
Easy and Versatile Technique for the Preparation of Stable and Active Lipase-Based CLEA-like Copolymers by Using Two Homofunctional Cross-Linking Agents: Application to the Preparation of Enantiopure Ibuprofen


Abstract
:1. Introduction
2. Results
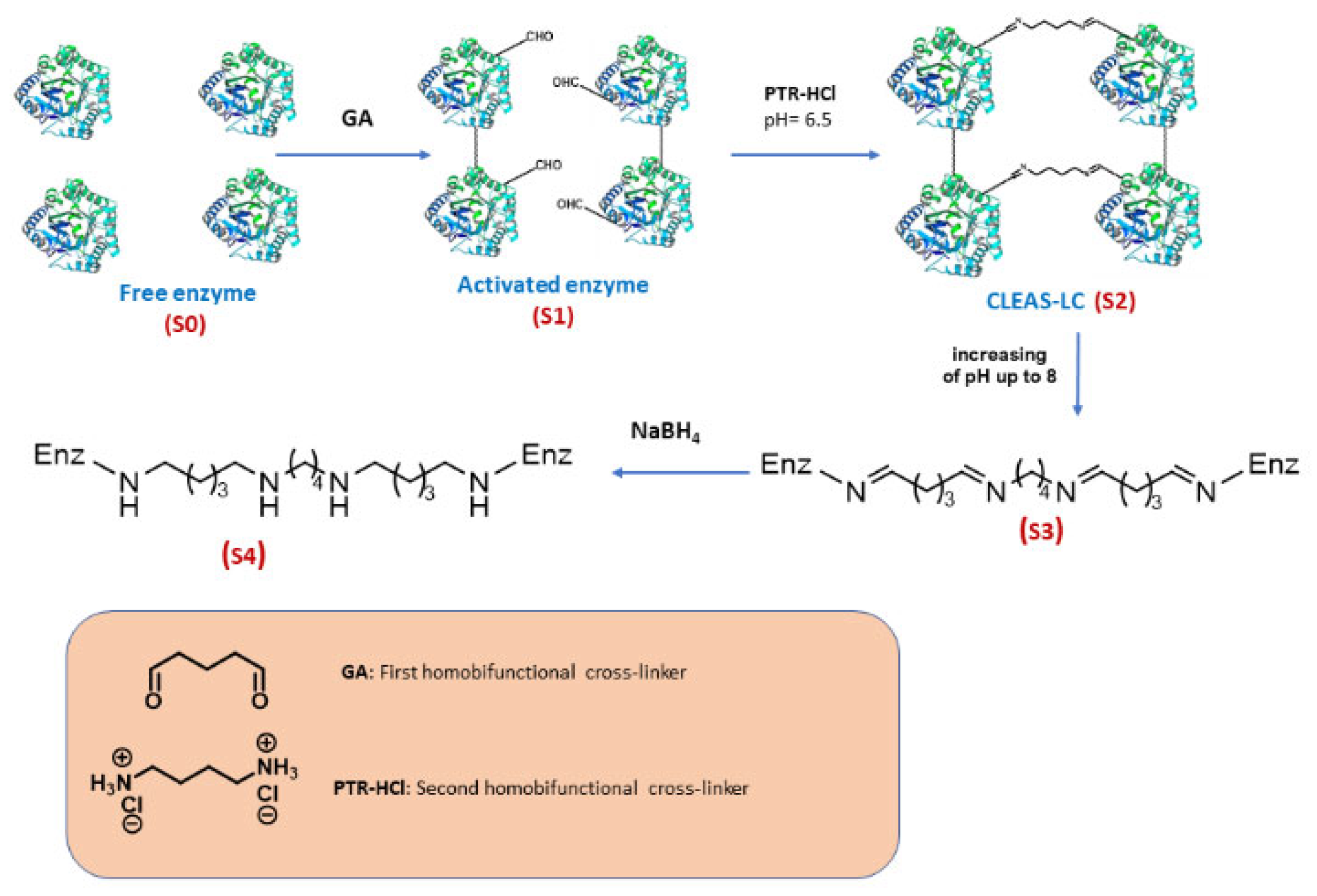
2.1. Preliminary Adjustment of Enzyme and Cross-Linkers Amounts for Further CLEA-LC Preparation
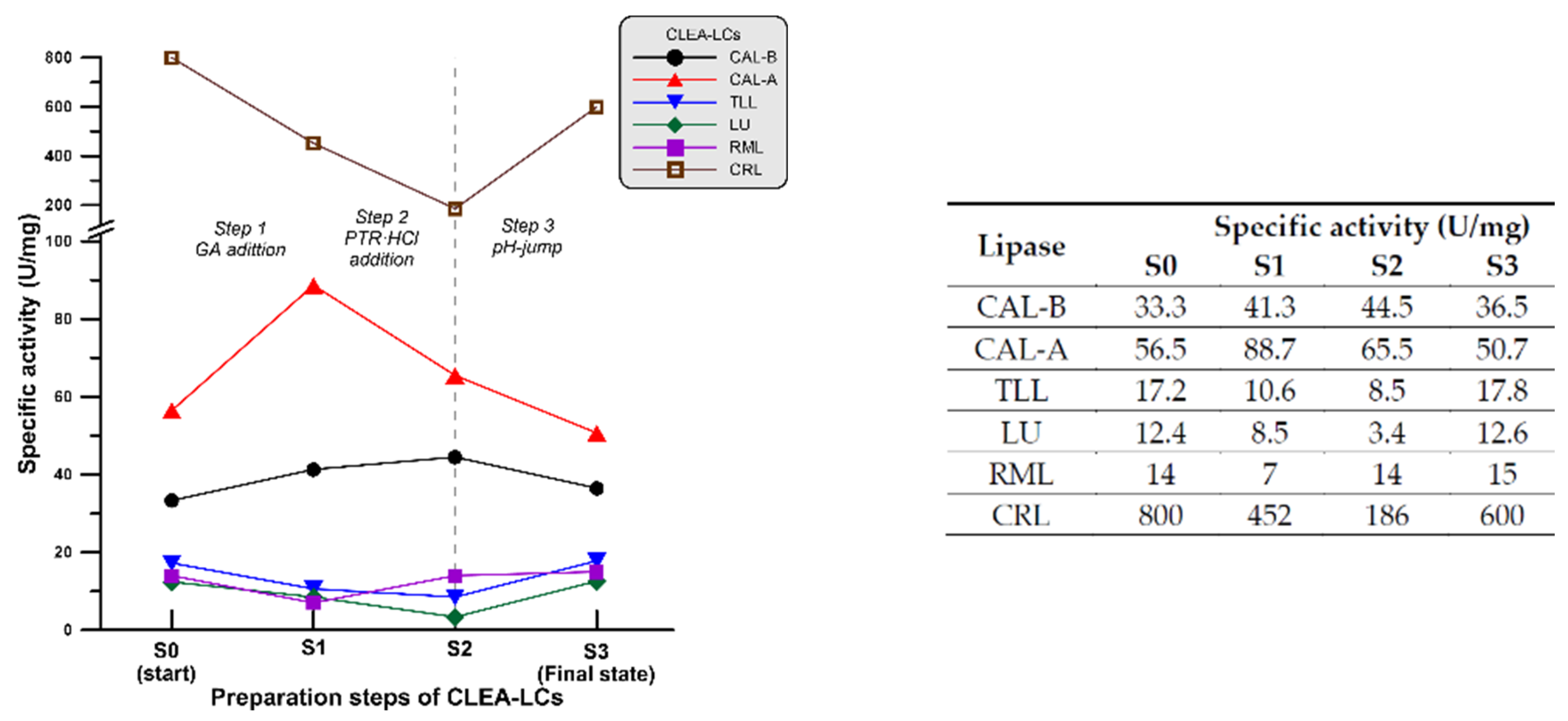
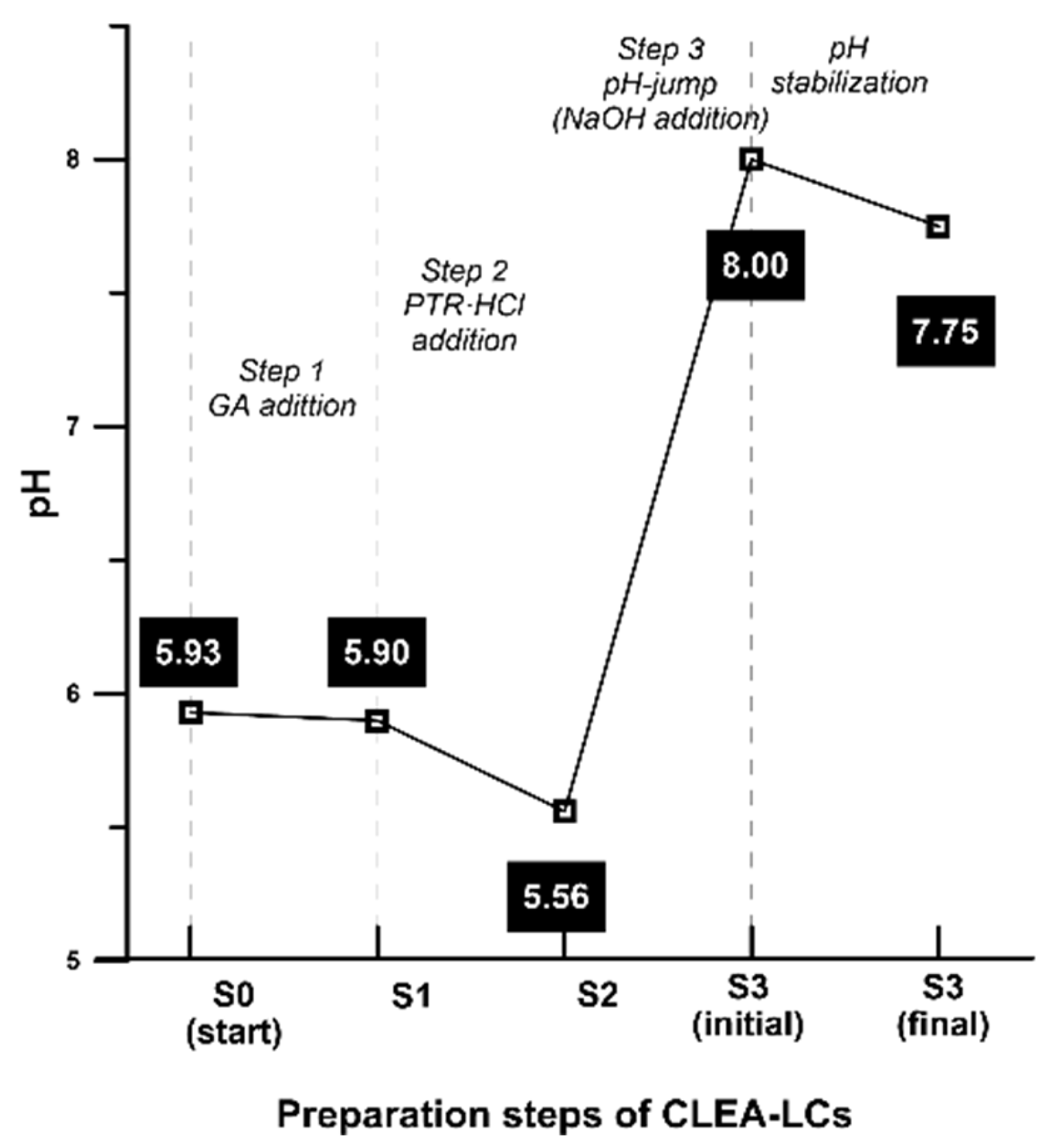
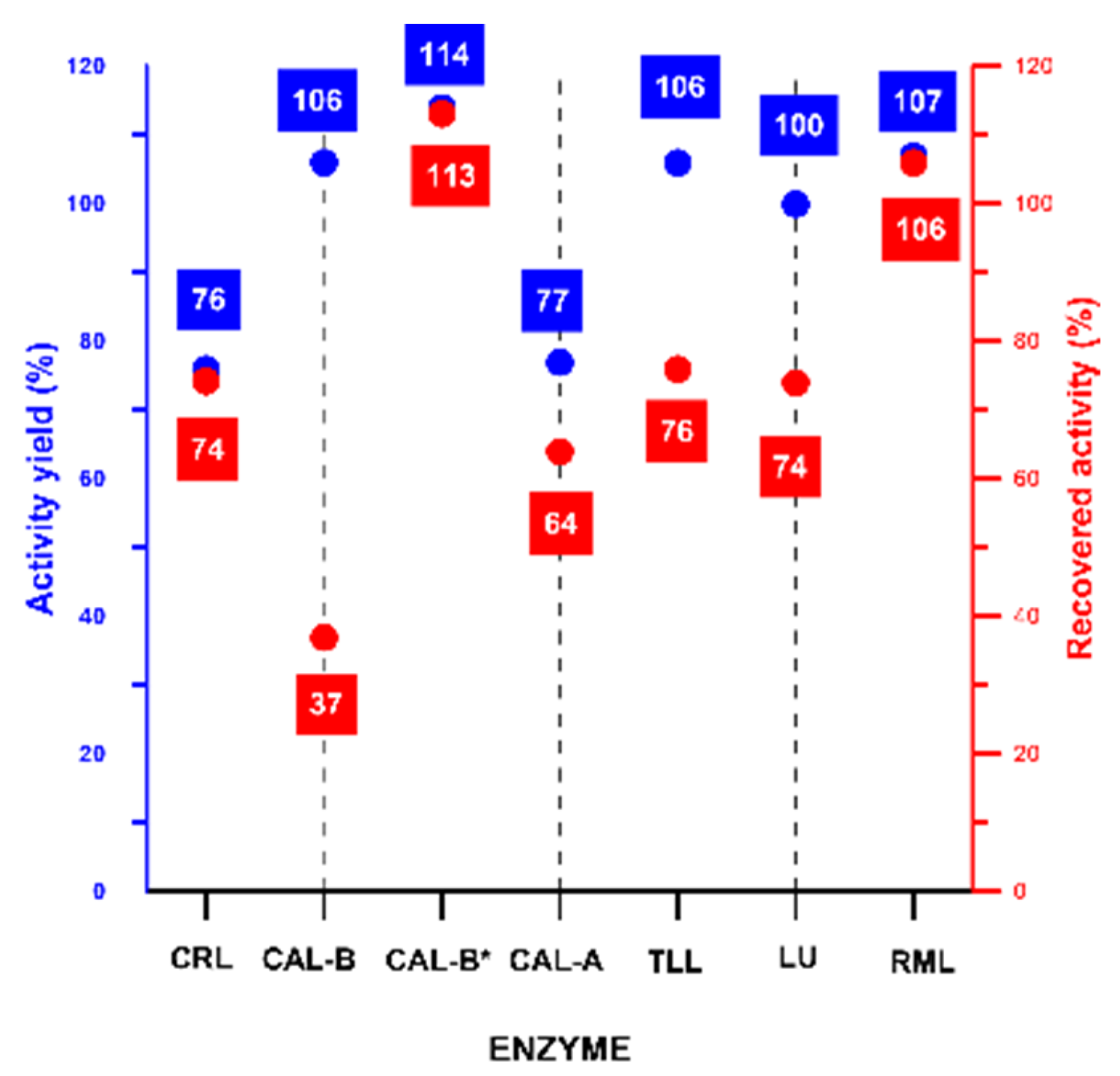
2.2. In-Depth Study of CLEA-like Copolymerization Procedure as a Versatile Technique
2.3. Characterization of CLEA-LC Using FTIR Spectroscopy and SEM
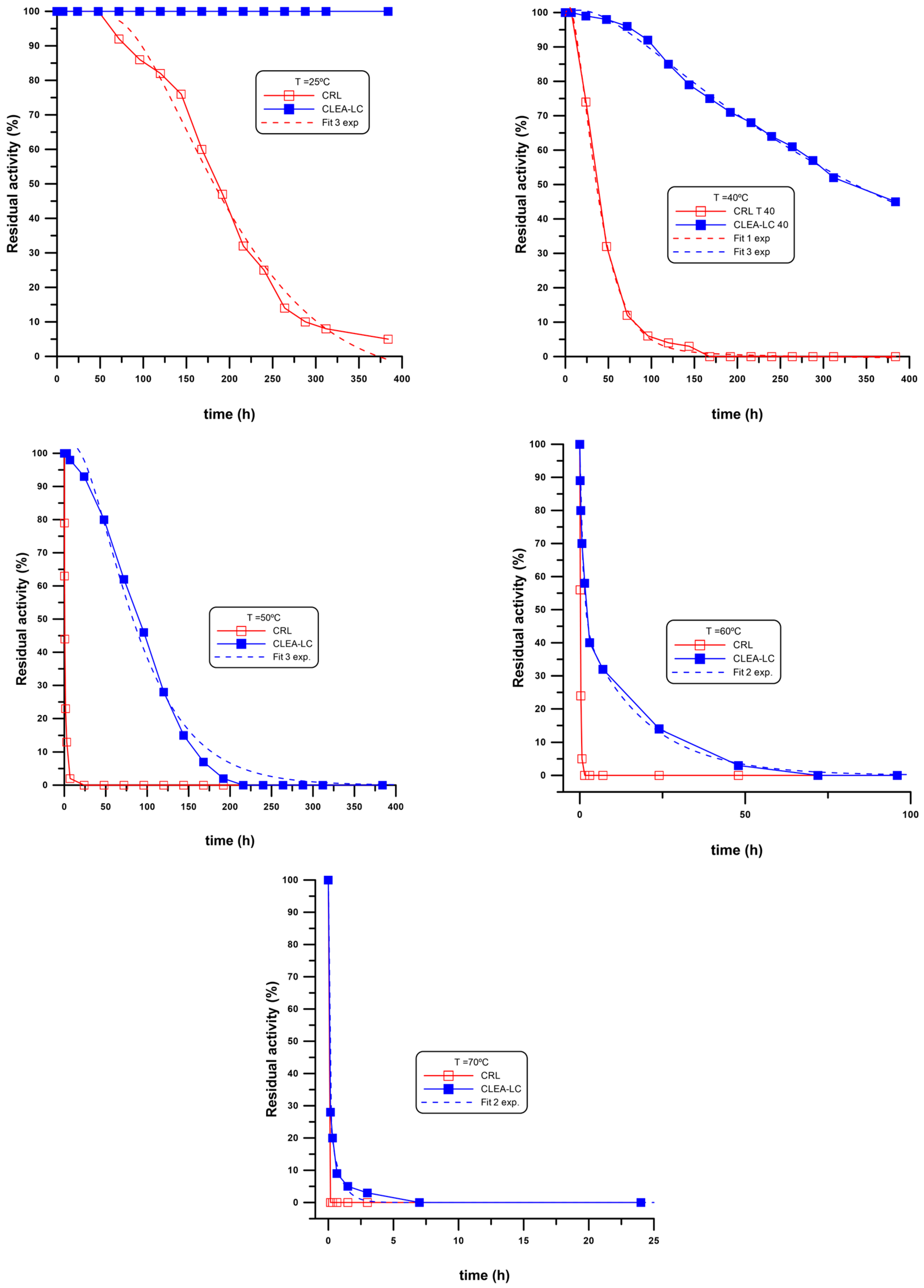
2.4. Thermal Stability of CRL CLEA-LC
2.5. Long Term Storage Stability at 4 °C
2.6. Application of CLEA-LCs in the Kinetic Resolution of Rac-Ibuprofen Ethyl ester
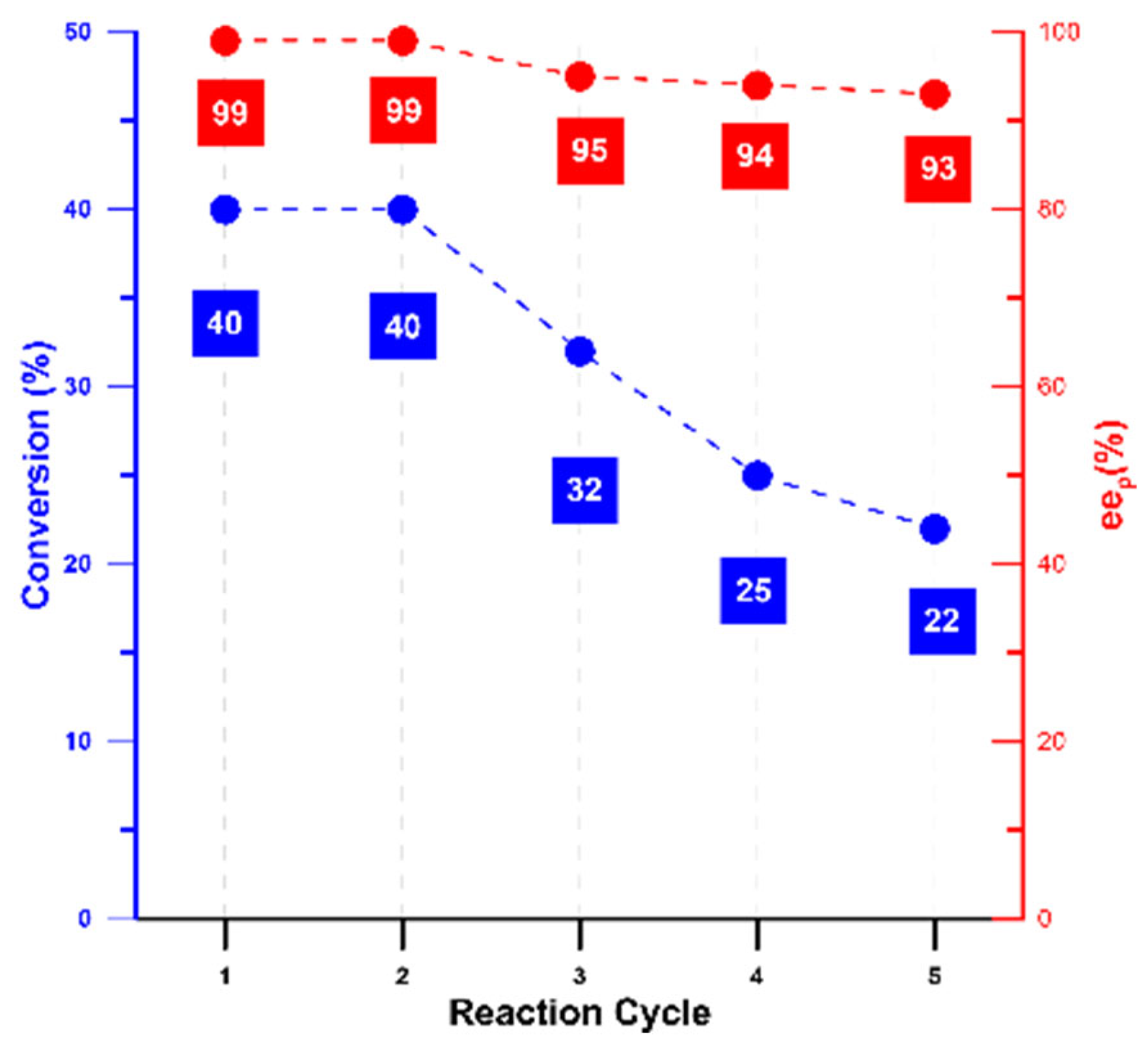
2.7. Reuse of CRL CLEA-LC for the Kinetic Resolution of Rac-Ibuprofen Ethyl Ester
3. Discussion
3.1. Preparation of CLEA-LCs
3.2. Thermal Stability of CRL CLEA-LCs
3.3. Application and Reuse of CLEA-LC
4. Materials and Methods
4.1. Materials
4.1.1. Enzymes
4.1.2. Chemicals
4.2. Methods
4.2.1. Protein Quantification
4.2.2. Preparation of Putrescine HCl pH 6.5 Aqueous Suspension (PTR-HCl)
4.2.3. Enzymatic Activity Assays
4.2.4. Preliminary Preparation of CLEA-LC Using CAL-B As Model Enzyme
4.2.5. Preparation of CLEA-LC Using Six Enzymes
4.2.6. SDS Page Electrophoresis
4.2.7. FT-Infrared of CRL CLEA-CL and Free CRL
4.2.8. Scanning Electron Microscopy Analysis
4.2.9. Thermal Stability CRL CLEA-LC Compared with Free CRL
4.2.10. Preparation of Rac-Ibuprofen Ethyl Ester
4.2.11. Kinetic Resolution of rac-Ibuprofen Ethyl Ester CLEA-LC and Free Enzymes: Comparative Study
4.2.12. Reuse of CRL CLEA-LC
4.2.13. Long-Term Storage of CLEA-LC
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alcántara, A.R.; de María, P.D.; Littlechild, J.; Schürmann, M.; Sheldon, R.A.; Wohlgemuth, R. Biocatalysis as Key to Sustainable Industrial Chemistry. ChemSusChem 2022, 15, e202102709. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A.; Brady, D. Green Chemistry, Biocatalysis, and the Chemical Industry of the Future. ChemSusChem 2022, 15, e202102628. [Google Scholar] [CrossRef]
- Woodley, J.M. Biocatalysis for future sustainable manufacturing. Biochemist 2022, 44, 6–8. [Google Scholar] [CrossRef]
- Zetzsche, L.E.; Chakrabarty, S.; Narayan, A.R.H. The Transformative Power of Biocatalysis in Convergent Synthesis. J. Am. Chem. Soc. 2022, 144, 5214–5225. [Google Scholar] [CrossRef]
- France, S.P.; Lewis, R.D.; Martinez, C.A. The Evolving Nature of Biocatalysis in Pharmaceutical Research and Development. JACS Au 2023, 3, 715–735. [Google Scholar] [CrossRef] [PubMed]
- Holtmann, D.; Hollmann, F.; Bouchaut, B. Contribution of Enzyme Catalysis to the Achievement of the United Nations Sustainable Development Goals. Molecules 2023, 28, 4125. [Google Scholar] [CrossRef]
- Bottcher, D.; Bornscheuer, U.T. Protein engineering of microbial enzymes. Curr. Opin. Microbiol. 2010, 13, 274–282. [Google Scholar] [CrossRef]
- Porter, J.L.; Rusli, R.A.; Ollis, D.L. Directed Evolution of Enzymes for Industrial Biocatalysis. ChemBioChem 2016, 17, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Begley, T.P.; Tsai, M.-D. Biocatalysis and biotransformation Enzymology in the genomics era. Curr. Opin. Chem. Biol. 2003, 7, 228–229. [Google Scholar] [CrossRef]
- Wu, H.; Chen, Q.M.; Zhang, W.L.; Mu, W.M. Overview of strategies for developing high thermostability industrial enzymes: Discovery, mechanism, modification and challenges. Crit. Rev. Food Sci. Nutr. 2023, 63, 2057–2073. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Vivek, N.; Kumar, V.; Chandel, N.; Thakur, M.; Kumar, D.; Yang, Y.H.; Pugazendhi, A.; Kumar, G. Molecular biology interventions for activity improvement and production of industrial enzymes. Bioresour. Technol. Rep. 2021, 324, 124596. [Google Scholar] [CrossRef]
- Sheldon, R.A. Cross-linked enzyme aggregates (CLEA®s): Stable and recyclable biocatalysts. Biochem. Soc.Trans. 2007, 35, 1583–1587. [Google Scholar] [CrossRef]
- Jung, S.; Park, S. Improving the expression yield of Candida antarctica lipase B in Escherichia coli by mutagenesis. Biotechnol. Lett. 2008, 30, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Boudrant, J.; Woodley, J.M.; Fernandez-Lafuente, R. Parameters necessary to define an immobilized enzyme preparation. Process Biochem. 2020, 90, 66–80. [Google Scholar] [CrossRef]
- Bolivar, J.M.; Woodley, J.M.; Fernandez-Lafuente, R. Is enzyme immobilization a mature discipline? Some critical considerations to capitalize on the benefits of immobilization. Chem. Soc. Rev. 2022, 51, 6251–6290. [Google Scholar] [CrossRef]
- Da Rocha, T.N.; Carballares, D.; Guimarães, J.R.; Rocha-Martin, J.; Tardioli, P.W.; Gonçalves, L.R.B.; Fernandez-Lafuente, R. Determination of immobilized lipase stability depends on the substrate and activity determination condition: Stress inactivations and optimal temperature as biocatalysts stability indicators. Sustain. Chem. Pharm. 2022, 29, 100823. [Google Scholar] [CrossRef]
- Rodríguez-Núñez, K.; Bernal, C.; Bolívar, J.M. Chapter 1—The enzyme, the support, and the immobilization strategy: The key findings to a desirable biocatalyst. In Biocatalyst Immobilization; Ferreira, M.L., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 1–16. [Google Scholar]
- Wilson, L.; Illanes, A.; Romero, O.; Ottone, C. Chapter 17—Future perspectives in enzyme immobilization. In Biocatalyst Immobilization; Ferreira, M.L., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 403–426. [Google Scholar]
- Tufvesson, P.; Törnvall, U.; Carvalho, J.; Karlsson, A.J.; Hatti-Kaul, R. Towards a cost-effective immobilized lipase for the synthesis of specialty chemicals. J. Mol. Catal. B Enzym. 2011, 68, 200–205. [Google Scholar] [CrossRef]
- Remonatto, D.; Miotti, R.H.; Monti, R.; Bassan, J.C.; de Paula, A.V. Applications of immobilized lipases in enzymatic reactors: A review. Process Biochem. 2022, 114, 1–20. [Google Scholar] [CrossRef]
- Guisan, J.M.; Fernandez-Lorente, G.; Rocha-Martin, J.; Moreno-Gamero, D. Enzyme immobilization strategies for the design of robust and efficient biocatalysts. Curr. Opin. Green Sustain. Chem. 2022, 35, 100593. [Google Scholar] [CrossRef]
- Goncalves, D.; Silva, A.G.; Guidini, C.Z. Lipases: Sources, immobilization methods, and industrial applications. Appl. Microbiol. Biotechnol. 2019, 103, 7399–7423. [Google Scholar] [CrossRef]
- Enespa; Chandra, P.; Singh, D.P. Sources, purification, immobilization and industrial applications of microbial lipases: An overview. Crit. Rev. Food Sci. Nutr. 2022, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Hanefeld, U.; Gardossi, L.; Magner, E. Understanding enzyme immobilisation. Chem. Soc. Rev. 2009, 38, 453–468. [Google Scholar] [CrossRef] [PubMed]
- Secundo, F. Conformational changes of enzymes upon immobilisation. Chem. Soc. Rev. 2013, 42, 6250–6261. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lorente, G.; Lopez-Gallego, F.; Bolivar, J.M.; Rocha-Martin, J.; Moreno-Perez, S.; Guisan, J.M. Immobilization of Proteins on Highly Activated Glyoxyl Supports: Dramatic Increase of the Enzyme Stability via Multipoint Immobilization on Pre-existing Carriers. Curr. Org. Chem. 2015, 19, 1719–1731. [Google Scholar] [CrossRef]
- Weltz, J.S.; Kienle, D.F.; Schwartz, D.K.; Kaar, J.L. Reduced Enzyme Dynamics upon Multipoint Covalent Immobilization Leads to Stability-Activity Trade-off. J. Am. Chem. Soc. 2020, 142, 3463–3471. [Google Scholar] [CrossRef]
- Bilal, M.; Asgher, M.; Cheng, H.; Yan, Y.; Iqbal, H.M.N. Multi-point enzyme immobilization, surface chemistry, and novel platforms: A paradigm shift in biocatalyst design. Crit. Rev. Biotechnol. 2019, 39, 202–219. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Sanchez, D.; Lopez-Iglesias, M.; Gotor-Fernandez, V. Hydrolases in Organic Chemistry. Recent Achievements in the Synthesis of Pharmaceuticals. Curr.Org. Chem. 2016, 20, 1186–1203. [Google Scholar] [CrossRef]
- Kazlauskas, R.J.; Bornscheuer, U.T.; Bornscheuer, U.T.; Kazlauskas, R.J. Hydrolases in Organic Synthesis Regio- and Stereoselective Biotransformations Introduction; John Wiley & Sons: Hoboken, NJ, USA, 2006; pp. 1–3. [Google Scholar]
- Hall, M. Enzymatic strategies for asymmetric synthesis. RSC Chem. Biol. 2021, 2, 958–989. [Google Scholar] [CrossRef]
- Gutarra, M.L.E.; Miranda, L.S.M.; de Souza, R.O.M.A. Chapter 4—Enzyme Immobilization for Organic Synthesis. In Organic Synthesis Using Biocatalysis; Goswami, A., Stewart, J.D., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 99–126. [Google Scholar]
- Swapnil, S.J.; Arpana, H.J. Applications of Lipases. Res. J. Biotechnol. 2019, 14, 130–138. [Google Scholar]
- Mahfoudhi, A.; Benmabrouk, S.; Fendri, A.; Sayari, A. Fungal lipases as biocatalysts: A promising platform in several industrial biotechnology applications. Biotechnol. Bioeng. 2022, 119, 3370–3392. [Google Scholar] [CrossRef]
- De Gonzalo, G.; Alcántara, A.R.; Domínguez de María, P.; Sánchez-Montero, J.M. Biocatalysis for the asymmetric synthesis of Active Pharmaceutical Ingredients (APIs): This time is for real. Expert Opin. Drug Deliv. 2022, 17, 1159–1171. [Google Scholar] [CrossRef]
- Sanchez Montero, J. Biotechnology: Impact on our Life. Drug Des. Dev. Deliv. J. 2018, 1, 103. [Google Scholar]
- Muñoz Solano, D.; Hoyos, P.; Hernáiz, M.J.; Alcántara, A.R.; Sánchez-Montero, J.M. Industrial biotransformations in the synthesis of building blocks leading to enantiopure drugs. Bioresour. Technol. 2012, 115, 196–207. [Google Scholar] [CrossRef]
- Hoyos, P.; Pace, V.; Hernáiz, M.J.; Alcántara, A.R. Biocatalysis in the Pharmaceutical Industry. A greener future. Curr. Green Chem. 2014, 1, 155–181. [Google Scholar] [CrossRef]
- Patel, R.N. Chapter 11—Applications of Biocatalysis for Pharmaceuticals and Chemicals. In Organic Synthesis Using Biocatalysis; Goswami, A., Stewart, J.D., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 339–411. [Google Scholar]
- Patel, A.K.; Singhania, R.R.; Pandey, A. Novel enzymatic processes applied to the food industry. Curr. Opin. Food Sci. 2016, 7, 64–72. [Google Scholar] [CrossRef]
- Yushkova, E.D.; Nazarova, E.A.; Matyuhina, A.V.; Noskova, A.O.; Shavronskaya, D.O.; Vinogradov, V.V.; Skvortsova, N.N.; Krivoshapkina, E.F. Application of Immobilized Enzymes in Food Industry. J. Agric. Food Chem. 2019, 67, 11553–11567. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, C.S.; Angelotti, J.A.F.; Fernandez-Lafuente, R.; Hirata, D.B. Lipase immobilization via cross-linked enzyme aggregates: Problems and prospects—A review. Int. J. Biol. Macromol. 2022, 215, 434–449. [Google Scholar] [CrossRef]
- Thangaraj, B.; Solomon, P.R. Immobilization of Lipases—A Review. Part I: Enzyme Immobilization. ChemBioEng Rev. 2019, 6, 157–166. [Google Scholar] [CrossRef]
- Thangaraj, B.; Solomon, P.R. Immobilization of Lipases—A Review. Part II: Carrier Materials. ChemBioEng Rev. 2019, 6, 167–194. [Google Scholar] [CrossRef]
- Alnoch, R.C.; Alves dos Santos, L.; Marques de Almeida, J.; Krieger, N.; Mateo, C. Recent Trends in Biomaterials for Immobilization of Lipases for Application in Non-Conventional Media. Catalysts 2020, 10, 697. [Google Scholar] [CrossRef]
- Liu, J.; Ma, R.T.; Shi, Y.P. Recent advances on support materials for lipase immobilization and applicability as biocatalysts in inhibitors screening methods-A review. Anal. Chim. Acta 2020, 1101, 9–22. [Google Scholar] [CrossRef]
- Aggarwal, S.; Chakravarty, A.; Ikram, S. A comprehensive review on incredible renewable carriers as promising platforms for enzyme immobilization & thereof strategies. Int. J. Biol. Macromol. 2021, 167, 962–986. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, V.; Kaushal, D.; Dhiman, V.K.; Kanwar, S.S.; Singh, D.; Dhiman, V.K.; Pandey, H. An Insight in Developing Carrier-Free Immobilized Enzymes. Front. Bioeng. Biotechnol. 2022, 10, 794411. [Google Scholar] [CrossRef] [PubMed]
- Cuenca, S.; Mansilla, C.; Aguado, M.; Yuste-Calvo, C.; Sánchez, F.; Sánchez-Montero, J.M.; Ponz, F. Nanonets Derived from Turnip Mosaic Virus as Scaffolds for Increased Enzymatic Activity of Immobilized Candida antarctica Lipase B. Front. Plant Sci. 2016, 7, 464. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.L.; Choi, D.M.; Xu, H.L.; Baeck, S.H.; Row, K.H.; Tang, W.Y. Lipase-based MIL-100(Fe) biocomposites as chiral stationary phase for high-efficiency capillary electrochromatographic enantioseparation. Mikrochim. Acta 2023, 190, 84. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, O.; Ortiz, C.; Berenguer-Murcia, A.; Torres, R.; Rodrigues, R.C.; Fernandez-Lafuente, R. Glutaraldehyde in bio-catalysts design: A useful crosslinker and a versatile tool in enzyme immobilization. RSC Adv. 2014, 4, 1583–1600. [Google Scholar] [CrossRef]
- German, N.; Ramanaviciene, A.; Voronovic, J.; Ramanavicius, A. Glucose biosensor based on graphite electrodes modified with glucose oxidase and colloidal gold nanoparticles. Microchim. Acta 2010, 168, 221–229. [Google Scholar] [CrossRef]
- Ramanavicius, A. Amperometric biosensor for the determination of creatine. Anal. Bioanal. Chem. 2007, 387, 1899–1906. [Google Scholar] [CrossRef]
- Betancor, L.; López-Gallego, F.; Hidalgo, A.; Alonso-Morales, N.; Dellamora-Ortiz, G.; CesarMateo, G.D.; Fernández-Lafuente, R.; Guisán, J.M. Different mechanisms of protein immobilization on glutaraldehyde activated supports: Effect of support activation and immobilization conditions. Enzym. Microb. Technol. 2006, 39, 877–882. [Google Scholar] [CrossRef]
- López-Gallego, F.; Guisan, J.M.; Betancor, L. Immobilization of Enzymes on Supports Activated with Glutaraldehyde: A Very Simple Immobilization Protocol. In Immobilization of Enzymes and Cells: Methods and Protocols; Guisan, J.M., Bolivar, J.M., López-Gallego, F., Rocha-Martín, J., Eds.; Springer: New York, NY, USA, 2020; pp. 119–127. [Google Scholar]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Castañeda-Valbuena, D.; Berenguer-Murcia, Á.; Kamli, M.R.; Tavano, O.; Fernandez-Lafuente, R. Immobilization of papain: A review. Int. J. Biol. Macromol. 2021, 188, 94–113. [Google Scholar] [CrossRef]
- Sheldon, R.A. CLEAs, Combi-CLEAs and “Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy. Catalysts 2019, 9, 31. [Google Scholar] [CrossRef]
- Sheldon, R.A. Characteristic features and biotechnological applications of cross-linked enzyme aggregates (CLEAs). Appl. Microbiol. Biotechnol. 2011, 92, 467–477. [Google Scholar] [CrossRef]
- Roy, I.; Mukherjee, J.; Gupta, M.N. Cross-Linked Enzyme Aggregates for Applications in Aqueous and Nonaqueous Media. Methods Mol. Biol. 2017, 1504, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Chang, B.; Shi, N.; Yan, W.; Lu, F.; Liu, F. Cross-linked enzyme aggregates immobilization: Preparation, characterization, and applications. Crit. Rev. Biotechnol. 2022, 43, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Cavalcante, F.T.T.; Cavalcante, A.L.G.; de Sousa, I.G.; Neto, F.S.; dos Santos, J.C.S. Current Status and Future Perspectives of Supports and Protocols for Enzyme Immobilization. Catalysts 2021, 11, 1222. [Google Scholar] [CrossRef]
- Galvis, M.; Barbosa, O.; Ruiz, M.; Cruz, J.; Ortiz, C.; Torres, R.; Fernandez-Lafuente, R. Chemical amination of lipase B from Candida antarctica is an efficient solution for the preparation of crosslinked enzyme aggregates. Process Biochem. 2012, 47, 2373–2378. [Google Scholar] [CrossRef]
- Guérin-Dubiard, C.; Pasco, M.; Mollé, D.; Désert, C.; Croguennec, T.; Nau, F. Proteomic Analysis of Hen Egg White. J. Agric. Food Chem. 2006, 54, 3901–3910. [Google Scholar] [CrossRef]
- Mayer, J.M. Stereoselective metabolism of anti-inflammatory 2-arylpropionates. Acta Pharm. Nord. 1990, 2, 197–216. [Google Scholar]
- Chen, C.S.; Fujimoto, Y.; Girdaukas, G.; Sih, C.J. Quantitative Analyses of Biochemical Kinetic Resolutions of Enantiomers. J. Am. Chem. Soc. 1982, 104, 7294–7299. [Google Scholar] [CrossRef]
- Straathof, A.J.J.; Jongejan, J.A. The enantiomeric ratio: Origin, determination and prediction. Enzym. Microb. Technol. 1997, 21, 559–571. [Google Scholar] [CrossRef]
- Jiaojiao, X.; Yan, Y.; Bin, Z.; Feng, L. Improved catalytic performance of carrier-free immobilized lipase by advanced cross-linked enzyme aggregates technology. Bioprocess Biosyst. Eng. 2022, 45, 147–158. [Google Scholar] [CrossRef]
- Hu, Y.; Shi, C.Y.; Xun, X.M.; Huang, B.R.; You, S.; Wu, F.A.; Wang, J. Xylanase-polymer conjugates as new catalysts for xylooligosaccharides production from lignocellulose. Biochem. Eng. J. 2021, 171, 108025. [Google Scholar] [CrossRef]
- Modenez, I.A.; Sastre, D.E.; Moares, F.C.; Marques Netto, C.G.C. Influence of glutaraldehyde cross-linking modes on the recyclability of immobilized lipase B from Candida antarctica for transesterification of soy bean oil. Molecules 2018, 23, 2230. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, O.; Torres, R.; Ortiz, C.; Fernandez-Lafuente, R. The slow-down of the CALB immobilization rate permits to control the inter and intra molecular modification produced by glutaraldehyde. Process Biochem. 2012, 47, 766–774. [Google Scholar] [CrossRef]
- Wilson, L.; Fernández-Lorente, G.; Fernández-Lafuente, R.; Illanes, A.; Guisán, J.M.; Palomo, J.M. CLEAs of lipases and poly-ionic polymers: A simple way of preparing stable biocatalysts with improved properties. Enzym. Microb. Technol. 2006, 39, 750–755. [Google Scholar] [CrossRef]
- Cruz, J.; Barbosa, O.; Rodrigues, R.C.; Fernandez-Lafuente, R.; Torres, R.; Ortiz, C. Optimized preparation of CALB-CLEAs by response surface methodology: The necessity to employ a feeder to have an effective crosslinking. J. Mol. Catal. B Enzym. 2012, 80, 7–14. [Google Scholar] [CrossRef]
- Chen, G.J.; Kuo, C.H.; Chen, C.I.; Yu, C.C.; Shieh, C.J.; Liu, Y.C. Effect of membranes with various hydrophobic/hydrophilic properties on lipase immobilized activity and stability. J. Biosci. Bioeng. 2012, 113, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.H.; Peng, L.T.; Kan, S.C.; Liu, Y.C.; Shieh, C.J. Lipase-immobilized biocatalytic membranes for biodiesel production. Bioresour. Technol. 2013, 145, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Yoo, H.Y.; Kim, S.W. Chapter 2—Selection and modification of enzymes prior to immobilization. In Biocatalyst Immobilization; Ferreira, M.L., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 17–35. [Google Scholar]
- Henley, J.P.; Sadana, A. Categorization of enzyme deactivations using a series-type mechanism. Enzym. Microb. Technol. 1985, 7, 50–60. [Google Scholar] [CrossRef]
- Sadana, A.; Henley, J.P. Single-step unimolecular non-first-order enzyme deactivation kinetics. Biotechnol. Bioeng. 1987, 30, 717–723. [Google Scholar] [CrossRef]
- Alcantara, A.R.; Sanchez-Montero, M.J.; Sinisterra, J.V. Chemoenzymatic Preparation of Enantiomerically Pure S(+)-2-Arylpropionic Acids with Anti-inflammatory Activity. In Stereoselective Biocatalysis; Patel, R.M., Ed.; CRC Press: Boca Raton, FL, USA, 2000; pp. 659–702. [Google Scholar]
- Gandomkar, S.; Habibi, Z.; Mohammadi, M.; Yousefi, M.; Salimi, S. Enantioselective resolution of racemic ibuprofen esters using different lipases immobilized on epoxy-functionalized silica. Biocatal. Agric. Biotechnol. 2015, 4, 550–554. [Google Scholar] [CrossRef]
- José, C.; Toledo, M.V.; Briand, L.E. Enzymatic kinetic resolution of racemic ibuprofen: Past, present and future. Crit. Rev. Biotechnol. 2016, 36, 891–903. [Google Scholar] [CrossRef]
- Brabcova, J.; Filice, M.; Gutarra, M.; Palomo, J.M. Synthesis of enantiopure drugs and drug intermediates by immobilized lipase-catalysis. Curr. Bioact. Compd. 2013, 9, 113–136. [Google Scholar] [CrossRef]
- Contesini, F.J.; Junior, J.V.M.; Macedo, G.A.; Sato, H.H.; de Oliveira Carvalho, P. Lipases in the pharmaceutical industry: An approach for racemic drugs. In Lipase: Functions, Synthesis and Role in Disease; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2012; pp. 171–184. [Google Scholar]
- López, N.; Pérez, R.; Vázquez, F.; Valero, F.; Sánchez, A. Immobilisation of different Candida rugosa lipases by adsorption onto polypropylene powder: Application to chiral synthesis of ibuprofen and trans-2-phenyl-1-cyclohexanol esters. J. Chem. Technol. Biotechnol. 2002, 77, 175–182. [Google Scholar] [CrossRef]
- De La Casa, R.M.; Guisán, J.M.; Sánchez-Montero, J.M.; Sinisterra, J.V. Modification of the activities of two different lipases from Candida rugosa with dextrans. Enzym. Microb. Technol. 2002, 30, 30–40. [Google Scholar] [CrossRef]
- Çakmak, R.; Topal, G.; Çınar, E. Covalent Immobilization of Candida rugosa Lipase on Epichlorohydrin-Coated Magnetite Nanoparticles: Enantioselective Hydrolysis Studies of Some Racemic Esters and HPLC Analysis. Appl. Biochem. Biotechnol. 2020, 191, 1411–1431. [Google Scholar] [CrossRef] [PubMed]
- James, J.J.; Lakshmi, B.S.; Raviprasad, V.; Ananth, M.J.; Kangueane, P.; Gautam, P. Insights from molecular dynamics simulations into pH-dependent enantioselective hydrolysis of ibuprofen esters by Candida rugosa lipase. Protein Eng. 2003, 16, 1017–1024. [Google Scholar] [CrossRef]
- Lakshmi, B.S.; Kangueane, P.; Guo, Y.; Chen, Y.Z.; Gautam, P. Molecular basis for the stereospecificity of Candida rugosa lipase (CRL) towards ibuprofen. Biocatal. Biotransform. 2000, 17, 475–486. [Google Scholar] [CrossRef]
- Alcántara, A.R.; de Fuentes, I.E.; Sinisterra, J.V. Rhizomucor miehei lipase as the catalyst in the resolution of chiral compounds: An overview. Chem. Phys. Lipids 1998, 93, 169–184. [Google Scholar] [CrossRef]
- Habibi, Z.; Mohammadi, M.; Yousefi, M. Enzymatic hydrolysis of racemic ibuprofen esters using Rhizomucor miehei lipase immobilized on different supports. Process Biochem. 2013, 48, 669–676. [Google Scholar] [CrossRef]
- Henke, E.; Schuster, S.; Yang, H.; Bornscheuer, U.T. Lipase-catalyzed resolution of ibuprofen. Monatsh. Chem. 2000, 131, 633–638. [Google Scholar] [CrossRef]
- Li, H.X.; Xie, Z.B.; Chen, G.Q.; Zhang, C.; Lei, G.Y.; Le, Z.G. Study on the Process of Resolution of Ibuprofen Ethyl Ester by Enzymatic Hydrolysis. J. Mol. Catal. 2019, 33, 447–452. [Google Scholar]
- Sandström, A.G.; Wikmark, Y.; Engström, K.; Nyhlén, J.; Bäckvall, J.E. Combinatorial reshaping of the Candida antarctica lipase A substrate pocket for enantioselectivity using an extremely condensed library. Proc. Natl. Acad. Sci. USA 2012, 109, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.C.L.M.; Fonseca, T.S.; De Mattos, M.C.; De Oliveira, M.C.F.; De Lemos, T.L.G.; Molinari, F.; Romano, D.; Serra, I. Recent advances in lipase-mediated preparation of pharmaceuticals and their intermediates. Int. J. Mol. Sci. 2015, 16, 29682–29716. [Google Scholar] [CrossRef]
- Faber, K. Biotransformations in Organic Chemistry: A Textbook, 7th Extended and Corrected ed.; Springer International Publishing AG: Cham, Switzerland, 2018; pp. 31–313. [Google Scholar]
- Monteiro, R.R.C.; Virgen-Ortiz, J.J.; Berenguer-Murcia, A.; da Rocha, T.N.; dos Santos, J.C.S.; Alcantara, A.R.; Fernandez-Lafuente, R. Biotechnological relevance of the lipase A from Candida antarctica. Catal. Today 2021, 362, 141–154. [Google Scholar] [CrossRef]
- Velasco-Lozano, S.; López-Gallego, F.; Rocha-Martin, J.; Guisán, J.M.; Favela-Torres, E. Improving enantioselectivity of lipase from Candida rugosa by carrier-bound and carrier-free immobilization. J. Mol. Catal. B Enzym. 2016, 130, 32–39. [Google Scholar] [CrossRef]
- Xiao, X.; Liu, Z.; Chen, Y.; Wang, G.; Li, X.; Fang, Z.; Huang, S.; Liu, Z.; Yan, Y.; Xu, L. Over-expression of active Candida rugosa lip1 in Pichia pastoris via high cell-density fermentation and its application to resolve racemic ibuprofen. Biocatal. Biotransform. 2015, 33, 260–269. [Google Scholar] [CrossRef]
- Salgın, S.; Çakal, M.; Salgın, U. Kinetic resolution of racemic naproxen methyl ester by magnetic and non-magnetic cross-linked lipase aggregates. Prep. Biochem. Biotechnol. 2020, 50, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Sampath, C.; Belur, P.D.; Iyyasami, R. Enhancement of n-3 polyunsaturated fatty acid glycerides in Sardine oil by a bioimprinted cross-linked Candida rugosa lipase. Enzym. Microb. Technol. 2018, 110, 20–29. [Google Scholar] [CrossRef]
- Bradford, M.M. Rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Fu, S.; Liu, H.; Lucia, L.A. Accurately Determining Esterase Activity via the Isosbestic Point of p-Nitrophenol. BioResources 2016, 11, 10099–10111. [Google Scholar] [CrossRef]
- Galliani, M.; Santi, M.; Del Grosso, A.; Cecchettini, A.; Santorelli, F.M.; Hofmann, S.L.; Lu, J.Y.; Angella, L.; Cecchini, M.; Signore, G. Cross-Linked Enzyme Aggregates as Versatile Tool for Enzyme Delivery: Application to Polymeric Nanoparticles. Bioconjugate Chem. 2018, 29, 2225–2231. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Case | Molar Equivalents. CAL-B.: GA: PTR. | V GA./V% (µL) | V PTR. (µL) | I.Y. (%) | S.A. CLEAs (U/mg) | A.Y. (%) |
|---|---|---|---|---|---|---|---|
| G1 | 01 | 1: 250: 250 | 15/0.15 | 110 | 10 | 35 | 100 |
| G2 | 02 | 1: 500: 250 | 30/0.30 | 110 | 13 | 35 | 100 |
| 03 | 1: 500: 500 | 30/0.30 | 220 | 15 | 34 | 96 | |
| 04 | 1: 500: 1000 | 30/0.30 | 440 | 18 | 34 | 97 | |
| G3 | 05 | 1: 1000: 250 | 60/0.60 | 110 | 15 | 35 | 100 |
| 06 | 1: 1000: 500 | 60/0.60 | 220 | 19 | 35 | 100 | |
| 07 | 1: 1000: 1000 | 60/0.60 | 440 | 25 | 36 | 103 | |
| G4 | 08 | 1: 2000: 250 | 120/1.18 | 110 | 20 | 34 | 97 |
| 09 | 1: 2000: 500 | 120/1.18 | 220 | 23 | 35 | 100 | |
| 10 | 1: 2000: 1000 | 120/1.18 | 440 | 35 | 37 | 106 | |
| 11 | 1: 2000: 2000 | 120/1.18 | 880 | 29 | 30 | 86 |
| Enzyme | pH Jump (S2–S3) |
|---|---|
| CRL | 5.46–8.05 |
| CAL-B | 5.48–8.19 |
| CAL-B* | 8.19–12.0 |
| CAL-A | 5.81–8.02 |
| LU | 5.90–8.08 |
| TLL | 5.79–8.15 |
| RML | 5.98–8.08 |
| Lipases | Enzyme (mg) | I.Y (%) | WCLEA-LC S3* (g) | S.A. Free Enzyme (U/mg) | S.A. CLEA-LC S3* (U/mg) | CLEA-LC Activity S3* (U/g) | %H2O S3* |
|---|---|---|---|---|---|---|---|
| CRL | 50 | 99 | 8.5 | 800 | 600 | 3494 | 93.9 |
| CAL-B | 50 | 35 | 1.36 | 35 | 37 | 476 | 93.8 |
| CAL-B* | 50 | >99 | 6.5 | 35 | >40 | 307 | 93.7 |
| CAL-A | 50 | 83 | 1.73 | 65 | 50 | 1199 | 93.8 |
| TLL | 50 | 72 | 1.4 | 17 | 18 | 468 | 94.2 |
| LU | 50 | 74 | 1.47 | 12 | 12 | 302 | 93.1 |
| RML | 50 | 99 | 1 | 14 | 15 | 745 | 93.7 |
| Enzyme | Free Form | CLEA-LC Form | ||||
|---|---|---|---|---|---|---|
| C (%) | eep (%), SP. | E | C (%) | eep (%), sp. | E | |
| CRL | 49 | 97, (S) | >200 | 40 | 99, (S) | >200 |
| CAL-B | 76 | 27, (R) | ---- | 64 | 30, (R) | ---- |
| CAL-B* | 25 | 15, (R) | ---- | 45 | 48, (R) | 8.47 |
| CAL-A | 23 | 22, (S) | 3 | 46 | 66, (R) | 14.30 |
| RML | 81 | 22, (S) | ---- | 47 | 44, (S) | 9.24 |
| LU | 8 | 06, (R) | 2 | 33 | 47, (R) | 5.56 |
| TLL | 74 | 75, (S) | ---- | 3.4 | >99 | 2.24 |
| T (°C) | t1/2 (h) | A1 | A2 | A3 | k1 | k2 | k3 | R2 | |
|---|---|---|---|---|---|---|---|---|---|
| CRL | 25 | 192 | −1.11 × 103 | 8.76 × 101 | 1.13 × 103 | 1.33 × 10−2 | 1.21 × 10−2 | 1.19 × 10−2 | 0.9836 |
| 40 | 35 | 1.05 × 102 | --- | --- | 2.29 × 10−2 | --- | --- | 0.981 | |
| 50 | 0.67 | 9.56 × 101 | --- | --- | 1.06 | --- | --- | 0.9891 | |
| 60 | 0.5 | 1.01 × 102 | --- | --- | 4.01 | 0.9977 | |||
| 70 a | --- | --- | --- | --- | --- | --- | --- | --- | |
| CLEA-LC | 25 b | --- | --- | --- | --- | --- | --- | --- | --- |
| 40 | 312 | −1.66 × 101 | −1.23 × 101 | 1.29 × 102 | 2.51 × 10−2 | 2.51 × 10−3 | 2.51 × 10−3 | 0.9976 | |
| 50 | 96 | −3.12 × 102 | 1.39 × 101 | 3.97 × 102 | 2.86 × 10−2 | 1.98 × 10−2 | 1.99 × 10−2 | 0.9961 | |
| 60 | 2 | 4.65 × 101 | 5.15 × 101 | --- | 5.30 × 10−2 | 1.02 | --- | 0.9986 | |
| 70 | <0.5 | 2.80 × 101 | 7.20 × 101 | --- | 1.33 | 1.52 × 101 | 0.9984 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khiari, O.; Bouzemi, N.; Sánchez-Montero, J.M.; Alcántara, A.R. Easy and Versatile Technique for the Preparation of Stable and Active Lipase-Based CLEA-like Copolymers by Using Two Homofunctional Cross-Linking Agents: Application to the Preparation of Enantiopure Ibuprofen. Int. J. Mol. Sci. 2023, 24, 13664. https://doi.org/10.3390/ijms241713664
Khiari O, Bouzemi N, Sánchez-Montero JM, Alcántara AR. Easy and Versatile Technique for the Preparation of Stable and Active Lipase-Based CLEA-like Copolymers by Using Two Homofunctional Cross-Linking Agents: Application to the Preparation of Enantiopure Ibuprofen. International Journal of Molecular Sciences. 2023; 24(17):13664. https://doi.org/10.3390/ijms241713664
Chicago/Turabian StyleKhiari, Oussama, Nassima Bouzemi, José María Sánchez-Montero, and Andrés R. Alcántara. 2023. "Easy and Versatile Technique for the Preparation of Stable and Active Lipase-Based CLEA-like Copolymers by Using Two Homofunctional Cross-Linking Agents: Application to the Preparation of Enantiopure Ibuprofen" International Journal of Molecular Sciences 24, no. 17: 13664. https://doi.org/10.3390/ijms241713664