
LH/hCG Regulation of Circular RNA in Mural Granulosa Cells during the Periovulatory Period in Mice
 ,
,  , and
, and
Abstract
:1. Introduction
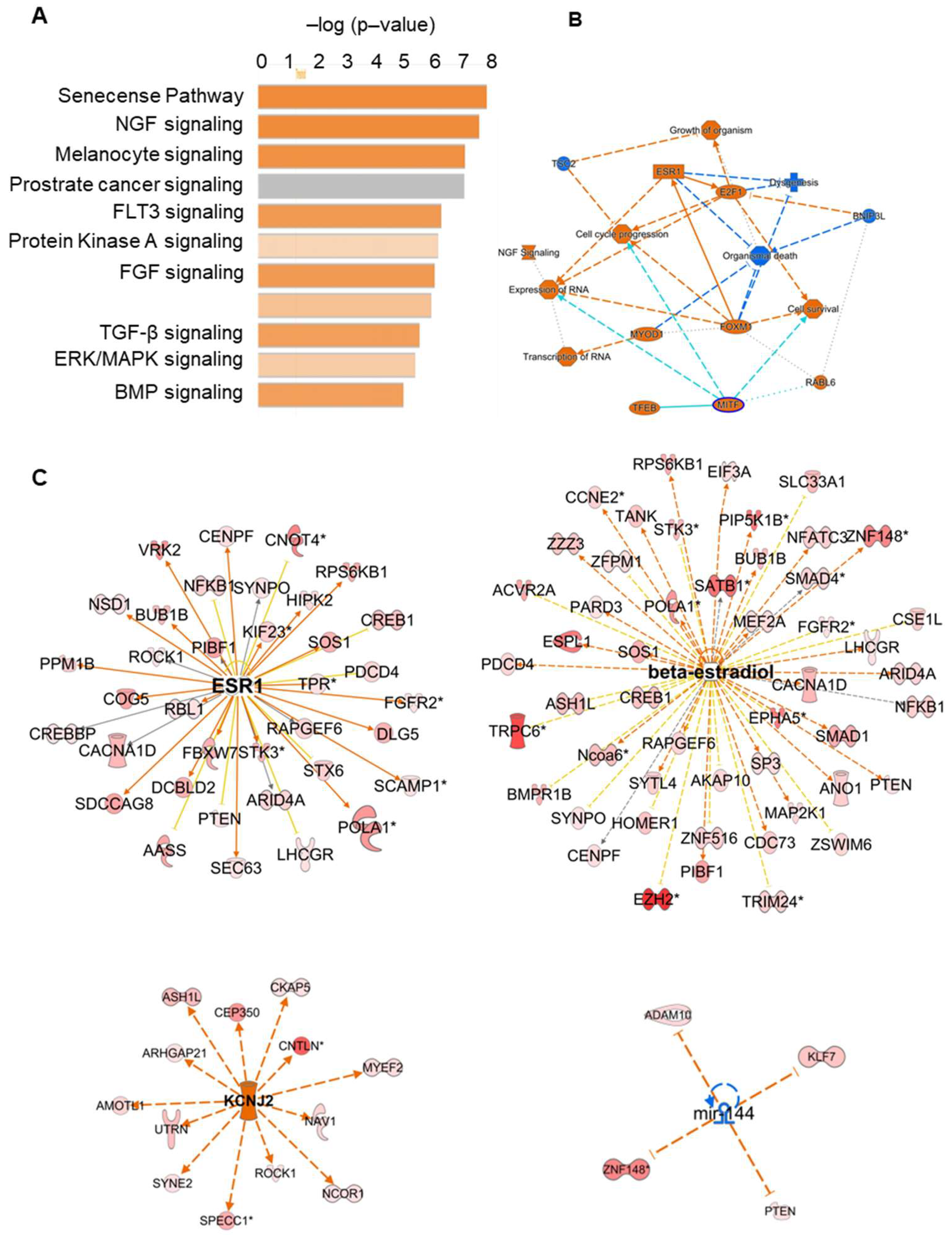
2. Results
2.1. Circular RNA Isoforms in Mural Granulosa Cells
2.2. circRNA vs. Its Linear RNA Form in Granulosa Cells before (0-h) and after (4-h) LH/hCG Surge
2.3. Differentially Expressed circRNA in Mural Granulosa Cells before and after the LH/hCG Surge
2.4. miRNA and Protein Interaction Network of Selected circRNA
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Isolation of RNA from Mural Granulosa Cells
4.3. RNA Sequencing and circRNA Expression Analyses
4.4. Pathway Analyses
4.5. miRNA and Protein Interaction Prediction
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gougeon, A. Human ovarian follicular development: From activation of resting follicles to preovulatory maturation. Ann. d’Endocrinol. 2010, 71, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Salzman, J.; Chen, R.E.; Olsen, M.N.; Wang, P.L.; Brown, P.O. Cell-type specific features of circular RNA expression. PLoS Genet. 2013, 9, e1003777. [Google Scholar] [CrossRef]
- Zhao, X.; Cai, Y.; Xu, J. Circular RNAs: Biogenesis, Mechanism, and Function in Human Cancers. Int. J. Mol. Sci. 2019, 20, 3926. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, C.; Sun, H.; Wang, J.; Liang, Y.; Wang, Y.; Wong, G. The bioinformatics toolbox for circRNA discovery and analysis. Brief. Bioinform. 2021, 22, 1706–1728. [Google Scholar] [CrossRef]
- Chen, N.; Zhao, G.; Yan, X.; Lv, Z.; Yin, H.; Zhang, S.; Song, W.; Li, X.; Li, L.; Du, Z.; et al. A novel FLI1 exonic circular RNA promotes metastasis in breast cancer by coordinately regulating TET1 and DNMT1. Genome Biol. 2018, 19, 218. [Google Scholar] [CrossRef]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Faculty Opinions recommendation of Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.-O.; Chen, T.; Xiang, J.-F.; Yin, Q.-F.; Xing, Y.-H.; Zhu, S.; Yang, L.; Chen, L.-L. Circular Intronic Long Noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef]
- Lei, M.; Zheng, G.; Ning, Q.; Zheng, J.; Dong, D. Translation and functional roles of circular RNAs in human cancer. Mol. Cancer 2020, 19, 30. [Google Scholar] [CrossRef]
- Yu, C.-Y.; Kuo, H.-C. The emerging roles and functions of circular RNAs and their generation. J. Biomed. Sci. 2019, 26, 29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, T.; Xiao, J. Circular RNAs: Promising Biomarkers for Human Diseases. Ebiomedicine 2018, 34, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Fang, L.; Yang, Q.; Hibberd, S.; Du, W.W.; Wu, N.; Yang, B.B. Posttranscriptional regulation of AKT by circular RNA angiomotin-like 1 mediates chemoresistance against paclitaxel in breast cancer cells. Aging 2019, 11, 11369–11381. [Google Scholar] [CrossRef] [PubMed]
- Santer, L.; Bär, C.; Thum, T. Circular RNAs: A Novel Class of Functional RNA Molecules with a Therapeutic Perspective. Mol. Ther. 2019, 27, 1350–1363. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Mang, G.; Wu, J.; Sun, P.; Li, T.; Zhang, H.; Wang, N.; Tong, Z.; Wang, W.; Zheng, Y.; et al. Circular RNA circSnx5 Controls Immunogenicity of Dendritic Cells through the miR-544/SOCS1 Axis and PU.1 Activity Regulation. Mol. Ther. 2020, 28, 2503–2518. [Google Scholar] [CrossRef]
- Zhao, W.; Cui, Y.; Liu, L.; Qi, X.; Liu, J.; Ma, S.; Hu, X.; Zhang, Z.; Wang, Y.; Li, H.; et al. Splicing factor derived circular RNA circUHRF1 accelerates oral squamous cell carcinoma tumorigenesis via feedback loop. Cell Death Differ. 2020, 27, 919–933. [Google Scholar] [CrossRef]
- Hu, H.; Xi, J.; Zhou, B.; Zhang, J.; Li, Z.; Liu, Z.; Jia, Q. Ovarian Circular RNAs Associated with High and Low Fertility in Large White Sows during the Follicular and Luteal Phases of the Estrous Cycle. Animals 2020, 10, 696. [Google Scholar] [CrossRef]
- Gao, Q.; Wang, T.; Pan, L.; Qian, C.; Wang, J.; Xin, Q.; Liu, Y.; Zhang, Z.; Xu, Y.; He, X.; et al. Circular RNAs: Novel potential regulators in embryogenesis, female infertility, and pregnancy-related diseases. J. Cell. Physiol. 2021, 236, 7223–7241. [Google Scholar] [CrossRef]
- Zhou, X.-Y.; Li, Y.; Zhang, J.; Liu, Y.-D.; Zhe, J.; Zhang, Q.-Y.; Chen, Y.-X.; Chen, X.; Chen, S.-L. Expression profiles of circular RNA in granulosa cells from women with biochemical premature ovarian insufficiency. Epigenomics 2020, 12, 319–332. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, J.; Lai, M.; Li, J.; Zhan, J.; Wen, Q.; Ma, H. Circular RNA expression profiling of granulosa cells in women of reproductive age with polycystic ovary syndrome. Arch. Gynecol. Obstet. 2019, 300, 431–440. [Google Scholar] [CrossRef]
- Dudekula, D.B.; Panda, A.C.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Gorospe, M. CircInteractome: A web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016, 13, 34–42. [Google Scholar] [CrossRef]
- Richards, J.S.; Pangas, S.A. The ovary: Basic biology and clinical implications. J. Clin. Investig. 2010, 120, 963–972. [Google Scholar] [CrossRef]
- Przygrodzka, E.; Plewes, M.R.; Davis, J.S. Luteinizing Hormone Regulation of Inter-Organelle Communication and Fate of the Corpus Luteum. Int. J. Mol. Sci. 2021, 22, 9972. [Google Scholar] [CrossRef]
- Kim, J.; Bagchi, I.C.; Bagchi, M.K. Control of ovulation in mice by progesterone receptor-regulated gene networks. Mol. Hum. Reprod. 2009, 15, 821–828. [Google Scholar] [CrossRef]
- Chakravarthi, V.P.; Ratri, A.; Masumi, S.; Borosha, S.; Ghosh, S.; Christenson, L.K.; Roby, K.F.; Wolfe, M.W.; Rumi, M.K. Granulosa cell genes that regulate ovarian follicle development beyond the antral stage: The role of estrogen receptor β. Mol. Cell. Endocrinol. 2021, 528, 111212. [Google Scholar] [CrossRef]
- Abbassi, L.; El-Hayek, S.; Carvalho, K.F.; Wang, W.; Yang, Q.; Granados-Aparici, S.; Mondadori, R.; Bordignon, V.; Clarke, H.J. Epidermal growth factor receptor signaling uncouples germ cells from the somatic follicular compartment at ovulation. Nat. Commun. 2021, 12, 1438. [Google Scholar] [CrossRef]
- Zhou, J.; Lei, B.; Li, H.; Zhu, L.; Wang, L.; Tao, H.; Mei, S.; Li, F. MicroRNA-144 is regulated by CP2 and decreases COX-2 expression and PGE2 production in mouse ovarian granulosa cells. Cell Death Dis. 2017, 8, e2597. [Google Scholar] [CrossRef]
- Rodriguez, K.F.; Couse, J.F.; Jayes, F.L.; Hamilton, K.J.; Burns, K.A.; Taniguchi, F.; Korach, K.S. Insufficient luteinizing hormone-induced intracellular signaling disrupts ovulation in preovulatory follicles lacking estrogen receptor-β. Endocrinology 2010, 151, 2826–2834. [Google Scholar] [CrossRef]
- Lykke-Andersen, K.; Gilchrist, M.J.; Grabarek, J.B.; Das, P.; Miska, E.; Zernicka-Goetz, M. Maternal Argonaute 2 Is Essential for Early Mouse Development at the Maternal-Zygotic Transition. Mol. Biol. Cell 2008, 19, 4383–4392. [Google Scholar] [CrossRef]
- Fowler, D.J.; Nicolaides, K.H.; Miell, J.P. Insulin-like growth factor binding protein-1 (IGFBP-1): A multifunctional role in the human female reproductive tract. Hum. Reprod. Up. 2000, 6, 495–504. [Google Scholar] [CrossRef]
- Rajasingh, J. The many facets of RNA-binding protein HuR. Trends Cardiovasc. Med. 2015, 25, 684–686. [Google Scholar] [CrossRef]
- Katsanou, V.; Milatos, S.; Yiakouvaki, A.; Sgantzis, N.; Kotsoni, A.; Alexiou, M.; Harokopos, V.; Aidinis, V.; Hemberger, M.; Kontoyiannis, D.L. The RNA-binding protein Elavl1/HuR is essential for placental branching morphogenesis and embryonic development. Mol. Cell Biol. 2009, 29, 2762–2776. [Google Scholar] [CrossRef]
- Valcárcel, J.; Gebauer, F. Post-transcriptional regulation: The dawn of PTB. Curr. Biol. 1997, 7, R705–R708. [Google Scholar] [CrossRef]
- Kamma, H.; Portman, D.S.; Dreyfuss, G. Cell Type-Specific Expression of hnRNP Proteins. Exp. Cell Res. 1995, 221, 187–196. [Google Scholar] [CrossRef]
- Lu, C.; Lin, L.; Tan, H.; Wu, H.; Sherman, S.L.; Gao, F.; Jin, P.; Chen, D. Fragile X premutation RNA is sufficient to cause primary ovarian insufficiency in mice. Hum. Mol. Genet. 2012, 21, 5039–5047. [Google Scholar] [CrossRef]
- Chen, E.; Joseph, S. Fragile X mental retardation protein: A paradigm for translational control by RNA-binding proteins. Biochimie 2015, 114, 147–154. [Google Scholar] [CrossRef]
- Rios, C.; Warren, D.; Olson, B.; Abbott, A.L. Functional analysis of microRNA pathway genes in the somatic gonad and germ cells during ovulation in C. elegans. Dev. Biol. 2017, 426, 115–125. [Google Scholar] [CrossRef]
- Chen, J.; Liu, W. Lin28a induced mitochondrial dysfunction in human granulosa cells via suppressing LARS2 expression. Cell. Signal. 2023, 103, 110536. [Google Scholar] [CrossRef]
- Mu, H.; Cai, S.; Wang, X.; Li, H.; Zhang, L.; Li, H.; Xiang, W. RNA binding protein IGF2BP1 meditates oxidative stress-induced granulosa cell dysfunction by regulating MDM2 mRNA stability in an m6A-dependent manner. Redox Biol. 2022, 57, 102492. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, Q.; Chen, T.; Zhao, X.; Xin, Z.; Yang, X. Differentially expressed IGF2BP2 in ovarian disorders: Strongly associates with alternative splicing regulation in human granulosa cells. Res. Sq. 2021. preprint. [Google Scholar]
- Russell, D.L.; Ochsner, S.A.; Hsieh, M.; Mulders, S.; Richards, J.S. Hormone-Regulated Expression and Localization of Versican in the Rodent Ovary. Endocrinology 2003, 144, 1020–1031. [Google Scholar] [CrossRef] [PubMed]
- Tsafriri, A.; Chun, S.-Y.; Zhang, R.; Hsueh, A.; Conti, M. Oocyte Maturation Involves Compartmentalization and Opposing Changes of cAMP Levels in Follicular Somatic and Germ Cells: Studies Using Selective Phosphodiesterase Inhibitors. Dev. Biol. 1996, 178, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.C.; Dostie, J.; Diem, M.D.; Feng, W.; Mann, M.; Rappsilber, J.; Dreyfuss, G. eIF4A3 is a novel component of the exon junction complex. RNA 2004, 10, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Macklon, N.S.; Fauser, B.C. Follicle-Stimulating Hormone and Advanced Follicle Development in the Human. Arch. Med. Res. 2001, 32, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Shirafuta, Y.; Tamura, I.; Ohkawa, Y.; Maekawa, R.; Doi-Tanaka, Y.; Takagi, H.; Mihara, Y.; Shinagawa, M.; Taketani, T.; Sato, S.; et al. Integrated Analysis of Transcriptome and Histone Modifications in Granulosa Cells During Ovulation in Female Mice. Endocrinology 2021, 162, bqab128. [Google Scholar] [CrossRef]
- Myers, M.; Britt, K.L.; Wreford, N.G.M.; Ebling, F.J.P.; Kerr, J.B. Methods for quantifying follicular numbers within the mouse ovary. Reproduction 2004, 127, 569–580. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 1 October 2018).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Ma, X.-K.; Xue, W.; Chen, L.-L.; Yang, L. CIRCexplorer pipelines for circRNA annotation and quantification from non-polyadenylated RNA-seq datasets. Methods 2021, 196, 3–10. [Google Scholar] [CrossRef]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | circRNA Chromosomal Region | Corresponding Human circRNA | circRNA Interacting Factors | |
| miRNA | Protein | |||
| Gpr98 | 13:81270744-81283472 | hsa_circ_0073295 | hsa-miR:1252 (7); 197 (7) | FUS (6); HuR (15); IGF2BP1 (3); IGF2BP2 (6); IGF2BP3 (6); LIN28A (12). |
| Crim1 | 17:78344350-78355177 | hsa_circ_0002938 | AGO2 (11); EIF4A3 (8); PTB (3). | |
| Slc23a2 | 2:132089096-132094240 | hsa_circ_0059360 | AGO2 (3); EIF4A3 (28); FMRP (3); PTB (8). | |
| Vcan | 13:89721567-89731534 | hsa_circ_0073234 | hsa-miR: 326 (3); 330-5p (3); 431 (3); 494 (3); 587 (7); 622 (4); 640 (5); 767-3p (4). | AGO1 (3); CAPRIN1 (3); DGCR8 (5); EWSR1 (6); FMRP (71); FUS (8); HuR (43); IGF2BP1 (41); IGF2BP2 (38); IGF2BP3 (43); LIN28A (33); METTL3 (1); ZC3H7B (6). |
| Ano4 | 10:89016075-89035181 | hsa_circ_0027873 | EIF4A3 (11). | |
| Chst15 | 7:132262683-132271001 | hsa_circ_0000264 | AGO2 (6); EIF4A3 (12); FMRP (3); IGF2BP1 (3); PTB (5). | |
| Vcl | 14:20982574-20987118 | EIF4A3 (3); FMRP (7); LIN28A (3); PTB (3). | ||
| Tlk2 | 11:105240361-105241661 | hsa_circ_0008371 | EIF4A3 (3). | |
| Ptpn4 | 1:119715947-119773271 | hsa_circ_0117171 | hsa-miR: 197 (3); 532-3p (3). | AGO2 (5); EIF4A3 (7). |
| Slc7a11 | 3:50417988-50438915 | hsa_circ_0070981 | AGO2 (4); EIF4A3 (9); FUS (4); HuR (9); PTB (4). | |
| Clk1 | 1:58417013-58417383 | hsa_circ_0057725 | AGO2 (6); EIF4A3 (7). | |
| Zdhhc23 | 16:43973468-43974166 | hsa_circ_0066837 | EIF4A3 (16). | |
| Gm20459,Palm2 | 4:57638010-57648120 | hsa_circ_0137684 | AGO1 (7); AGO2 (30); DGCR8 (10); EIF4A3 (51); EWSR1 (4); FMRP (95); FXR2 (3); HuR (50); IGF2BP1 (20); IGF2BP2 (19); IGF2BP3 (25); LIN28A (6); LIN28B (5); PTB (4). | |
| Abca1 | 4:53127597-53133015 | hsa_circ_0005443 | EIF4A3 (3). | |
| Robo1 | 16:72742126-72742452 | hsa_circ_0004788 | AGO2 (4); DGCR8 (3); PTB (3). | |
| Smchd1 | 17:71436706-71448846 | hsa_circ_0046712 | hsa-miR:1183 (3); 1243 (3); 1245 (1); 1248 (4); 1305 (4); 331-5p (3); 513a-3p (5); 607 (5). | AGO2 (26); EIF4A3 (80); EWSR1 (8); FMRP (8); HuR (10); IGF2BP3 (4); LIN28A (3). |
| Lrp6 | 6:134541652-134542045 | hsa_circ_0000378 | EIF4A3 (5); FMRP (4). | |
| Eprs | 1:185386246-185397267 | hsa_circ_0007739 | AGO2 (6); FMRP (5); HuR (3). | |
| Eps15l1 | 8:72367904-72380306 | hsa_circ_0049885 | hsa-miR:1231 (3). | FMRP (3). |
| hsa_circ_0137685 | EIF4A312 (12). | |||
| Mast4,Gm6211 | 13:103140163-103171556 | hsa_circ_0129416 | hsa-miR:1178 (3); 1236 (3); 1243 (3); 1245 (3); 1256 (4); 1270 (5); 1299 (3); 1324 (4); 142-5p (4); 145 (3); 155 (3); 186 (6); 194 (5); 31 (4); 323-3p (3); 330-3p (3); 335 (4); 338-5p (4); 361-3p (3); 377 (3); 432 (5); 433 (3); 488 (4); 495 (3); 498 (3); 513a-3p (3); 516b (4); 532-3p (3); 545 (5); 548b-3p (6); 548c-3p (7); 548p (7); 549 (4); 555 (3); 556-3p (3); 569 (3); 576-5p (5); 578 (5); 580 (3); 587 (3); 605 (4); 607 (4); 620 (5); 623 (5); 633 (3); 646 (3); 649 (4); 668 (3); 877 (3); 888 (3). | |
| Sema3c | 5:17678312-17682083 | hsa_circ_0003634 | hsa-miR:503 (3). | AGO2 (7); IGF2BP3 (3); EIF4A3 (14); HuR (5); PTB (5). |
| Cnot4 | 6:35077981-35080226 | hsa_circ_0003629 | AGO2 (6); HuR (3). | |
| Atrnl1 | 19:57751630-57777945 | hsa_circ_0092760 | hsa-miR:1238 (4); 409-3p (3); 515-5p (3); 518a-5p (3); 526b (3); 527 (3); 548p (5); 567 (3); 577 (4); 579 (6); 607 (7); 671-5p (3). | FUS (6); IGF2BP1 (3); IGF2BP2 (3); IGF2BP3 (7). |
| Dennd1b | 1:139040000-139062971 | hsa_circ_0006324 | . | AGO2 (4); EIF4A3 (6). |
| Ttll5 | 12:85863564-85879475 | hsa_circ_0032699 | EIF4A3 (15); PTB (3). | |
| Ywhae | 11:75751881-75759428 | hsa_circ_0041192 | AGO1 (24); AGO2 (92); AGO3 (4); C22ORF28 (8); CAPRIN1 (8); DGCR8 (5); EIF4A3 (20); FMRP (20); FUS (8); HuR (16); IGF2BP1 (16); IGF2BP2 (16); IGF2BP3 (11); LIN28A (11); LIN28B (6); METTL3 (2); MOV10 (3); PTB (8); SFRS1 (12); TIA1 (3); TIAL1 (4); TNRC6 (1); U2AF65 (3); ZC3H7B (10). | |
| Kif24 | 4:41400437-41414999 | hsa_circ_0138678 | hsa-miR: 1825 (3); hsa-miR-640 (3); hsa-miR-647 (3); hsa-miR-942 (3). | EIF4A3 (12); PTB (4). |
| Cul5 | 9:53642518-53642826 | hsa_circ_0024169 | EIF4A3 (11). | |
| Erbb2ip | 13:103866649-103889285 | hsa_circ_0001492 | EIF4A3 (3). | |
| Emg1 | 6:124705158-124705656 | hsa_circ_0025313 | AGO2 (9); EIF4A3 (10); FMRP (7); HuR (8); LIN28A (3). | |
| Vps41 | 13:18810432-18814293 | hsa_circ_0002808 | EIF4A3 (6). | |
| Mllt10 | 2:18122245-18126229 | hsa_circ_0017934 | hsa-miR:1200 (4); 494 (3); 604 (3); 647 (3). | AGO2 (7); EIF4A3 (25); HuR (8); IGF2BP2 (4); IGF2BP3 (4). |
| Pip4k2b | 11:97726705-97732755 | hsa_circ_0043379 | EIF4A3 (8); FMRP (4); HuR (5). | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chakravarthi, V.P.; Hung, W.-T.; Yellapu, N.K.; Gunewardena, S.; Christenson, L.K. LH/hCG Regulation of Circular RNA in Mural Granulosa Cells during the Periovulatory Period in Mice. Int. J. Mol. Sci. 2023, 24, 13078. https://doi.org/10.3390/ijms241713078
Chakravarthi VP, Hung W-T, Yellapu NK, Gunewardena S, Christenson LK. LH/hCG Regulation of Circular RNA in Mural Granulosa Cells during the Periovulatory Period in Mice. International Journal of Molecular Sciences. 2023; 24(17):13078. https://doi.org/10.3390/ijms241713078
Chicago/Turabian StyleChakravarthi, V. Praveen, Wei-Ting Hung, Nanda Kumar Yellapu, Sumedha Gunewardena, and Lane K. Christenson. 2023. "LH/hCG Regulation of Circular RNA in Mural Granulosa Cells during the Periovulatory Period in Mice" International Journal of Molecular Sciences 24, no. 17: 13078. https://doi.org/10.3390/ijms241713078