com/flow-cytometry-doublet-discrimination.html (accessed on 3 July 2023).
- Petsch, D.; Deckwer, W.-D.; Anspach, F. Proteinase K Digestion of Proteins Improves Detection of Bacterial Endotoxins by the Limulus Amebocyte Lysate Assay: Application for Endotoxin Removal from Cationic Proteins. Anal. Biochem. 1998, 259, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.; Aglas, L.; Punz, B.; Dang, H.-H.; Christ, C.; Pointner, L.; Wenger, M.; Hofstaetter, N.; Hofer, S.; Geppert, M.; et al. Mechanistic insights into silica nanoparticle–allergen interactions on antigen presenting cell function in the context of allergic reactions. Nanoscale 2023, 15, 2262–2275. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casonato Melo, C.; Fux, A.C.; Himly, M.; Bastús, N.G.; Schlahsa, L.; Siewert, C.; Puntes, V.; Duschl, A.; Gessner, I.; Fauerbach, J.A. Recovering What Matters: High Protein Recovery after Endotoxin Removal from LPS-Contaminated Formulations Using Novel Anti-Lipid A Antibody Microparticle Conjugates. Int. J. Mol. Sci. 2023, 24, 13971. https://doi.org/10.3390/ijms241813971
Casonato Melo C, Fux AC, Himly M, Bastús NG, Schlahsa L, Siewert C, Puntes V, Duschl A, Gessner I, Fauerbach JA. Recovering What Matters: High Protein Recovery after Endotoxin Removal from LPS-Contaminated Formulations Using Novel Anti-Lipid A Antibody Microparticle Conjugates. International Journal of Molecular Sciences. 2023; 24(18):13971. https://doi.org/10.3390/ijms241813971
Chicago/Turabian StyleCasonato Melo, Cristiane, Alexandra C. Fux, Martin Himly, Neus G. Bastús, Laura Schlahsa, Christiane Siewert, Victor Puntes, Albert Duschl, Isabel Gessner, and Jonathan A. Fauerbach. 2023. "Recovering What Matters: High Protein Recovery after Endotoxin Removal from LPS-Contaminated Formulations Using Novel Anti-Lipid A Antibody Microparticle Conjugates" International Journal of Molecular Sciences 24, no. 18: 13971. https://doi.org/10.3390/ijms241813971