
The Effect of Aging on Nitric Oxide Production during Cerebral Ischemia and Reperfusion in Wistar Rats and Spontaneous Hypertensive Rats: An In Vivo Microdialysis Study
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. MABP
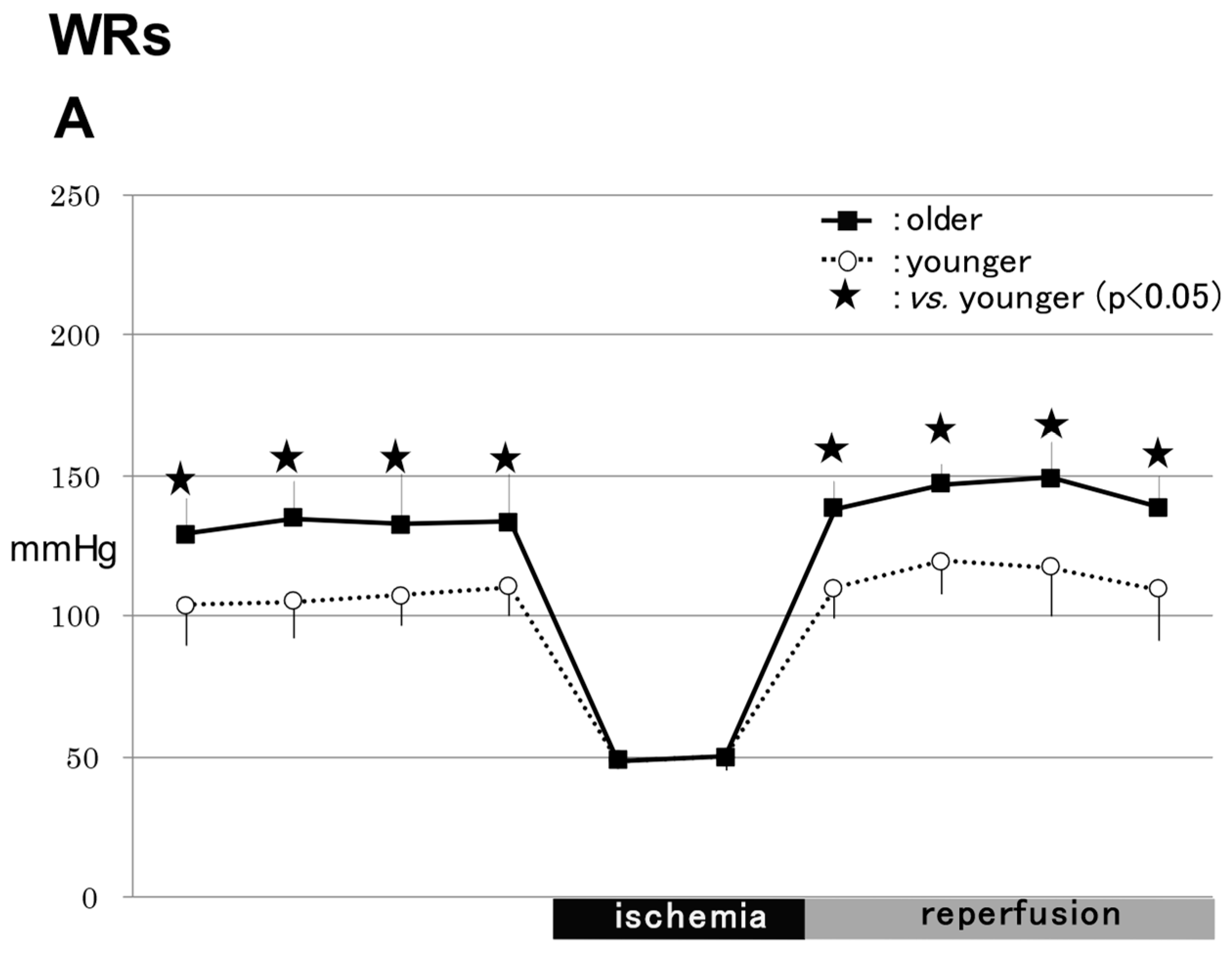
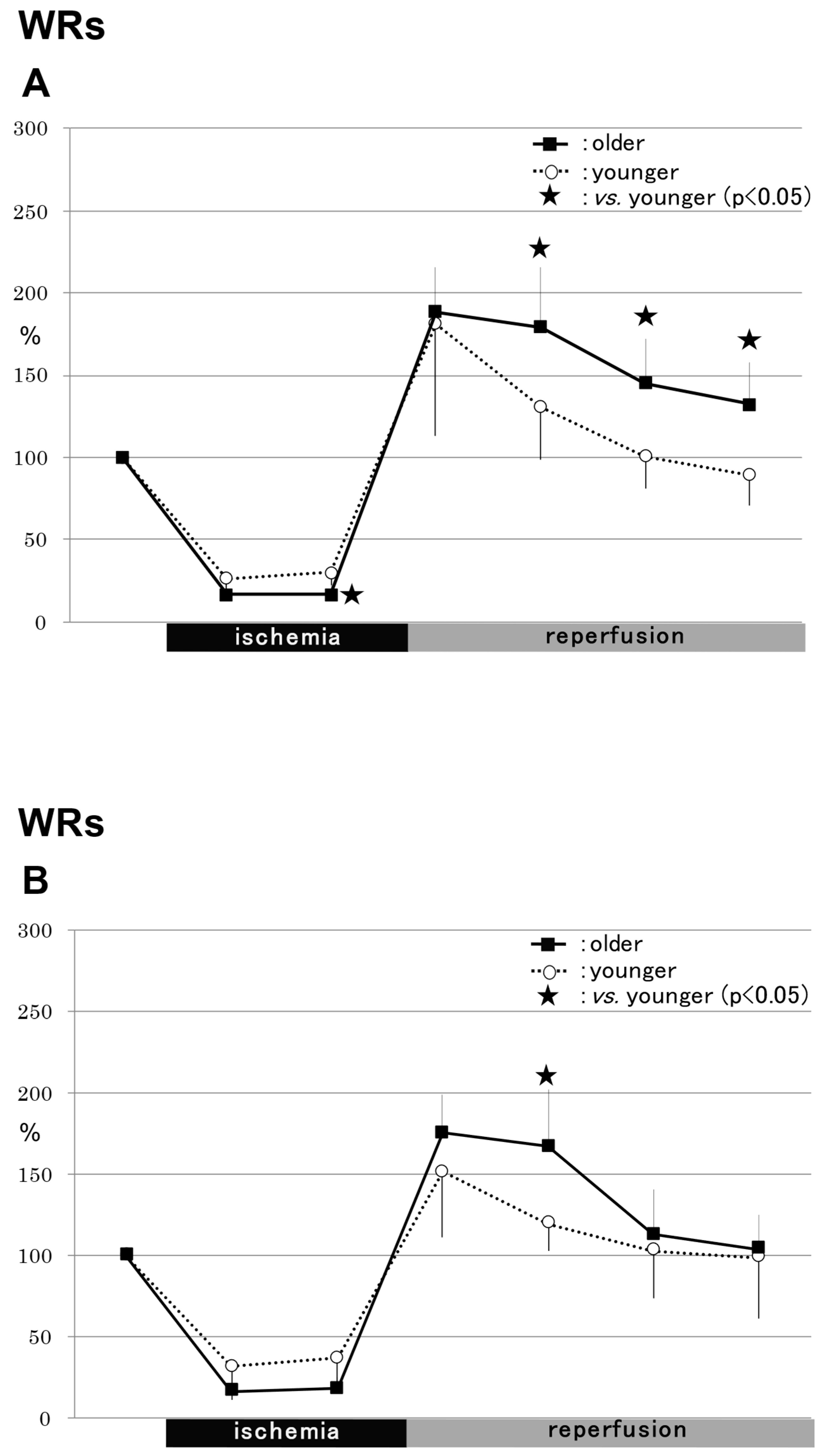
2.1.1. WRs
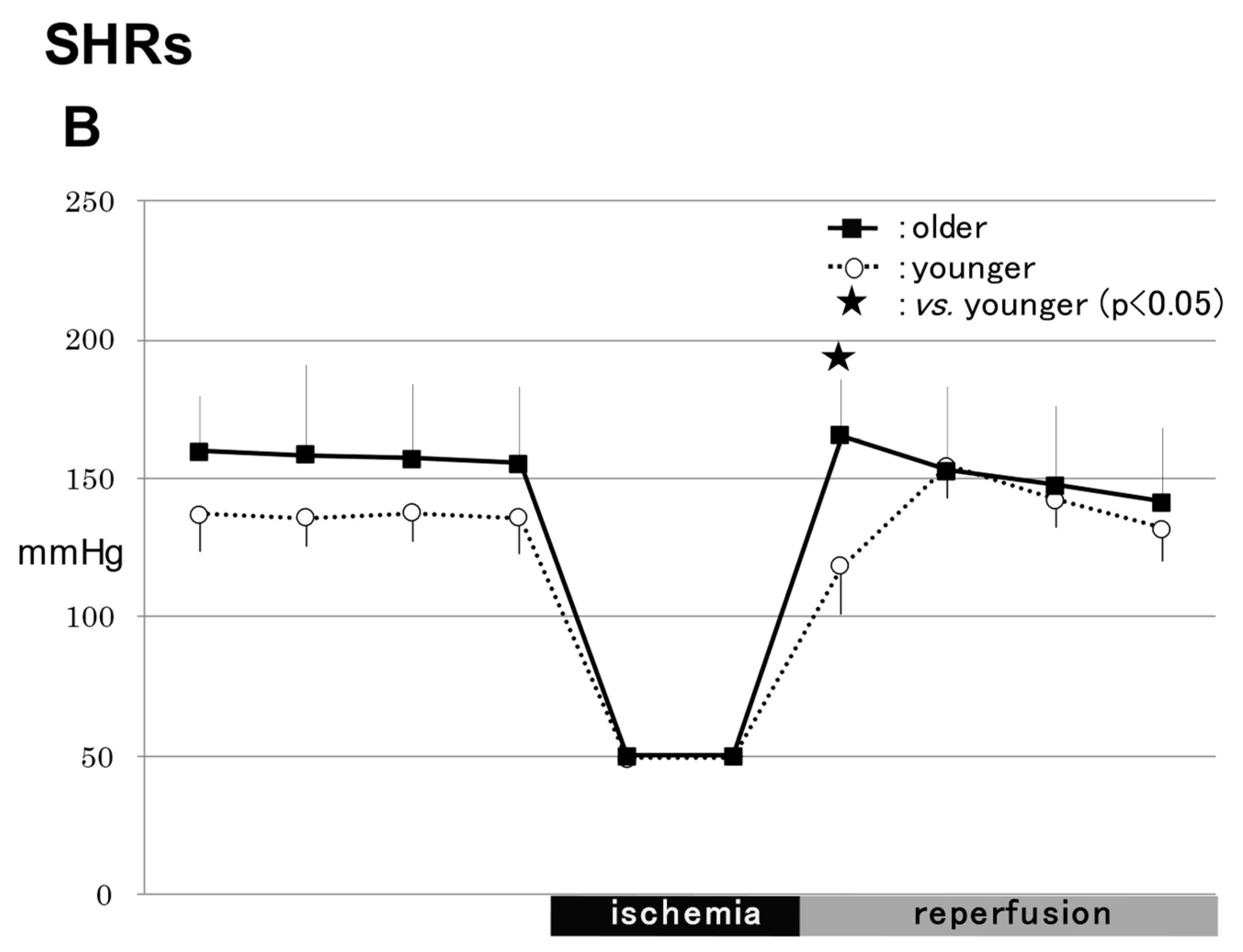
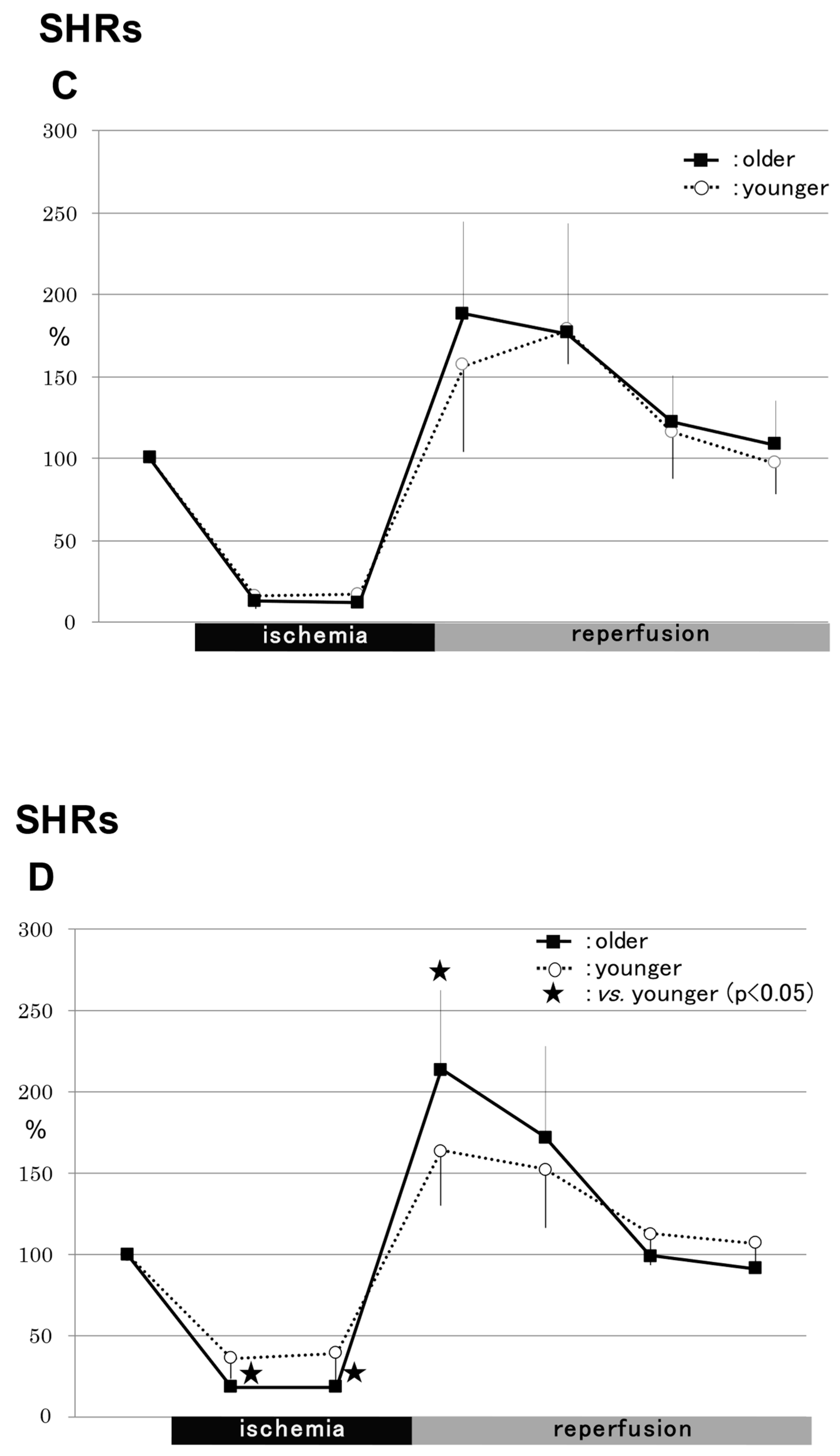
2.1.2. SHRs
2.1.3. Comparison of MABPs between WRs and SHRs
2.2. rCBF
2.2.1. WRs
2.2.2. SHRs
2.3. NO Metabolites
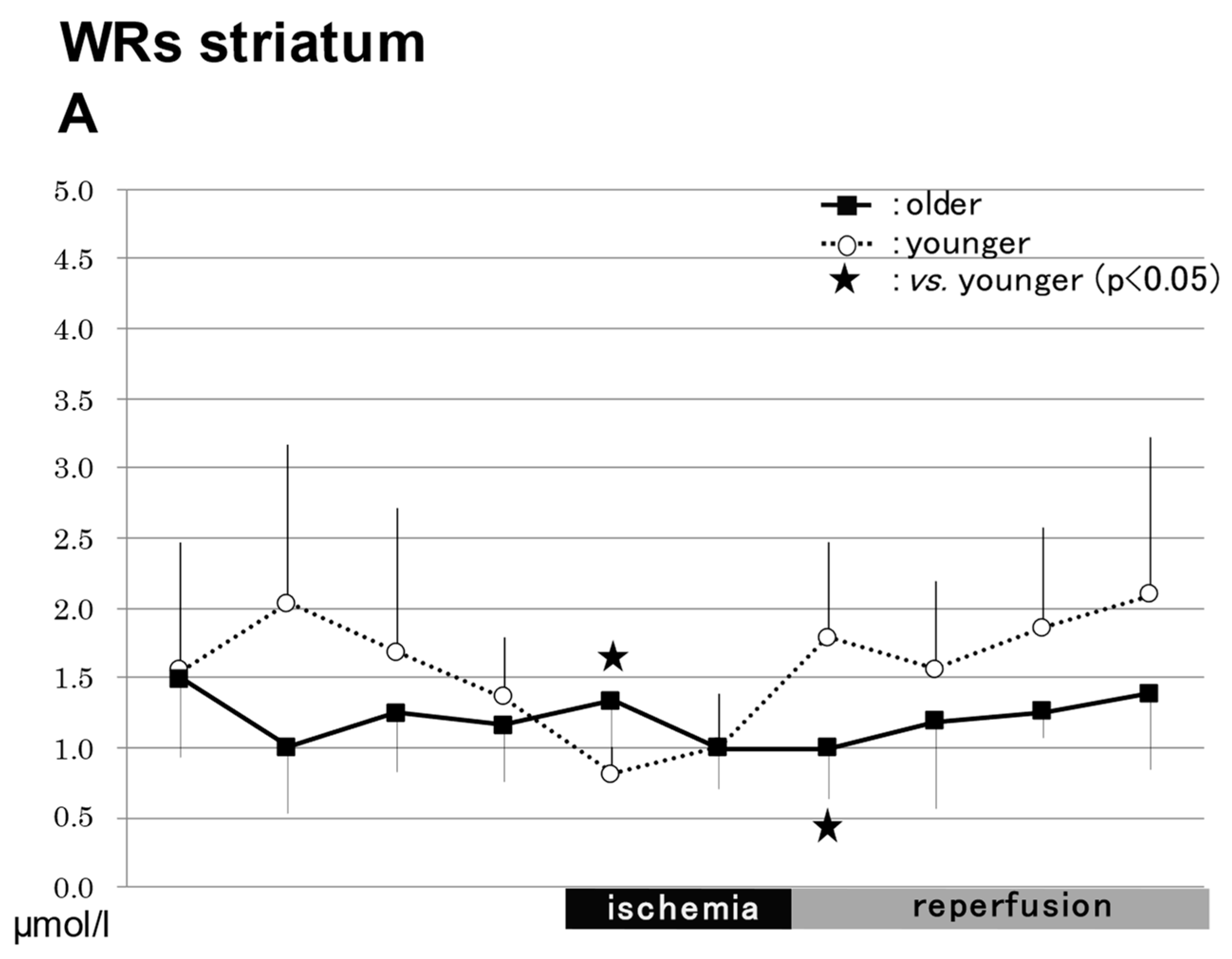
2.3.1. In the Striatum in WRs (Figure 3)
NO2−




NO3−
Total NO
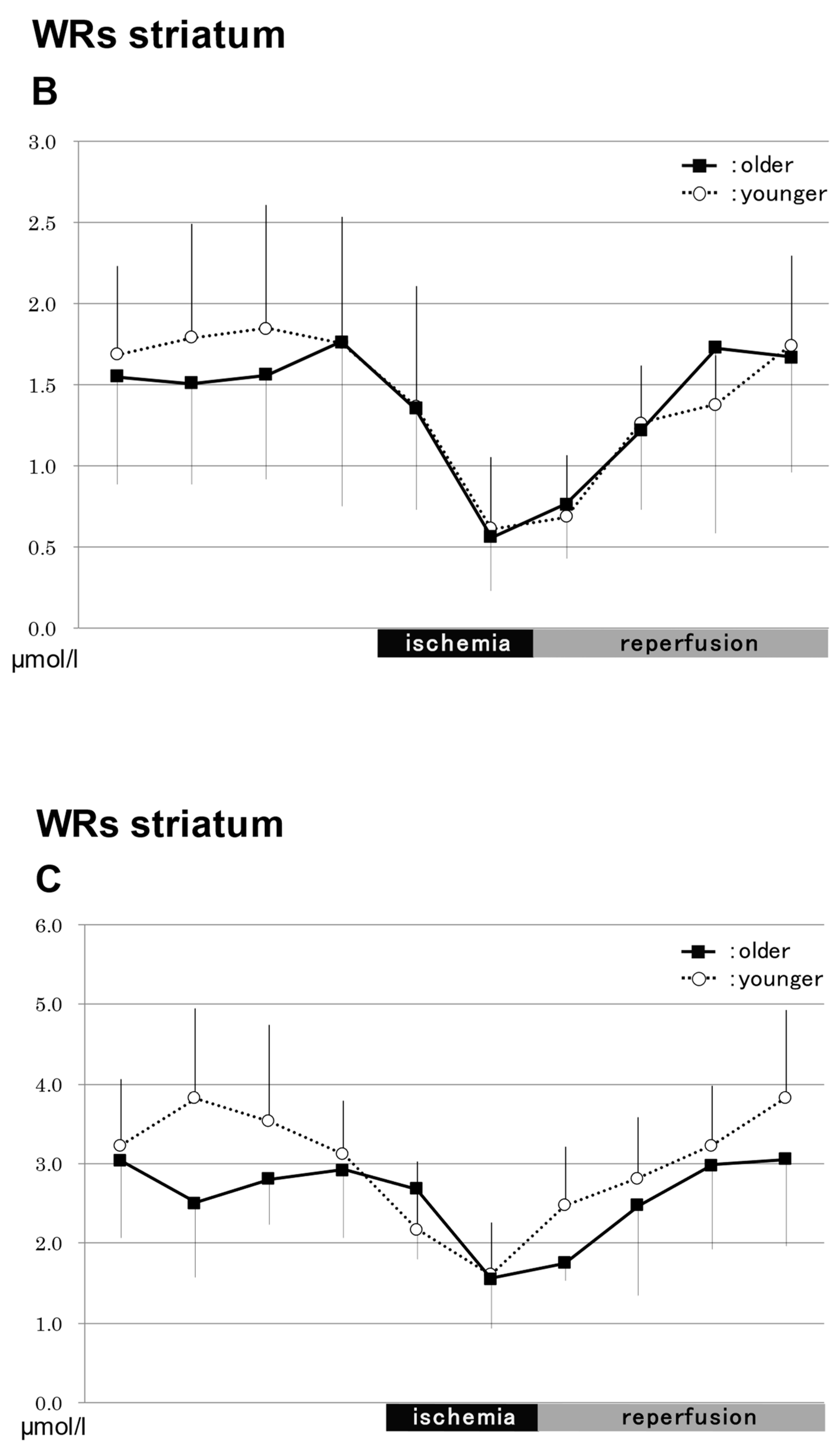
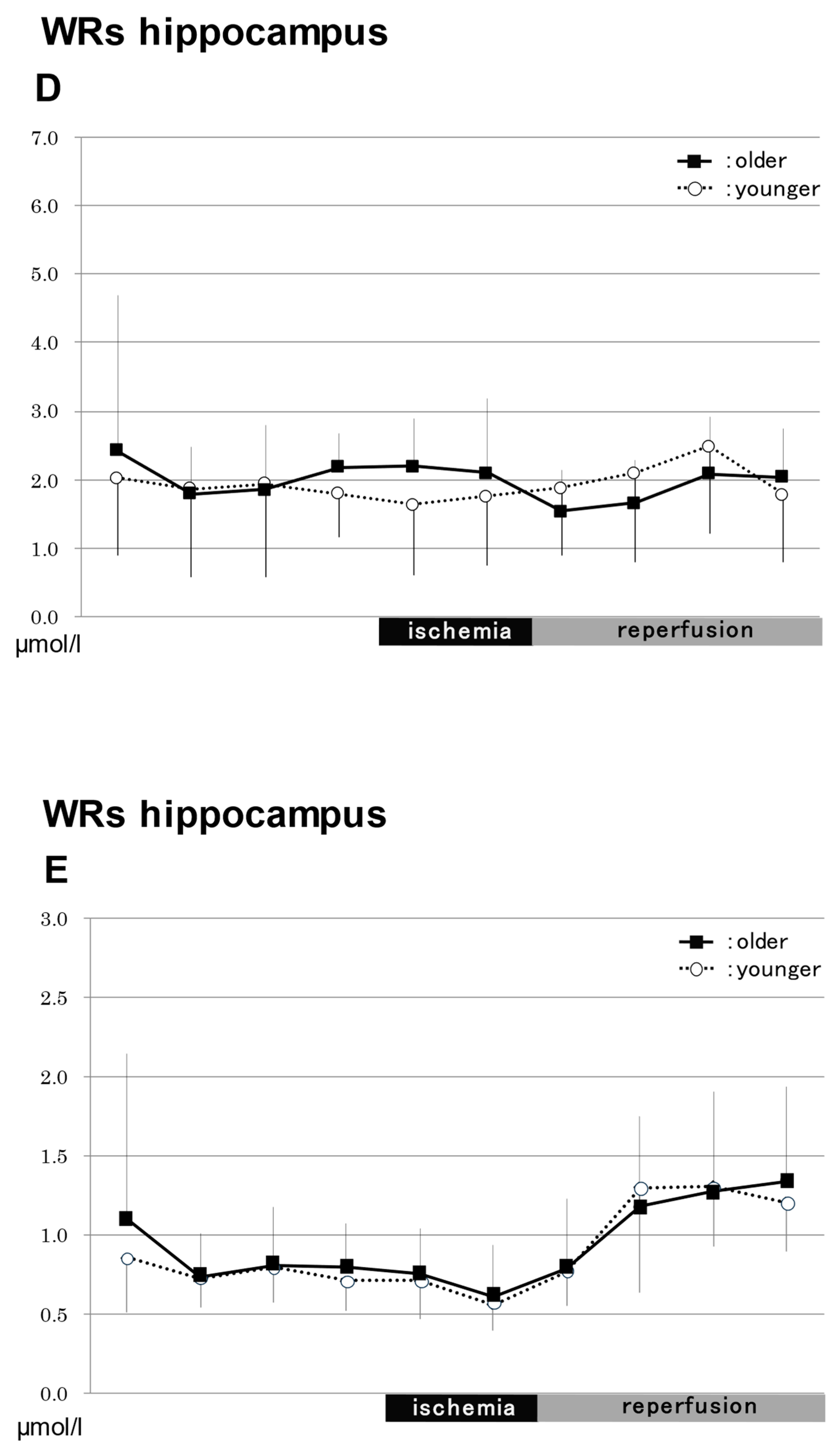
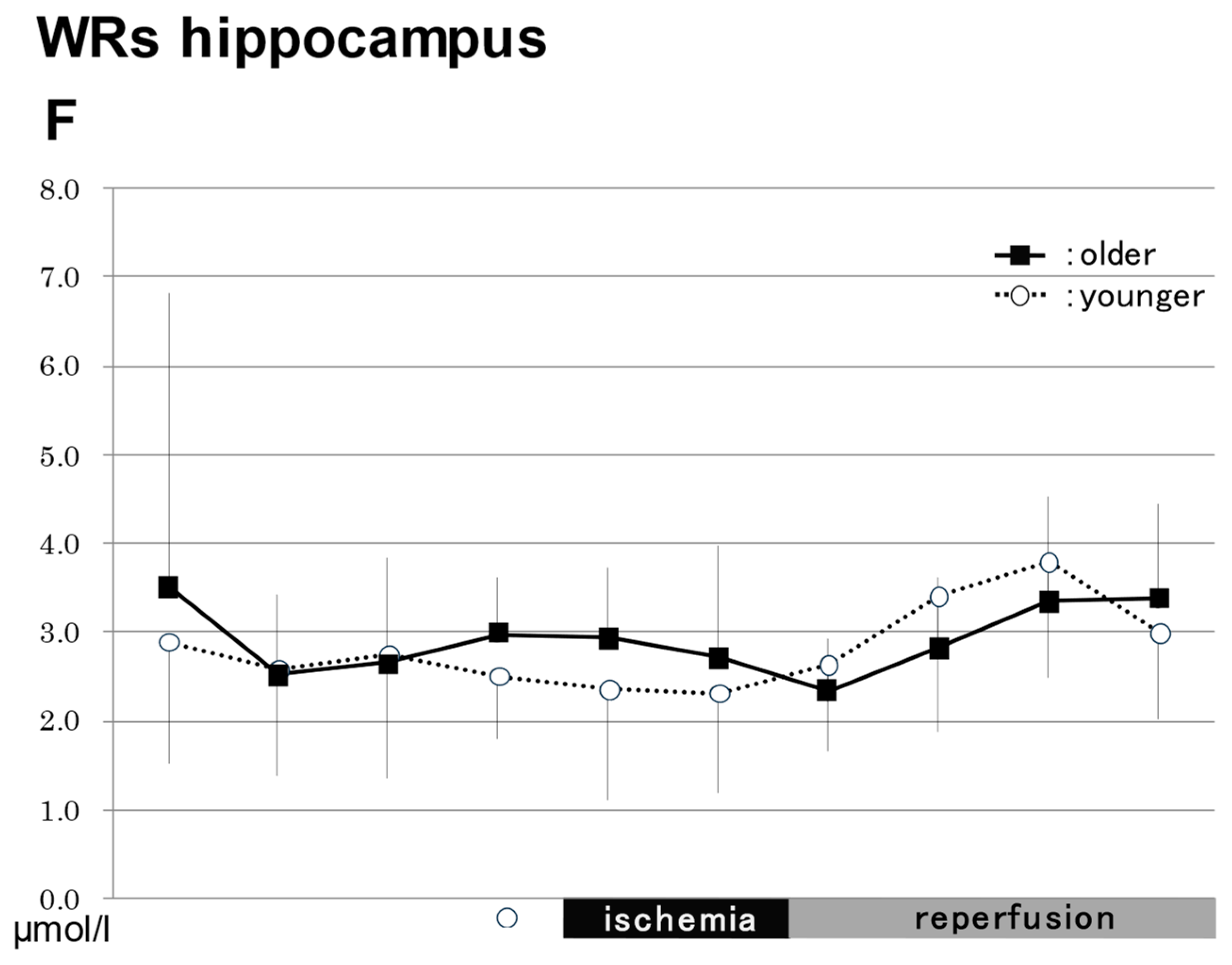
2.3.2. In the Hippocampus in WRs (Figure 3)
NO2−
NO3−
Total NO
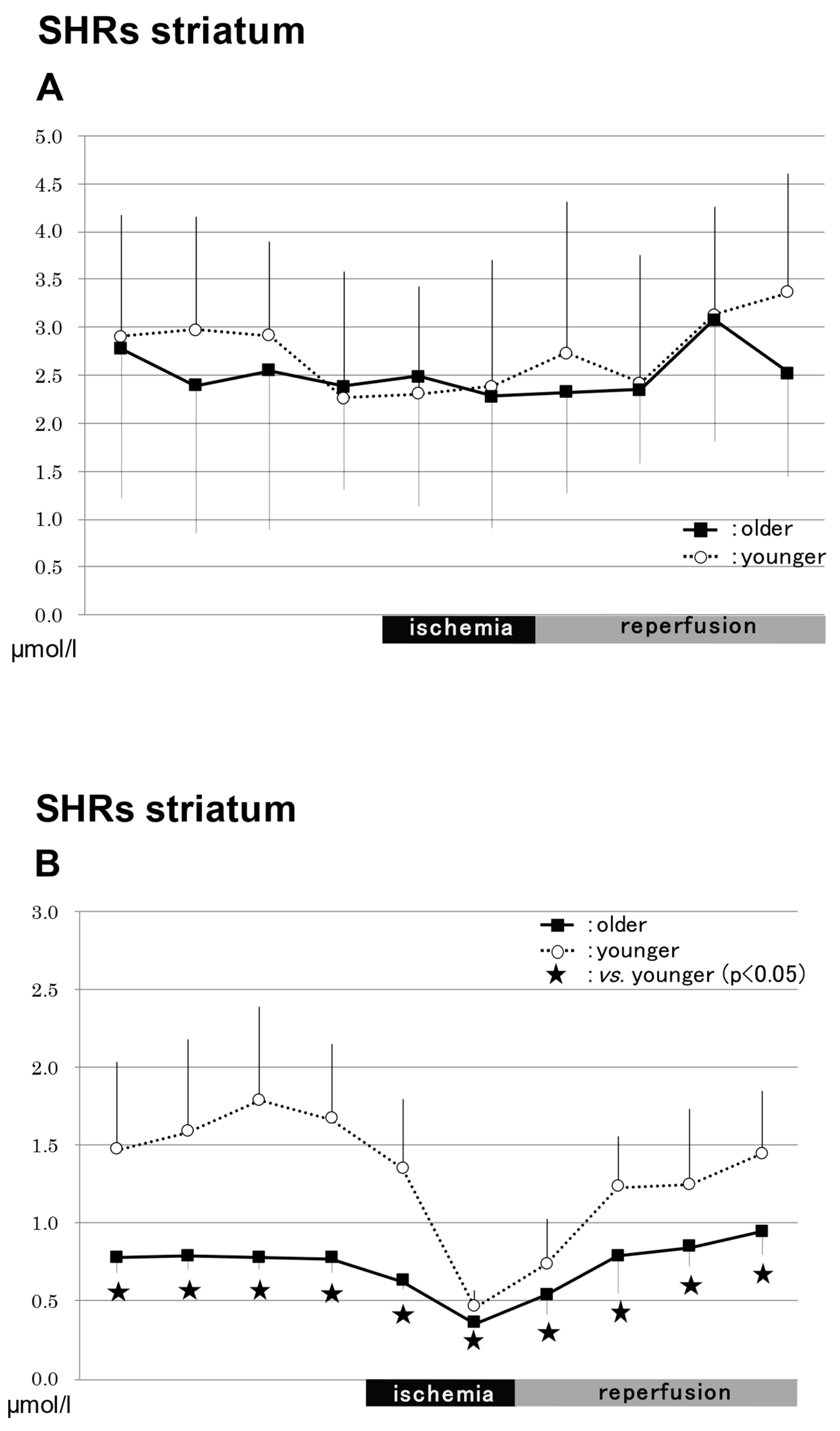
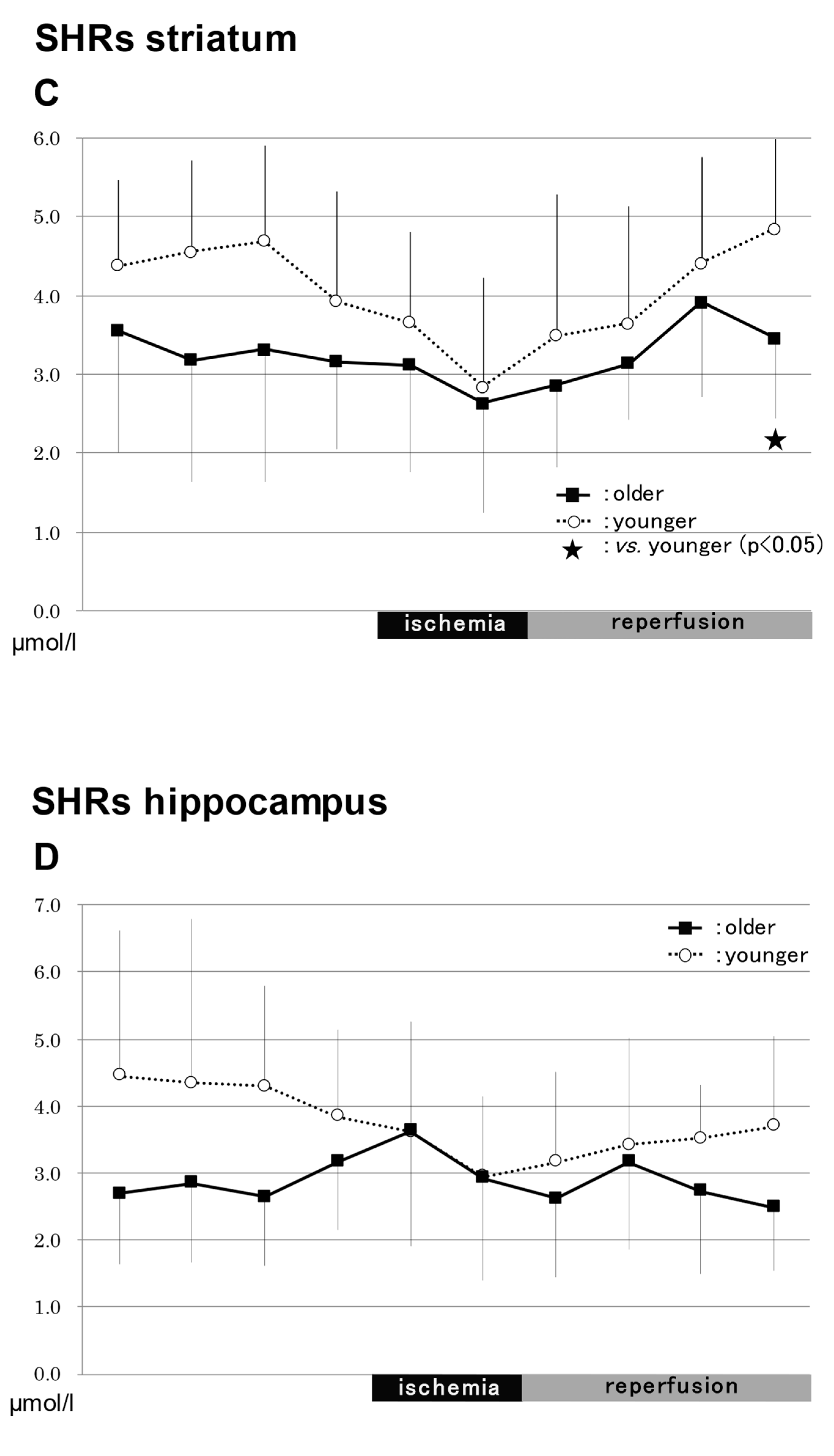
2.3.3. In the Striatum in SHRs (Figure 4)
NO2−



NO3−
Total NO
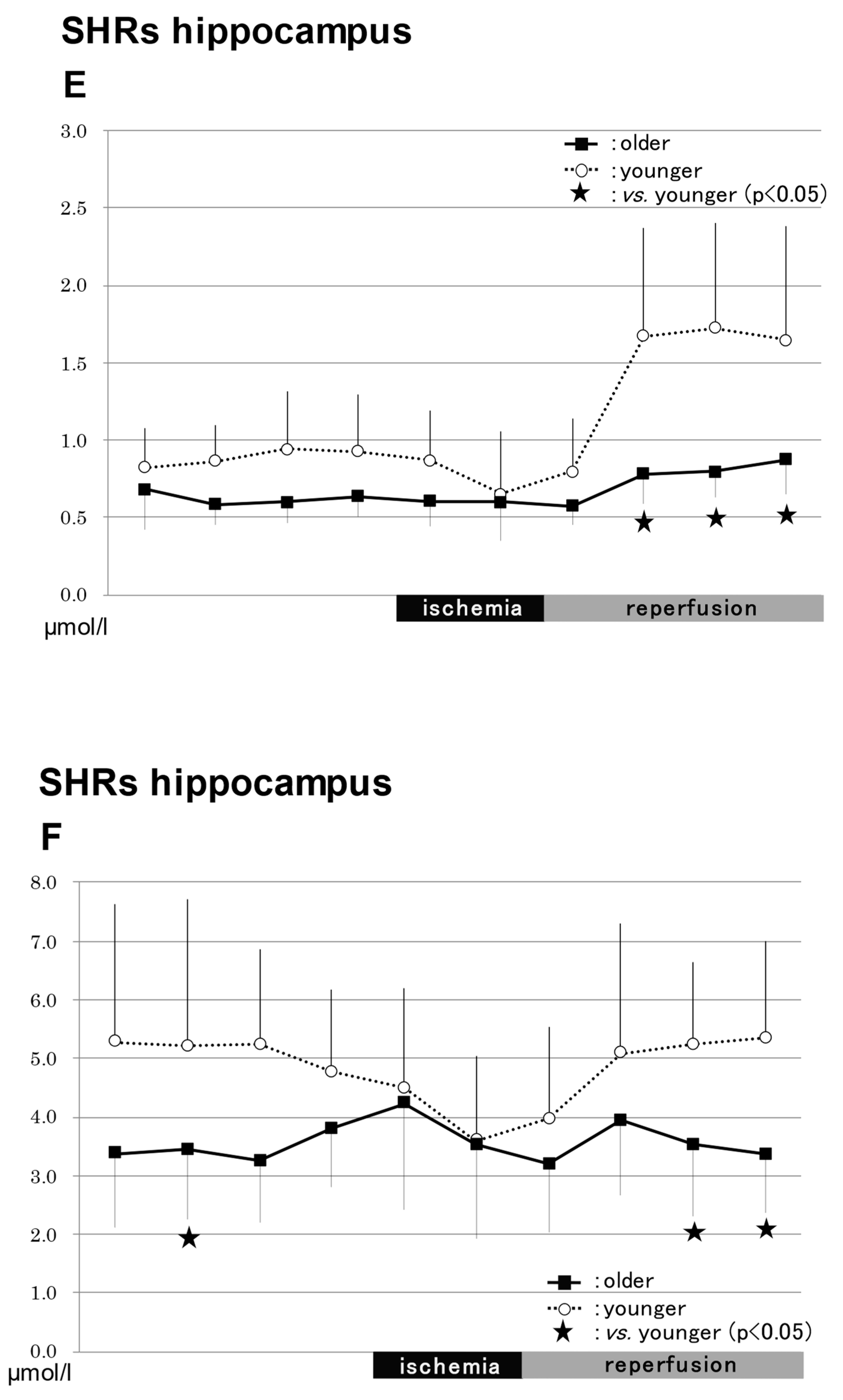
2.3.4. In the Hippocampus in SHRs (Figure 4)
NO2−
NO3−
Total NO
2.3.5. Comparison of Total NO between WRs and SHRs
3. Materials and Methods
3.1. Animal Groupings
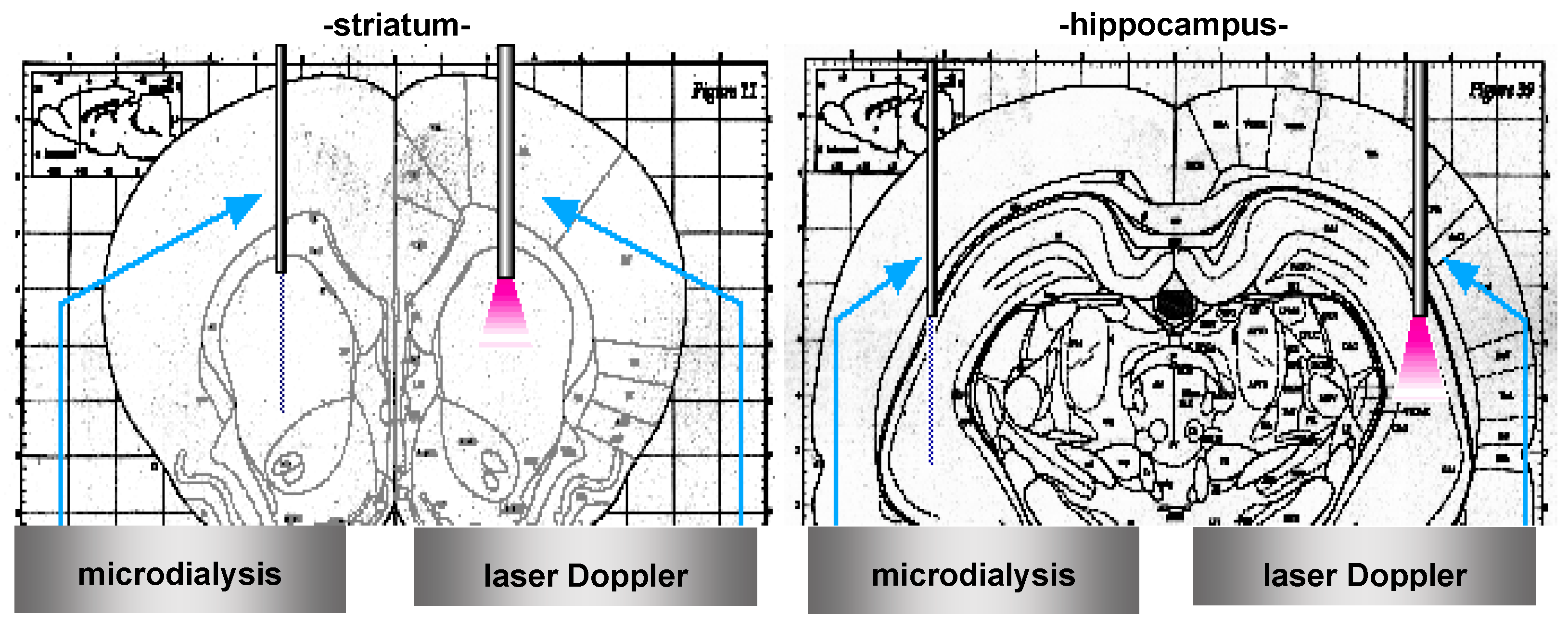
3.2. In Vivo Microdialysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garthwaite, J. Concepts of neural nitric oxide-mediated transmission. Eur. J. Neurosci. 2008, 27, 2783–2802. [Google Scholar] [CrossRef] [Green Version]
- Shibata, M.; Araki, N.; Hamada, J.; Sasaki, T.; Shimazu, K.; Fukuuchi, Y. Brain nitrite production during global ischemia and reperfusion: An in vivo microdialysis study. Brain Res. 1996, 734, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Ohkubo, T.; Asano, Y.; Hattori, K.; Shimazu, T.; Yamazato, M.; Nagoya, H.; Kato, Y.; Araki, N. Nitric Oxide Production during Cerebral Ischemia and Reperfusion in eNOS- and nNOS-Knockout Mice. Curr. Neurovascular Res. 2010, 7, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, H.; Ito, Y.; Kitabayashi, C.; Tanaka, A.; Nishioka, R.; Yamazato, M.; Ishizawa, K.; Nagai, T.; Hirayama, M.; Takahashi, K.; et al. Effects of Edaravone on Nitric Oxide, Hydroxyl Radicals and Neuronal Nitric Oxide Synthase During Cerebral Ischemia and Reperfusion in Mice. J. Stroke Cerebrovasc. Dis. 2020, 29, 104531. [Google Scholar] [CrossRef]
- Gratton, J.A.; Sauter, A.; Rudin, M.; Lees, K.R.; McColl, J.; Reid, J.L.; Dominiczak, A.F.; Macrae, I.M. Susceptibility to cerebral infarction in the stroke-prone spontaneously hypertensive rat is inherited as a dominant trait. Stroke 1998, 29, 690–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrucki, L.W.; Kalinowski, L.; Uracz, W.; Malinski, T. The protective role of nitic oxide in the brain ischemia. J. Physiol. Pharmaclol. 2000, 51, 695–703. [Google Scholar]
- Yamakawa, H.; Jezova, M.; Ando, H.; Saavedra, J.M. Normalization of endothelial and inducible nitric oxide synthase expression in brain microvessels of spontaneously hypertensive rats by angiotensin II AT1 receptor inhibition. J. Cereb. Blood Flow Metab. 2003, 23, 371–380. [Google Scholar] [CrossRef] [Green Version]
- Hojná, S.; Kuneš, J.; Zicha, J. Alterations of NO Synthase Isoforms in Brain and Kidney of Rats With Genetic and Salt Hypertension. Physiol. Res. 2010, 59, 997–1009. [Google Scholar] [CrossRef]
- Sverdlov, A.L.; Ngo, D.T.M.; Chan, W.P.A.; Chirkov, Y.Y.; Horowitz, J.D. Aging of nitric oxide system: Are we as old as our NO? J. Am. Heart Assoc. 2014, 3, e000973. [Google Scholar] [CrossRef] [Green Version]
- Shannon, O.M.; Clifford, T.; Seals, D.R.; Graighead, D.H. Nitic oxide, aging and aerobic exercise: Sedentary individuals to Master’s athletes. Nitric Oxide 2022, 125, 31–39. [Google Scholar] [CrossRef]
- Katsuta, T. Decreased Local Blood Flow in Young and Aged Spontaneously Hypertensive Rats. Fukuoka Acta Med. 1997, 88, 65–74. [Google Scholar] [PubMed]
- Kirsch, J.R.; Helfaer, M.A.; Blizzard, K.; Toung, T.J.; Traystman, R.J. Age-related cerebrovascular response to global ischemia in pigs. Am. J. Physiol. 1990, 259, H1551–H1558. [Google Scholar] [CrossRef]
- Furchgott, R.F.; Zawadzki, J.V. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 1980, 288, 373–376. [Google Scholar] [CrossRef]
- Sunano, S.; Osugi, S.; Shimamura, K. Blood pressure and impairment of endothelium-dependent relaxation in spontaneously hypertensive rats. Experientia 1989, 45, 705–708. [Google Scholar] [CrossRef] [PubMed]
- Van der Loo, B.; Labugger, R.; Skepper, J.N.; Bachschmid, M.; Kilo, J.; Powell, J.M.; Palacios-Callender, M.; Erusalimsky, J.D.; Quaschning, T.; Malinski, T.; et al. Enhanced peroxynitrite formation is associated with vascular aging. J. Exp. Med. 2000, 192, 1731–1744. [Google Scholar] [CrossRef] [PubMed]
- Bredt, D.S.; Hwang, P.M.; Glatt, C.E.; Lowenstein, C.; Reed, R.R.; Snyder, S.H. Cloned and expressed nitric oxide synthase structurally resembles cytochromeP-450 reductase. Nature 1991, 351, 714–718. [Google Scholar] [CrossRef]
- Hoshida, S.; Yamashita, N.; Igarashi, J.; Nishida, M.; Hori, M.; Kamada, T.; Kuzuya, T.; Tada, M. Nitric oxidase synthase protects the heart against ischemia-reperfusion injury in rabbits. J. Pharmacol. Exp. Ther. 1995, 274, 413–418. [Google Scholar]
- Kuppusamy, P.; Ohnishi, S.T.; Numagami, Y.; Ohnishi, T.; Zweier, J.L. Three-dimensional imaging of nitric oxide production in the rat brain subjected to ischemia-hypoxia. J. Cereb. Blood Flow Metab. 1995, 15, 899–903. [Google Scholar] [CrossRef] [Green Version]
- Asano, Y.; Araki, N.; Ohkubo, T.; Kim, K.; Sawada, M.; Furuya, D.; Takei, K.; Shimazu, K. The study of brain nitric oxide production and cerebral blood flow in a rat model of forebrain ischemia and reperfusion. Jpn Cereb. Blood Flow Metab. 2000, 12, 130–131. [Google Scholar]
- Strolin Benedetti, M.; Dostert, P.; Marrari, P.; Cini, M. Effect of ageing on tissue levels of amino acids involved in the nitric oxide pathway in rat brain. J. Neural. Transm. 1993, 94, 21–30. [Google Scholar] [CrossRef]
- Siles, E.; Martínez-Lara, E.; Cañuelo, A.; Sánchez, M.; Hernández, R.; López-Ramos, J.C.; Del Moral, M.L.; Esteban, F.J.; Blanco, S.; Pedrosa, J.A.; et al. Age-related changes of the nitric oxide system in the rat brain. Brain Res. 2002, 956, 385–392. [Google Scholar] [CrossRef]
- Bredt, D.S.; Snyder, S.H. Isolation of nitric oxide synthetase, a calmodulin-requiring enzyme. Proc. Natl. Acad. Sci. USA 1990, 87, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Doi, T.; Ando, M.; Akaike, T.; Suga, M.; Sato, K.; Maeda, H. Resistance to nitric oxide in Mycobacterium avium complex and its implication in pathogenesis. Infect. Immun. 1993, 61, 1980–1989. [Google Scholar] [CrossRef]
- Yamada, K.; Nabeshima, T. Simultaneous measurement of nitrite and nitrate levels as indices of nitric oxide release in the cerebellum of conscious rats. J. Neurochem. 1997, 68, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Reichner, J.S.; Mateo, R.B.; Albina, J.E. Activated murine macrophages induce apoptosis in tumor cells through nitric oxide-dependent or independent mechanisms. Cancer Res. 1994, 54, 2462–2467. [Google Scholar] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ito, Y.; Nagoya, H.; Yamazato, M.; Asano, Y.; Sawada, M.; Shimazu, T.; Hirayama, M.; Yamamoto, T.; Araki, N. The Effect of Aging on Nitric Oxide Production during Cerebral Ischemia and Reperfusion in Wistar Rats and Spontaneous Hypertensive Rats: An In Vivo Microdialysis Study. Int. J. Mol. Sci. 2023, 24, 12749. https://doi.org/10.3390/ijms241612749
Ito Y, Nagoya H, Yamazato M, Asano Y, Sawada M, Shimazu T, Hirayama M, Yamamoto T, Araki N. The Effect of Aging on Nitric Oxide Production during Cerebral Ischemia and Reperfusion in Wistar Rats and Spontaneous Hypertensive Rats: An In Vivo Microdialysis Study. International Journal of Molecular Sciences. 2023; 24(16):12749. https://doi.org/10.3390/ijms241612749
Chicago/Turabian StyleIto, Yasuo, Harumitsu Nagoya, Masamizu Yamazato, Yoshio Asano, Masahiko Sawada, Tomokazu Shimazu, Makiko Hirayama, Toshimasa Yamamoto, and Nobuo Araki. 2023. "The Effect of Aging on Nitric Oxide Production during Cerebral Ischemia and Reperfusion in Wistar Rats and Spontaneous Hypertensive Rats: An In Vivo Microdialysis Study" International Journal of Molecular Sciences 24, no. 16: 12749. https://doi.org/10.3390/ijms241612749