
The ATP Synthase Subunits FfATPh, FfATP5, and FfATPb Regulate the Development, Pathogenicity, and Fungicide Sensitivity of Fusarium fujikuroi

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
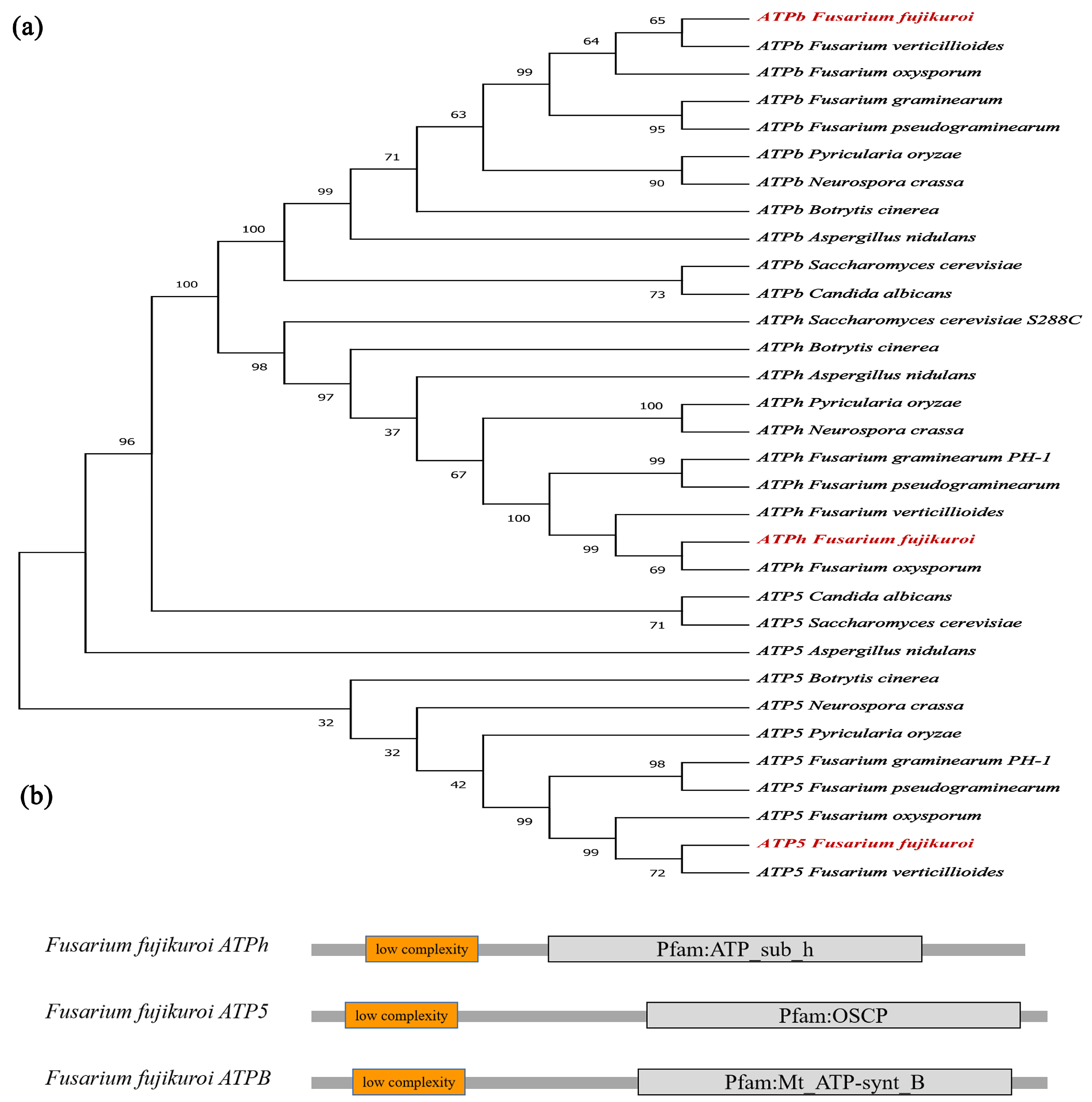
2.1. Identification of FfATPh, FfATP5, and FfATPb
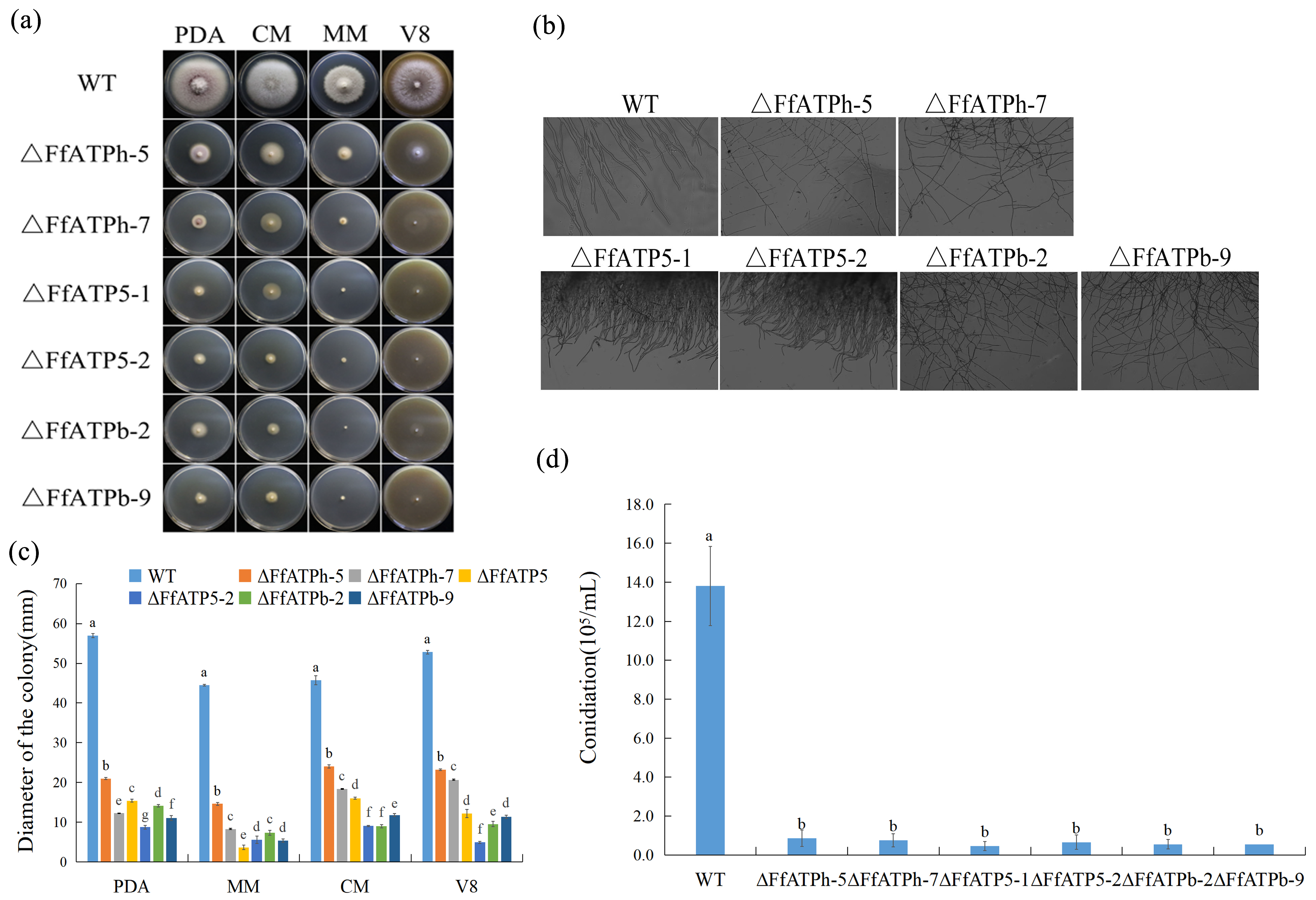
2.2. Regulation of FfATPh, FfATP5, and FfATPb in Vegetative Growth
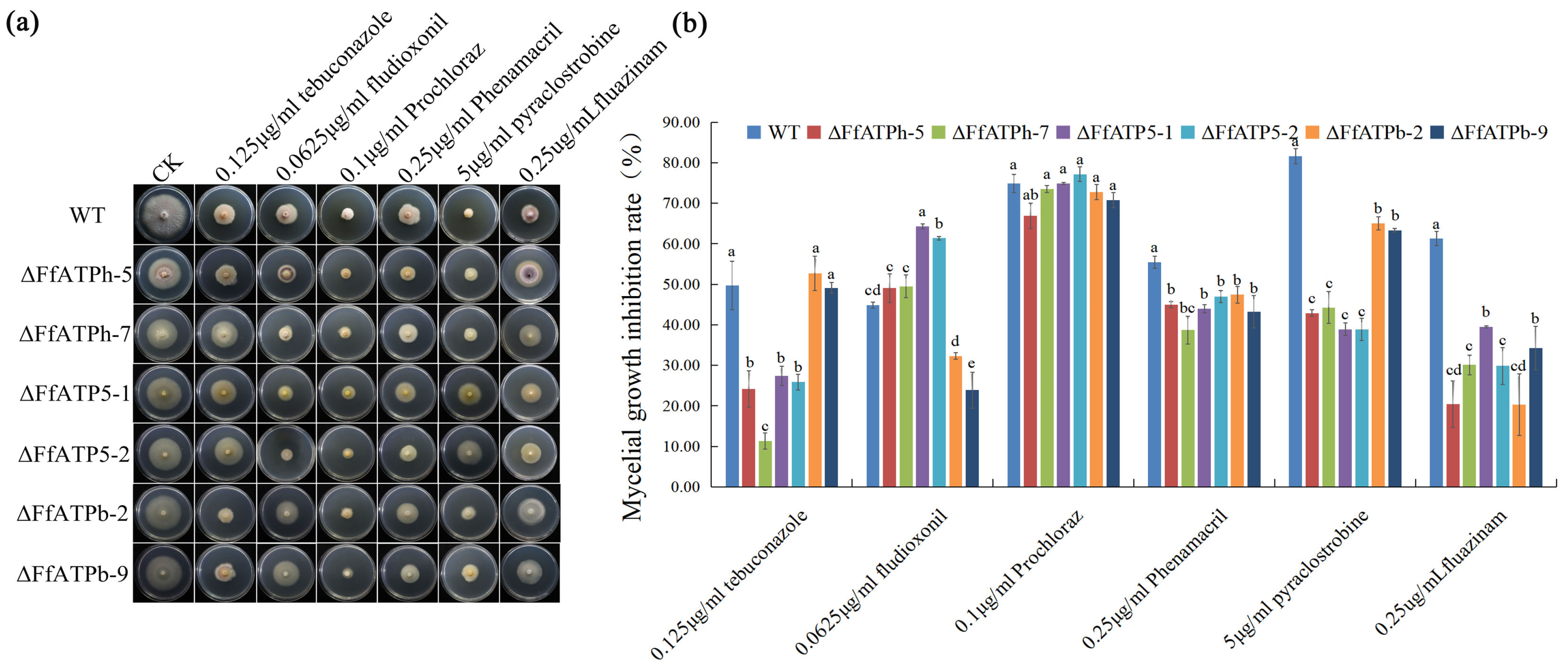
2.3. FfATPh, FfATP5, and FfATPb Regulate the Sensitivity to Different Fungicides
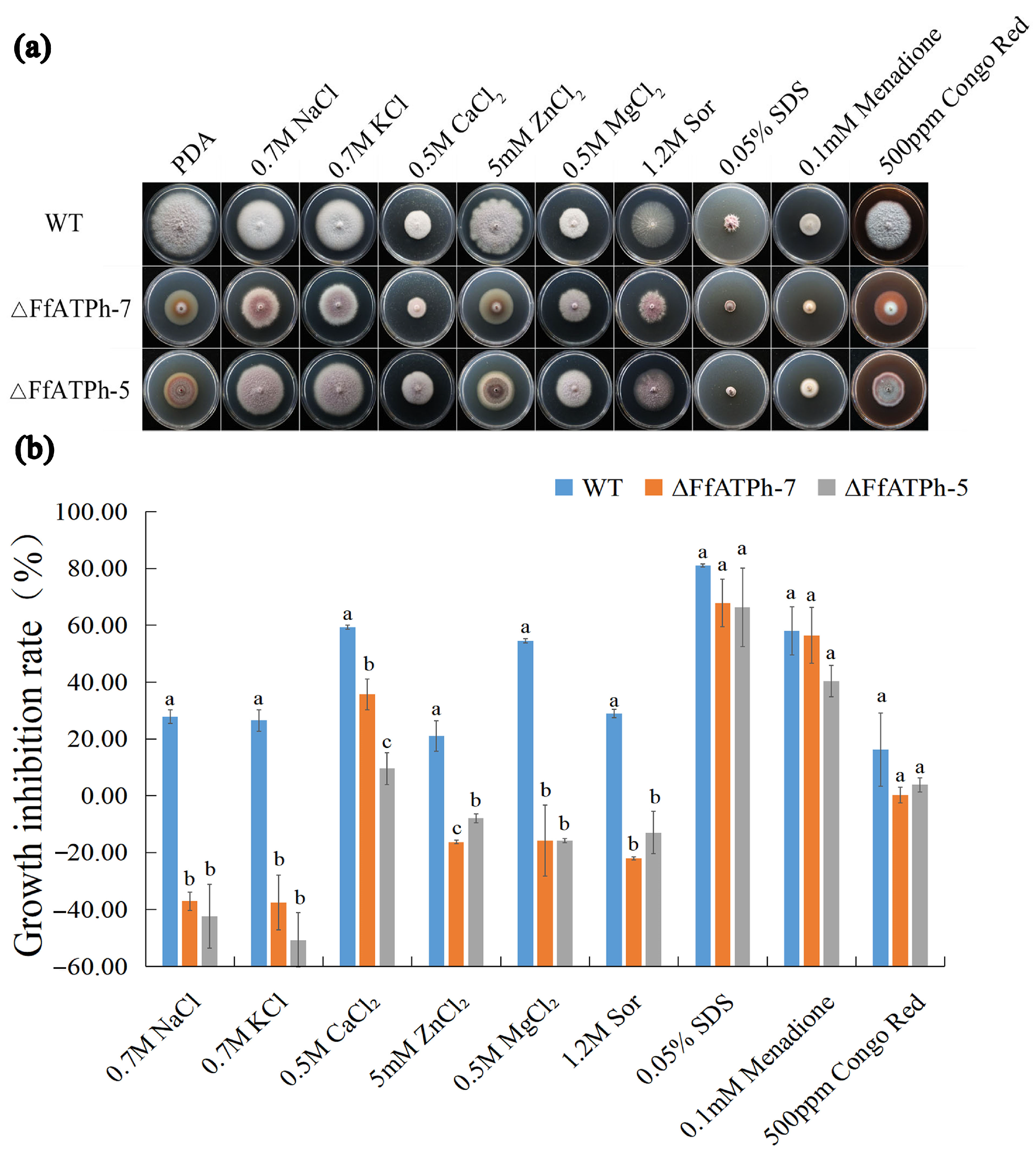
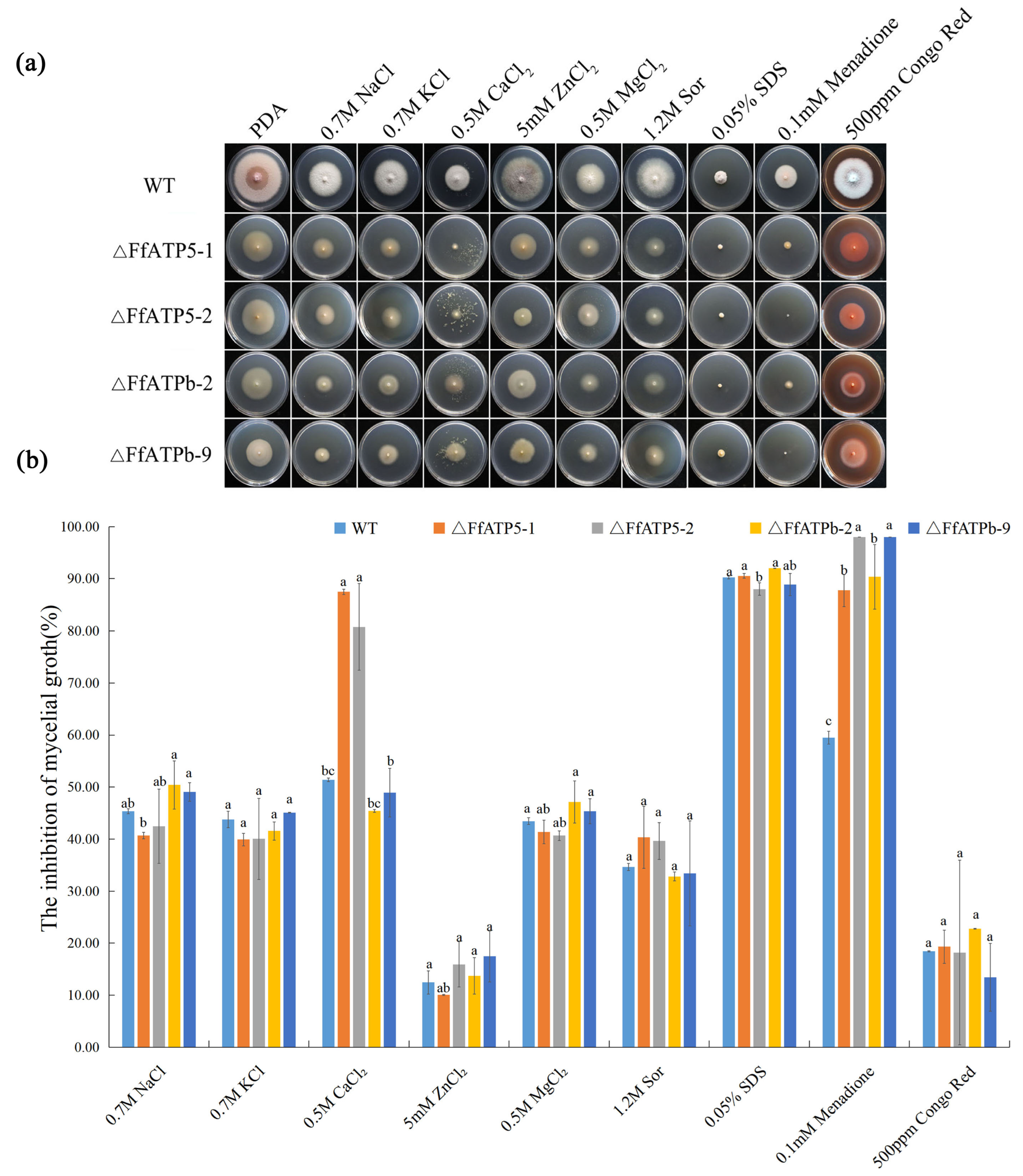
2.4. FfATPh, FfATP5, and FfATPb Are Involved in the Regulation of the Sensitivity to Various Stresses
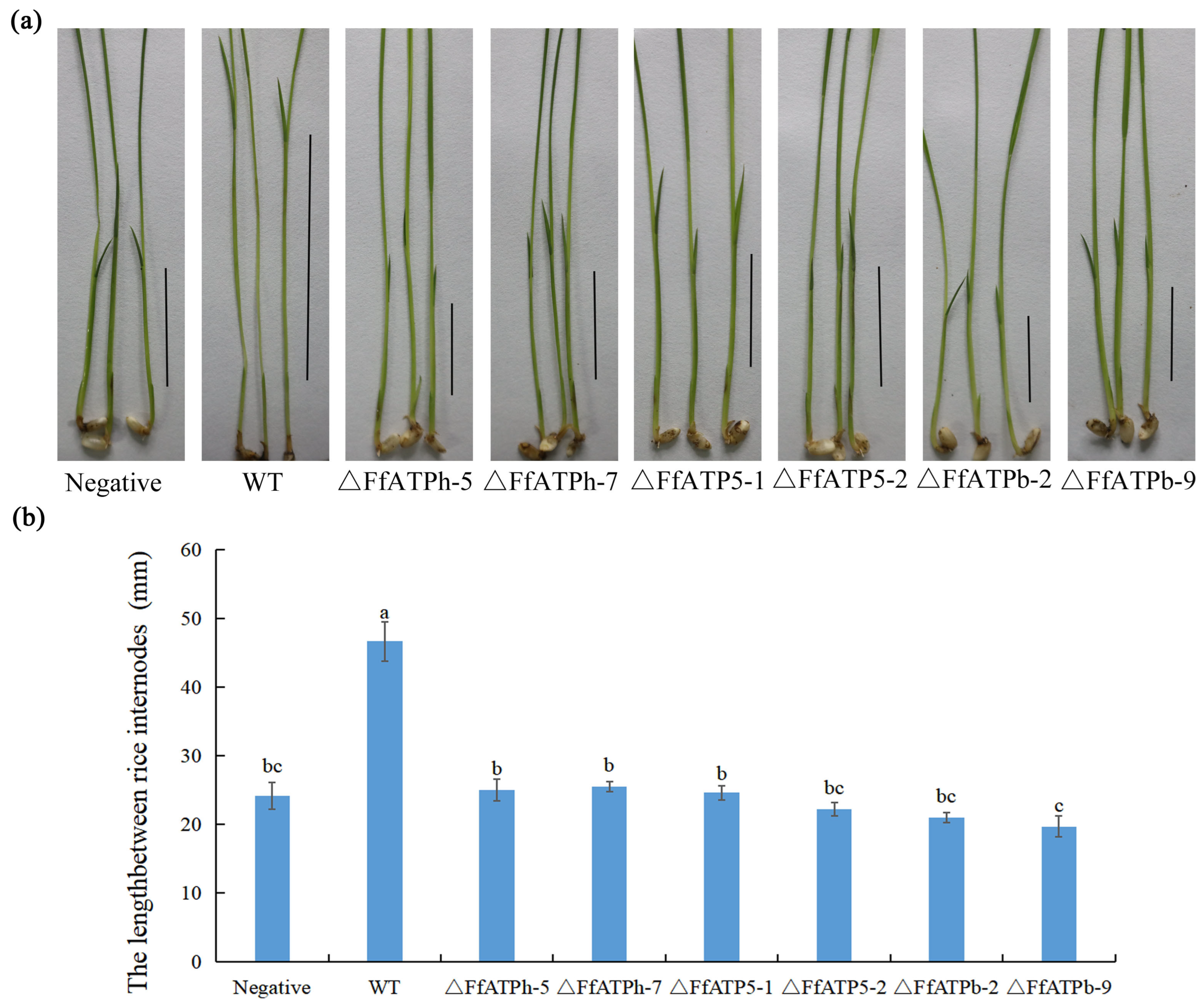
2.5. FfATPh, FfATPb, and FfATP5 Are Indispensable for Pathogenicity
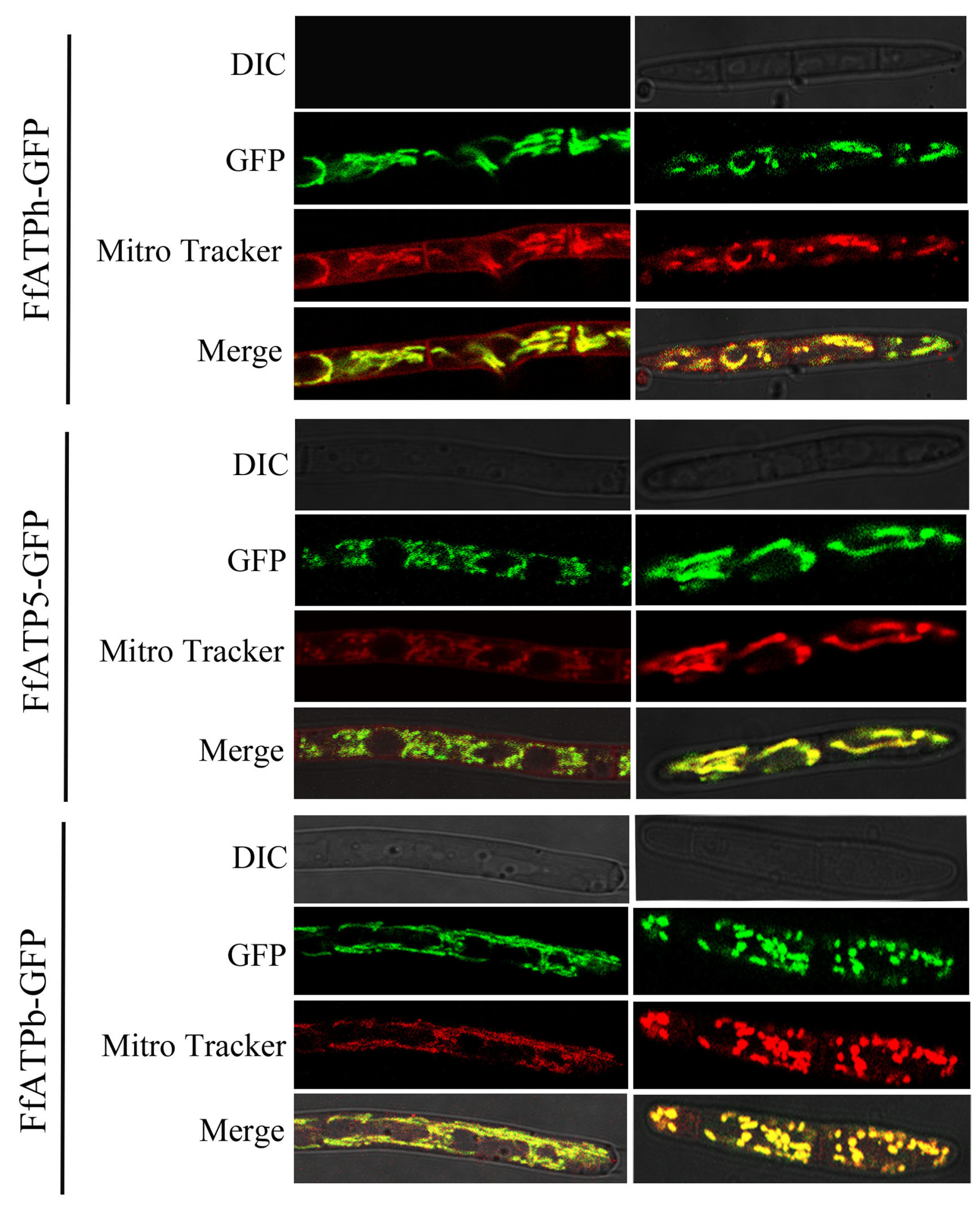
2.6. FfATPh, FfATP5, and FfATPb Are Located in Mitochondria
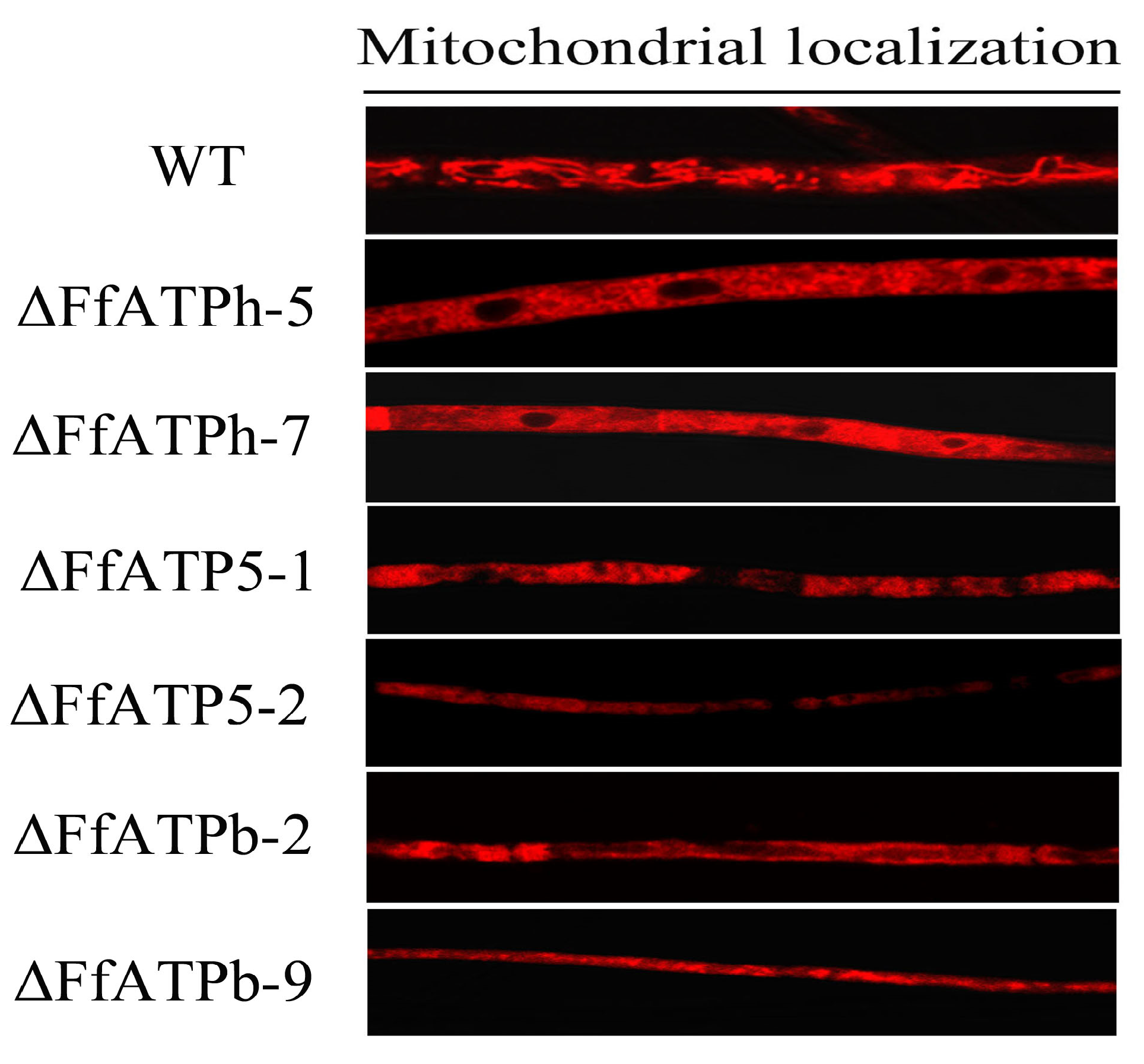
2.7. FfATPh, FfATP5, and FfATPb Regulate the Localization of Mitochondria
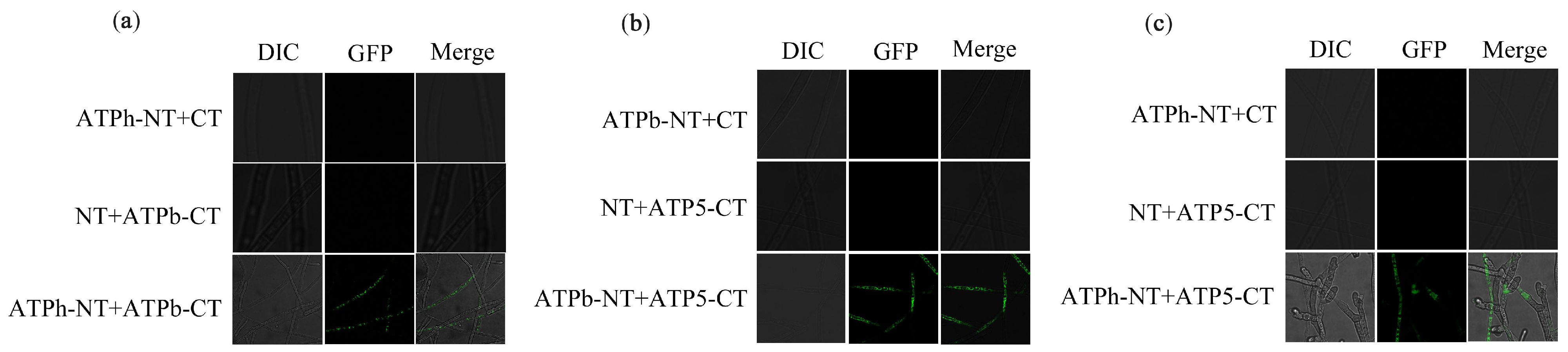
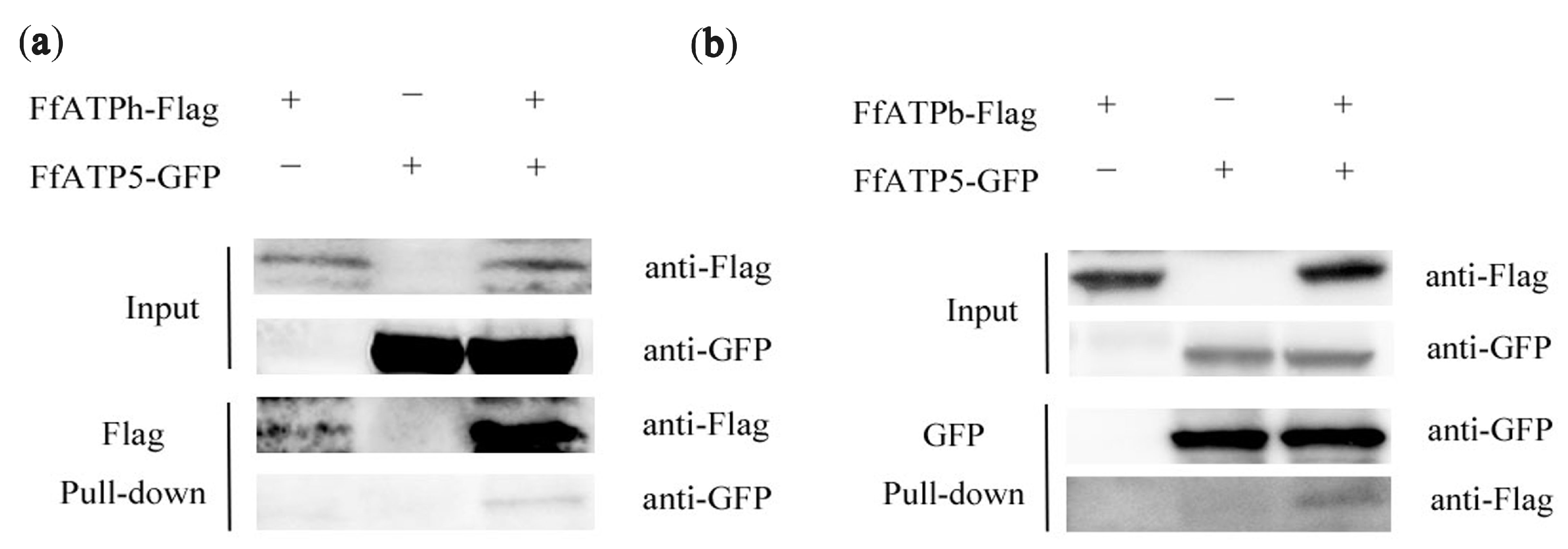
2.8. Interaction among FfATPh, FfATP5, and FfATPb
3. Discussion
4. Materials and Methods
4.1. Strains, Media, and Fungicides
4.2. Construction of Deletion Mutant Vectors, GFP, and Flag Fusion Cassettes
4.3. Vegetative Growth and Asexual Reproduction Assays
4.4. Sensitivity of Strains to Different Fungicides and Stress Factors
4.5. Pathogenicity Assay
4.6. Microscopic Examinations
4.7. Western Blotting Hybridization
4.8. Bimolecular Fluorescent Complimentary (BiFC) Assay
4.9. Co-Immunoprecipitation (Co-IP) Assay
4.10. Yeast Two-Hybrid Assay (Y2H)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ji, Z.; Zeng, Y.; Liang, Y.; Yang, C. Research and Progress of Bakanae Disease Resistance in Rice. Chin. J. Rice Sci. 2021, 35, 1. [Google Scholar]
- Matic, S.; Gullino, M.L.; Spadaro, D. The puzzle of bakanae disease through interactions between Fusarium fujikuroi and rice. Front. Biosci. (Elite Ed.) 2017, 9, 333–344. [Google Scholar] [PubMed]
- Yilmaz, N.; Sandoval-Denis, M.; Lombard, L.; Visagie, C.M.; Wingfield, B.D.; Crous, P.W. Redefining species limits in the Fusarium fujikuroi species complex. Persoonia 2021, 46, 129–162. [Google Scholar] [CrossRef] [PubMed]
- Piombo, E.; Rosati, M.; Sanna, M.; Mezzalama, M.; Gullino, M.L.; Spadaro, D. Sequencing of non-virulent strains of Fusarium fujikuroi reveals genes putatively involved in bakanae disease of rice. Fungal Genet. Biol. 2021, 156, 103622. [Google Scholar] [CrossRef] [PubMed]
- Sultana, S.; Kitajima, M.; Kobayashi, H.; Nakagawa, H.; Shimizu, M.; Kageyama, K.; Suga, H. A Natural Variation of Fumonisin Gene Cluster Associated with Fumonisin Production Difference in Fusarium fujikuroi. Toxins 2019, 11, 200. [Google Scholar] [CrossRef]
- Cen, Y.-K.; Lin, J.-G.; Wang, Y.-L.; Wang, J.-Y.; Liu, Z.-Q.; Zheng, Y.-G. The Gibberellin Producer Fusarium fujikuroi: Methods and Technologies in the Current Toolkit. Front. Bioeng. Biotechnol. 2020, 8, 232. [Google Scholar] [CrossRef]
- Matic, S.; Spadaro, D.; Prelle, A.; Gullino, M.L.; Garibaldi, A. Light affects fumonisin production in strains of Fusarium fujikuroi, Fusarium proliferatum, and Fusarium verticillioides isolated from rice. Int. J. Food Microbiol. 2013, 166, 515–523. [Google Scholar] [CrossRef]
- Studt, L.; Troncoso, C.; Gong, F.; Hedden, P.; Toomajian, C.; Leslie, J.F.; Humpf, H.U.; Rojas, M.C.; Tudzynski, B. Segregation of secondary metabolite biosynthesis in hybrids of Fusarium fujikuroi and Fusarium proliferatum. Fungal Genet. Biol. 2012, 49, 567–577. [Google Scholar] [CrossRef]
- Wiemann, P.; Sieber, C.M.K.; Von Bargen, K.W.; Studt, L.; Niehaus, E.-M.; Espino, J.J.; Huss, K.; Michielse, C.B.; Albermann, S.; Wagner, D.; et al. Deciphering the Cryptic Genome: Genome-wide Analyses of the Rice Pathogen Fusarium fujikuroi Reveal Complex Regulation of Secondary Metabolism and Novel Metabolites. PLoS Pathog. 2013, 9, e1003475. [Google Scholar] [CrossRef]
- Qu, X.-P.; Li, J.-S.; Wang, J.-X.; Wu, L.-Y.; Wang, Y.-F.; Chen, C.-J.; Zhou, M.-G.; Hou, Y.-P. Effects of the dinitroaniline fungicide fluazinam on Fusarium fujikuroi and rice. Pestic. Biochem. Physiol. 2018, 152, 98–105. [Google Scholar] [CrossRef]
- Dai, L.; Wang, B.; Wang, T.; Meyer, E.H.; Kettel, V.; Hoffmann, N.; McFarlane, H.E.; Li, S.; Wu, X.; Picard, K.L.; et al. The TOR complex controls ATP levels to regulate actin cytoskeleton dynamics in Arabidopsis. Proc. Natl. Acad. Sci. USA 2022, 119, e2122969119. [Google Scholar] [CrossRef] [PubMed]
- Capaldi, R.A.; Aggeler, R.; Turina, P.; Wilkens, S. Coupling between catalytic sites and the proton channel in F1-F0-type ATPases. Trends Biochem. Sci. 1994, 19, 284–289. [Google Scholar] [CrossRef]
- Noji, H.; Ueno, H.; McMillan, D.G.G. Catalytic robustness and torque generation of the F(1)-ATPase. Biophys. Rev. 2017, 9, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Gai, X.; Li, S.; Jiang, N.; Sun, Q.; Xuan, Y.H.; Xia, Z. Comparative transcriptome analysis reveals that ATP synthases regulate Fusarium oxysporum virulence by modulating sugar transporter gene expressions in tobacco. Front. Plant Sci. 2022, 13, 978951. [Google Scholar] [CrossRef] [PubMed]
- Li, S.X.; Wu, H.T.; Liu, Y.T.; Jiang, Y.Y.; Zhang, Y.S.; Liu, W.D.; Zhu, K.J.; Li, D.M.; Zhang, H. The F(1)F(o)-ATP Synthase beta Subunit Is Required for Candida albicans Pathogenicity Due to Its Role in Carbon Flexibility. Front. Microbiol. 2018, 9, 1025. [Google Scholar] [CrossRef]
- Esparza-Perusquía, M.; Langner, T.; García-Cruz, G.; Feldbrügge, M.; Zavala, G.; Pardo, J.P.; Martínez, F.; Flores-Herrera, O. Deletion of the ATP20 gene in Ustilago maydis produces an unstable dimer of F1FO-ATP synthase associated with a decrease in mitochondrial ATP synthesis and a high H2O2 production. Biochim. Et. Biophys. Acta (BBA) Bioenerg. 2023, 1864, 148950. [Google Scholar] [CrossRef]
- Kucharczyk, R.; Zick, M.; Bietenhader, M.; Rak, M.; Couplan, E.; Blondel, M.; Caubet, S.D.; di Rago, J.P. Mitochondrial ATP synthase disorders: Molecular mechanisms and the quest for curative therapeutic approaches. Biochim. Biophys. Acta 2009, 1793, 186–199. [Google Scholar] [CrossRef]
- Leroux, P. Recent Developments in the Mode of Action of Fungicides. Pestic. Sci. 1996, 47, 191–197. [Google Scholar] [CrossRef]
- Rohricht, H.; Schwartzmann, J.; Meyer, E.H. Complexome profiling reveals novel insights into the composition and assembly of the mitochondrial ATP synthase of Arabidopsis thaliana. Biochim. Biophys. Acta Bioenerg. 2021, 1862, 148425. [Google Scholar] [CrossRef]
- Dickson, V.K.; Silvester, J.A.; Fearnley, I.M.; Leslie, A.G.; Walker, J.E. On the structure of the stator of the mitochondrial ATP synthase. EMBO J. 2006, 25, 2911–2918. [Google Scholar] [CrossRef]
- Velours, J.; Arselin, G.; Paul, M.F.; Galante, M.; Durrens, P.; Aigle, M.; Guerin, B. The yeast ATP synthase subunit 4: Structure and function. Biochimie 1989, 71, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Tzagoloff, A. Assembly of the mitochondrial membrane system. 3. Function and synthesis of the oligomycin sensitivity-conferring protein of yeast mitochondria. J. Biol. Chem. 1970, 245, 1545–1551. [Google Scholar] [CrossRef] [PubMed]
- Fronzes, R.; Chaignepain, S.; Bathany, K.; Giraud, M.-F.; Arselin, G.; Schmitter, J.-M.; Dautant, A.; Velours, J.; Brèthes, D. Topological and functional study of subunit h of the F1Fo ATP synthase complex in yeast Saccharomyces cerevisiae. Biochemistry 2003, 42, 12038–12049. [Google Scholar] [CrossRef]
- Gavin, P.D.; Prescott, M.; Luff, S.E.; Devenish, R.J. Cross-linking ATP synthase complexes in vivo eliminates mitochondrial cristae. J. Cell Sci. 2004, 117 Pt 11, 2333–2343. [Google Scholar] [CrossRef] [PubMed]
- Paumard, P.; Vaillier, J.; Coulary, B.; Schaeffer, J.; Soubannier, V.; Mueller, D.M.; Brethes, D.; di Rago, J.P.; Velours, J. The ATP synthase is involved in generating mitochondrial cristae morphology. EMBO J. 2002, 21, 221–230. [Google Scholar] [CrossRef]
- Arselin, G.; Vaillier, J.; Graves, P.-V.; Velours, J. ATP Synthase of Yeast Mitochondria. J. Biol. Chem. 1996, 271, 20284–20290. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.K.; Stewart, A.G.; Donohoe, M.; Bernal, R.A.; Stock, D. The structure of the peripheral stalk of Thermus thermophilus H+-ATPase/synthase. Nat. Struct. Mol. Biol. 2010, 17, 373–378. [Google Scholar] [CrossRef]
- Rees, D.M.; Leslie, A.G.; Walker, J.E. The structure of the membrane extrinsic region of bovine ATP synthase. Proc. Natl. Acad. Sci. USA 2009, 106, 21597–21601. [Google Scholar] [CrossRef]
- Rubinstein, J.L.; Dickson, V.K.; Runswick, M.J.; Walker, J.E. ATP synthase from Saccharomyces cerevisiae: Location of subunit h in the peripheral stalk region. J. Mol. Biol. 2005, 345, 513–520. [Google Scholar] [CrossRef]
- Bald, D.; Amano, T.; Muneyuki, E.; Pitard, B.; Rigaud, J.L.; Kruip, J.; Hisabori, T.; Yoshida, M.; Shibata, M. ATP synthesis by F0F1-ATP synthase independent of noncatalytic nucleotide binding sites and insensitive to azide inhibition. J. Biol. Chem. 1998, 273, 865–870. [Google Scholar] [CrossRef]
- Matsumoto, T.; Nakane, T.; Chiba, S. Pharmacological analysis of responses to ATP in the isolated and perfused canine coronary artery. Eur. J. Pharmacol. 1997, 334, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Tourmente, M.; Varea-Sanchez, M.; Roldan, E.R.S. Faster and more efficient swimming: Energy consumption of murine spermatozoa under sperm competition. Biol. Reprod. 2019, 100, 420–428. [Google Scholar] [CrossRef]
- Carbajo, R.J.; Silvester, J.A.; Runswick, M.J.; Walker, J.E.; Neuhaus, D. Solution structure of subunit F(6) from the peripheral stalk region of ATP synthase from bovine heart mitochondria. J. Mol. Biol. 2004, 342, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Goyon, V.; Fronzes, R.; Salin, B.; di-Rago, J.P.; Velours, J.; Brethes, D. Yeast cells depleted in Atp14p fail to assemble Atp6p within the ATP synthase and exhibit altered mitochondrial cristae morphology. J. Biol. Chem. 2008, 283, 9749–9758. [Google Scholar] [CrossRef] [PubMed]
- Robison, M.M.; Ling, X.; Smid, M.P.; Zarei, A.; Wolyn, D.J. Antisense expression of mitochondrial ATP synthase subunits OSCP (ATP5) and gamma (ATP3) alters leaf morphology, metabolism and gene expression in Arabidopsis. Plant Cell Physiol. 2009, 50, 1840–1850. [Google Scholar] [CrossRef]
- Matić, S.; Bagnaresi, P.; Biselli, C.; Orru’, L.; Amaral Carneiro, G.; Siciliano, I.; Valé, G.; Gullino, M.L.; Spadaro, D. Comparative transcriptome profiling of resistant and susceptible rice genotypes in response to the seedborne pathogen Fusarium fujikuroi. BMC Genom. 2016, 17, 608. [Google Scholar] [CrossRef]
- Sunani, S.K.; Bashyal, B.M.; Kharayat, B.S.; Prakash, G.; Krishnan, S.G.; Aggarwal, R. Identification of rice seed infection routes of Fusarium fujikuroi inciting bakanae disease of rice. J. Plant Pathol. 2020, 102, 113–121. [Google Scholar] [CrossRef]
- Yang, C.-H.; Qin, G.; Liu, J.; Cao, B.; Li, B.; Tian, S. Hydrogen Peroxide Acts on Sensitive Mitochondrial Proteins to Induce Death of a Fungal Pathogen Revealed by Proteomic Analysis. PLoS ONE 2011, 6, e21945. [Google Scholar]
- Soubannier, V.; Vaillier, J.; Paumard, P.; Coulary, B.; Schaeffer, J.; Velours, J. In the absence of the first membrane-spanning segment of subunit 4(b), the yeast ATP synthase is functional but does not dimerize or oligomerize. J. Biol. Chem. 2002, 277, 10739–10745. [Google Scholar] [CrossRef]
- Mao, X.; Wu, Z.; Chen, F.; Zhou, M.; Hou, Y. FfCOX17 is Involved in Fumonisins Production, Growth, Asexual Reproduction, and Fungicide Sensitivity in Fusarium fujikuroi. Toxins 2022, 14, 427. [Google Scholar] [CrossRef]
- Yu, J.-H.; Hamari, Z.; Han, K.-H.; Seo, J.-A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef]
- Zheng, Z.; Gao, T.; Zhang, Y.; Hou, Y.; Wang, J.; Zhou, M. FgFim, a key protein regulating resistance to the fungicide JS399-19, asexual and sexual development, stress responses and virulence in Fusarium graminearum. Mol. Plant Pathol. 2014, 15, 488–499. [Google Scholar] [CrossRef]
- Tang, G.; Chen, A.; Dawood, D.H.; Liang, J.; Chen, Y.; Ma, Z. Capping proteins regulate fungal development, DON-toxisome formation and virulence in Fusarium graminearum. Mol. Plant Pathol. 2020, 21, 173–187. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, N.; Yin, Y.; Chen, Y.; Jiang, J.; Ma, Z. Histone H3K4 methylation regulates hyphal growth, secondary metabolism and multiple stress responses in Fusarium graminearum. Environ. Microbiol. 2015, 17, 4615–4630. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Yuan, Z.; Cai, X.; Gui, S.; Zhou, M.; Hou, Y. The ATP Synthase Subunits FfATPh, FfATP5, and FfATPb Regulate the Development, Pathogenicity, and Fungicide Sensitivity of Fusarium fujikuroi. Int. J. Mol. Sci. 2023, 24, 13273. https://doi.org/10.3390/ijms241713273
Yang X, Yuan Z, Cai X, Gui S, Zhou M, Hou Y. The ATP Synthase Subunits FfATPh, FfATP5, and FfATPb Regulate the Development, Pathogenicity, and Fungicide Sensitivity of Fusarium fujikuroi. International Journal of Molecular Sciences. 2023; 24(17):13273. https://doi.org/10.3390/ijms241713273
Chicago/Turabian StyleYang, Xin, Zhili Yuan, Xiaowei Cai, Shuai Gui, Mingguo Zhou, and Yiping Hou. 2023. "The ATP Synthase Subunits FfATPh, FfATP5, and FfATPb Regulate the Development, Pathogenicity, and Fungicide Sensitivity of Fusarium fujikuroi" International Journal of Molecular Sciences 24, no. 17: 13273. https://doi.org/10.3390/ijms241713273