
Regulation of Notch1 Signalling by Long Non-Coding RNAs in Cancers and Other Health Disorders
 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
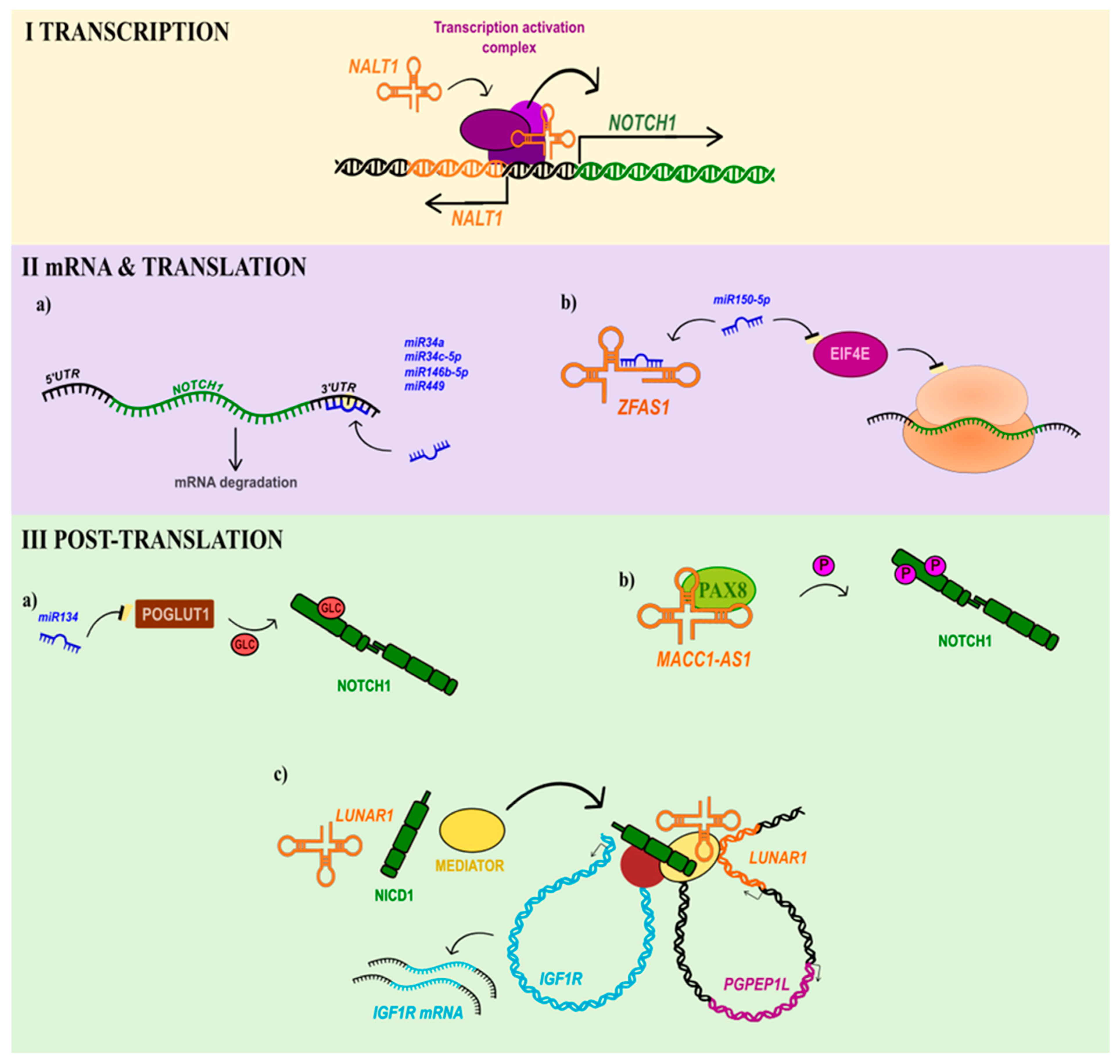
2. Non-Coding RNAs as a Pivotal Control Factor of Notch Signalling
2.1. Controlling Transcription of NOTCH1 and NOTCH1-Related Genes in Cancers
2.2. NOTCH1 mRNA and Translation Control
2.3. Controlling Notch1 Signalling by lncRNA at the Post-Translational Level in Cancers
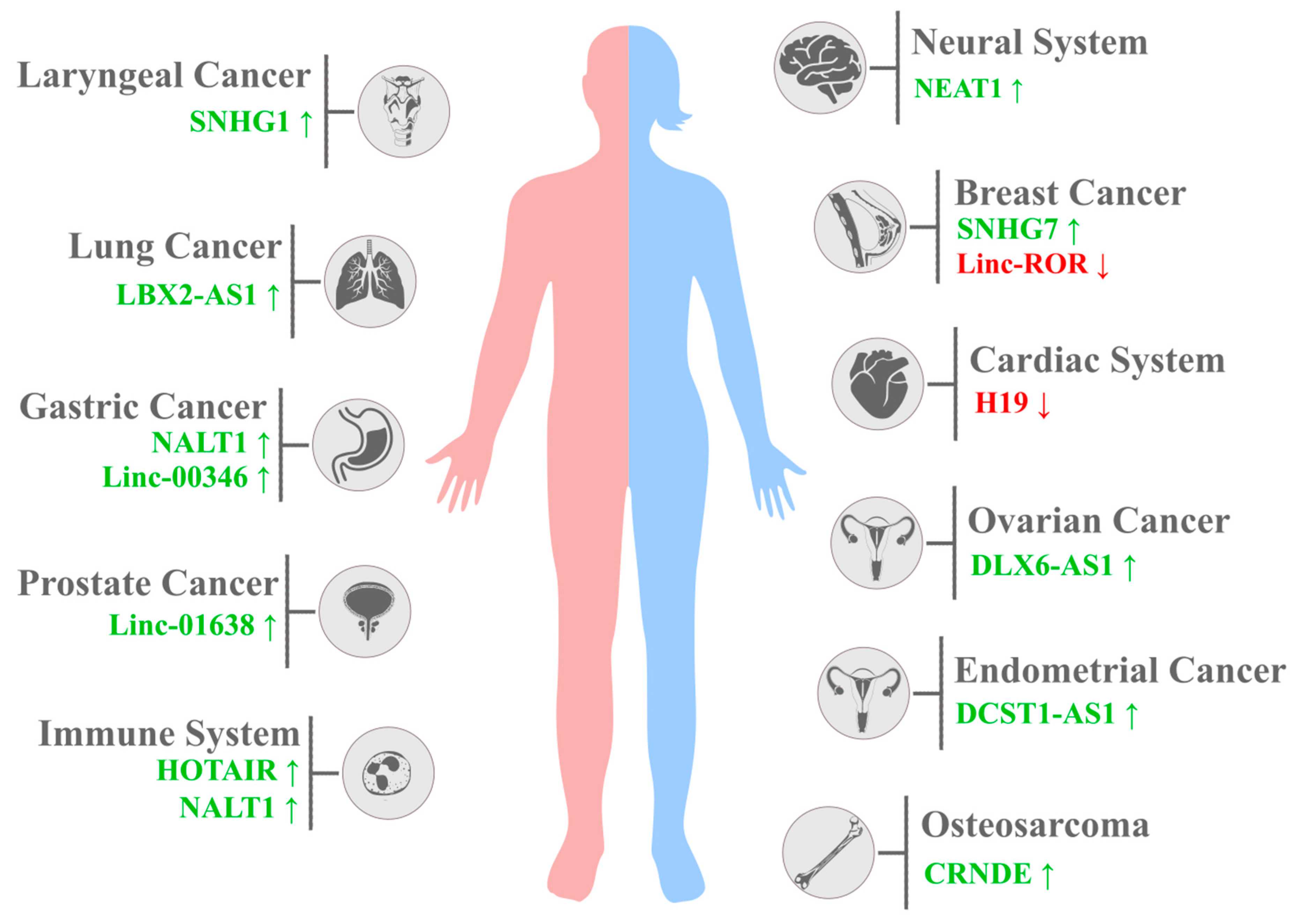
3. Notch1-Related lncRNAs and Other Diseases

{kind=link}
{kind=link}
| Localisation | Disease | lncRNA | NOTCH1 Expression | Cell Line | Animal Models | Patients | Reference |
|---|---|---|---|---|---|---|---|
| Cardiac system | Acute myocardial infarction | XIST↑ | ↑ | - | AMI rat model | - | [123] |
| Calcific aortic valve disease | H19↑ | ↓ | VICs, Saos2, COS7 | - | 36 patients | [124] | |
| Myocardial infarction | KCNQ1OT1↑ | ↑ | - | C57BL/6 male mice | - | [120] | |
| Ischemic stroke | H19↑ | ↓ | - | Male C57BL/6 J mice | 40 patients | [125] | |
| Immune system | T-ALL | NALT1↑ | ↑ | Jurkat cells | - | Bone marrow of 20 children | [78] |
| Systemic sclerosis (SSc) | HOTAIR↑ | ↑ | - | - | 12 adult patients | [118] | |
| Neural system | Epilepsy | NEAT1↑ | ↑ | CTX-TNA | - | 6 patients | [126] |
| Intervertebral disc degeneration | FAM83H-AS1↑ | ↑ | - | - | 10 patients | [127] | |
| Nasopharyngeal carcinoma | SNHG12↑ | ↑ | SUNE1, CNE1, CNE2 68, HNE-1 | - | 139 tissue samples | [128] | |
| Head and neck cancer | Oesophageal cancer | MALAT1↑ | ↑ | TE-1, EC109, KYSE30, OE21 | - | - | [129] |
| SNHG1↑ | ↑ | Eca109, TE-1 | - | 72 patients | [130] | ||
| Laryngeal cancer | SNHG1↑ | ↑ | - | - | 42 patients (different tumour stages) | [131] | |
| Digestive system cancer | Pancreatic carcinoma | MACC1-AS1↑ | ↑ | BxPC-3, PANC-1, MIA PaCa-2, KP-2, AsPC-1, Capan-1 | - | 2 cohorts (98 and 124 patients) of primary tissues | [105] |
| Gastric cancer | NALT1 (LINC01573)↑ | ↑ | SGC-7901, BGC-823 | - | 336 patients after D2 lymph node dissected gastrectomy | [79] | |
| LINC00346↑ (sponge for miR-34a-5p) | ↑ | MGC803, BGC823, MKN28, MKN45, SGC7901 | Xenografts in athymic (nu/nu) mice | 58 gastric adenocarcinoma tissue samples | [88] | ||
| Colorectal carcinoma | FAM83H-AS1↑ | ↑ | SW480, LoVo, HCT116, HT29 | - | 40 patients | [132] | |
| MALAT1↑ | ↑ | COLO205, HCT-116, LoVo, HT26, SW480 | Nude Balb/c mice | - | [133] | ||
| Hepatocellular carcinoma | LINC00261↓ | ↑ | SMCC-7721, MHCC97L, MHCC97H | - | 66 tissue samples | [134] | |
| Reproductive tract | Ovarian cancer | DLX6-AS1↑ | ↑ | HEY, SKOV3, OVCAR-3 | - | 128 tissue samples | [95] |
| MALAT1↑ | ↑ | A2780, OVCAR3, COC1, A2780/ CDDP, COC1/CDDP, OVCAR3/DDP | - | 20 paired tumour tissue samples | [135] | ||
| Endometrial carcinoma (EC) | MEG3↓ | ↑ | HEC-1A, KLE | - | 30 tumour tissue samples | [136] | |
| DCST1-AS1↑ | ↑ | HEC-1 | - | 62 patients | [90] | ||
| Prostate carcinoma | LINCO1638↑ | ↑ | - | - | 42 patients | [137] | |
| GHET1↑ | ↑ | LNCap, C4-2 | - | 30 patients | [138] | ||
| Other cancers | Breast cancer | linc-ROR↑ | ↓ | MDA-MB-231 | - | - | [40] |
| SNHG7↑ | ↑ | - | - | 37 pairs of tumour tissue samples | [84] | ||
| Lung cancer | NBR2↓ | ↑ | - | - | 50 NSCLC patient tissue samples | [101] | |
| LBX2-AS1↑ | ↑ | A549, PC9, H1975, SPC-A1, H1299 | - | 165 NSCLC patients | [139] | ||
| EGFR-AS1↓ | ↑ | NCH-H460, NCH-H23 | - | 87 NSCLC patients | [140] | ||
| LET↓ | ↑ | A549, 95D, NCI-H292, NCI-H1975 | - | 66 NSCLC patients | [141] | ||
| Osteosarcoma | MEG3↓ | ↑ | MG-63, U2OS | - | - | [142] | |
| NBR2↓ | ↑ | MG-63, U2OS, SAOS-2 | - | 62 patients | [143] | ||
| CRNDE↑ | ↑ | MG-63, SAOS-2, U2OS | - | 72 patients | [144] |
4. Medicinal Perspectives for lncRNAs in Notch1-Related Diseases
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bray, S.J. Notch signalling: A simple pathway becomes complex. Nat. Rev. Mol. Cell Biol. 2006, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Cordle, J.; Johnson, S.; Tay, J.Z.Y.; Roversi, P.; Wilkin, M.B.; de Madrid, B.H.; Shimizu, H.; Jensen, S.; Whiteman, P.; Jin, B.; et al. A conserved face of the Jagged/Serrate DSL domain is involved in Notch trans-activation and cis-inhibition. Nat. Struct. Mol. Biol. 2008, 15, 849–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopan, R.; Ilagan, M.X.G. The canonical Notch signaling pathway: Unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henrique, D.; Schweisguth, F. Mechanisms of Notch signaling: A simple logic deployed in time and space. Development 2019, 146, dev172148. [Google Scholar] [CrossRef] [Green Version]
- Nandagopal, N.; Santat, L.A.; LeBon, L.; Sprinzak, D.; Bronner, M.E.; Elowitz, M.B. Dynamic Ligand Discrimination in the Notch Signaling Pathway. Cell 2018, 172, 869–880.e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaya, O.; Binshtok, U.; Hersch, M.; Rivkin, D.; Weinreb, S.; Amir-Zilberstein, L.; Khamaisi, B.; Oppenheim, O.; Desai, R.A.; Goodyear, R.J.; et al. Cell-Cell Contact Area Affects Notch Signaling and Notch-Dependent Patterning. Dev. Cell 2017, 40, 505–511.e6. [Google Scholar] [CrossRef] [Green Version]
- Dexter, J.S. The Analysis of a Case of Continuous Variation in Drosophila by a Study of Its Linkage Relations. In The American Naturalist; The University of Chicago Press: Chicago, IL, USA, 1914; Volume 48, pp. 712–758. [Google Scholar]
- Morgan, T.H.; Bridges, C.B. The Inheritance of a Fluctuating Character. J. Gen. Physiol. 1919, 1, 639–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, S.; Schulze, K.L.; Bellen, H.J. Introduction to Notch Signaling. In Methods in Molecular Biology (Clifton, N.J.); Humana Press: Tutowa, NJ, USA, 2014; Volume 1187, pp. 1–14. [Google Scholar]
- Bigas, A.; Espinosa, L. Notch Signaling in Cell–Cell Communication Pathways. Curr. Stem Cell Rep. 2016, 2, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Brai, E.; Marathe, S.; Astori, S.; Ben Fredj, N.; Perry, E.; Lamy, C.; Scotti, A.; Alberi, L. Notch1 Regulates Hippocampal Plasticity Through Interaction with the Reelin Pathway, Glutamatergic Transmission and CREB Signaling. Front. Cell. Neurosci. 2015, 9, 447. [Google Scholar] [CrossRef] [Green Version]
- Kidd, S.; Lieber, T. Mechanism of Notch Pathway Activation and Its Role in the Regulation of Olfactory Plasticity in Drosophila melanogaster. PLoS ONE 2016, 11, e0151279. [Google Scholar] [CrossRef]
- Pitulescu, M.E.; Schmidt, I.; Giaimo, B.D.; Antoine, T.; Berkenfeld, F.; Ferrante, F.; Park, H.; Ehling, M.; Biljes, D.; Rocha, S.F.; et al. Dll4 and Notch signalling couples sprouting angiogenesis and artery formation. Nat. Cell Biol. 2017, 19, 915–927. [Google Scholar] [CrossRef] [Green Version]
- Hasan, S.S.; Tsaryk, R.; Lange, M.; Wisniewski, L.; Moore, J.C.; Lawson, N.D.; Wojciechowska, K.; Schnittler, H.; Siekmann, A.F. Endothelial Notch signalling limits angiogenesis via control of artery formation. Nat. Cell Biol. 2017, 19, 928–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aspalter, I.M.; Gordon, E.; Dubrac, A.; Ragab, A.; Narloch, J.; Vizán, P.; Geudens, I.; Collins, R.T.; Franco, C.A.; Abrahams, C.L.; et al. Alk1 and Alk5 inhibition by Nrp1 controls vascular sprouting downstream of Notch. Nat. Commun. 2015, 6, 7264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobov, I.; Mikhailova, N. The Role of Dll4/Notch Signaling in Normal and Pathological Ocular Angiogenesis: Dll4 Controls Blood Vessel Sprouting and Vessel Remodeling in Normal and Pathological Conditions. J. Ophthalmol. 2018, 2018, 3565292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antfolk, D.; Sjöqvist, M.; Cheng, F.; Isoniemi, K.; Duran, C.L.; Rivero-Muller, A.; Antila, C.; Niemi, R.; Landor, S.; Bouten, C.V.C.; et al. Selective regulation of Notch ligands during angiogenesis is mediated by vimentin. Proc. Natl. Acad. Sci. USA 2017, 114, E4574–E4581. [Google Scholar] [CrossRef] [PubMed]
- Travisano, S.I.; Oliveira, V.L.; Prados, B.; Grego-Bessa, J.; Piñeiro-Sabarís, R.; Bou, V.; Gómez, M.J.; Sánchez-Cabo, F.; MacGrogan, D.; de la Pompa, J.L. Coronary arterial development is regulated by a Dll4-Jag1-EphrinB2 signaling cascade. Elife 2019, 8, e49977. [Google Scholar] [CrossRef]
- Urbanek, K.; Lesiak, M.; Krakowian, D.; Koryciak-Komarska, H.; Likus, W.; Czekaj, P.; Kusz, D.; Sieroń, A.L. Notch signaling pathway and gene expression profiles during early in vitro differentiation of liver-derived mesenchymal stromal cells to osteoblasts. Lab. Investig. 2017, 97, 1225–1234. [Google Scholar] [CrossRef] [Green Version]
- Ziouti, F.; Ebert, R.; Rummler, M.; Krug, M.; Müller-Deubert, S.; Lüdemann, M.; Jakob, F.; Willie, B.M.; Jundt, F. NOTCH Signaling Is Activated through Mechanical Strain in Human Bone Marrow-Derived Mesenchymal Stromal Cells. Stem Cells Int. 2019, 2019, 5150634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sjöqvist, M.; Andersson, E.R. Do as I say, Not(ch) as I do: Lateral control of cell fate. Dev. Biol. 2019, 447, 58–70. [Google Scholar] [CrossRef]
- Koon, Y.L.; Zhang, S.; Rahmat, M.B.; Koh, C.G.; Chiam, K.-H. Enhanced Delta-Notch Lateral Inhibition Model Incorporating Intracellular Notch Heterogeneity and Tension-Dependent Rate of Delta-Notch Binding that Reproduces Sprouting Angiogenesis Patterns. Sci. Rep. 2018, 8, 9519. [Google Scholar] [CrossRef] [Green Version]
- Menchero, S.; Rollan, I.; Lopez-Izquierdo, A.; Andreu, M.J.; Sainz de Aja, J.; Kang, M.; Adan, J.; Benedito, R.; Rayon, T.; Hadjantonakis, A.-K.; et al. Transitions in cell potency during early mouse development are driven by Notch. Elife 2019, 8, e42930. [Google Scholar] [CrossRef]
- Garcıá-Leoń, M.J.; Fuentes, P.; de la Pompa, J.L.; Toribio, M.L. Dynamic regulation of NOTCH1 activation and notch ligand expression in human thymus development. Development 2018, 145, dev165597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mack, J.J.; Mosqueiro, T.S.; Archer, B.J.; Jones, W.M.; Sunshine, H.; Faas, G.C.; Briot, A.; Aragón, R.L.; Su, T.; Romay, M.C.; et al. NOTCH1 is a mechanosensor in adult arteries. Nat. Commun. 2017, 8, 1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Shang, X.; Zhang, H.; Wang, G.; Massey, P.A.; Barton, S.R.; Kevil, C.G.; Dong, Y. Notch Signaling in Osteogenesis, Osteoclastogenesis, and Angiogenesis. Am. J. Pathol. 2019, 189, 1495–1500. [Google Scholar] [CrossRef] [Green Version]
- Grazioli, P.; Felli, M.P.; Screpanti, I.; Campese, A.F. The mazy case of Notch and immunoregulatory cells. J. Leukoc. Biol. 2017, 102, 361–368. [Google Scholar] [CrossRef] [Green Version]
- Arcaini, L.; Rossi, D.; Lucioni, M.; Nicola, M.; Bruscaggin, A.; Fiaccadori, V.; Riboni, R.; Ramponi, A.; Ferretti, V.V.; Cresta, S.; et al. The NOTCH pathway is recurrently mutated in diffuse large B-cell lymphoma associated with hepatitis C virus infection. Haematologica 2015, 100, 246–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Masiero, M.; Banham, A.H. Notch signaling: Its roles and therapeutic potential in hematological malignancies. Oncotarget 2016, 7, 29804–29823. [Google Scholar] [CrossRef] [Green Version]
- Takam Kamga, P.; Collo, G.D.; Resci, F.; Bazzoni, R.; Mercuri, A.; Quaglia, F.M.; Tanasi, I.; Delfino, P.; Visco, C.; Bonifacio, M.; et al. Notch Signaling Molecules as Prognostic Biomarkers for Acute Myeloid Leukemia. Cancers 2019, 11, 1958. [Google Scholar] [CrossRef] [Green Version]
- Teodorczyk, M.; Schmidt, M.H.H. Notching on Cancer’s Door: Notch Signaling in Brain Tumors. Front. Oncol. 2015, 4, 341. [Google Scholar] [CrossRef] [Green Version]
- Krishna, B.M.; Jana, S.; Singhal, J.; Horne, D.; Awasthi, S.; Salgia, R.; Singhal, S.S. Notch signaling in breast cancer: From pathway analysis to therapy. Cancer Lett. 2019, 461, 123–131. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Y.; Liu, H.; Kang, X.; Li, W.; Wei, Q. Genetic variants of genes in the Notch signaling pathway predict overall survival of non-small cell lung cancer patients in the PLCO study. Oncotarget 2016, 7, 61716–61727. [Google Scholar] [CrossRef] [PubMed]
- Zou, B.; Zhou, X.; Lai, S.; Liu, J. Notch signaling and non-small cell lung cancer (Review). Oncol. Lett. 2018, 15, 3415–3421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Liu, N. A 10-gene expression signature of Notch pathway predicts recurrence in ovarian carcinoma. Oncol. Lett. 2015, 10, 1704–1708. [Google Scholar] [CrossRef] [Green Version]
- Bhagat, T.D.; Zou, Y.; Huang, S.; Park, J.; Palmer, M.B.; Hu, C.; Li, W.; Shenoy, N.; Giricz, O.; Choudhary, G.; et al. Notch Pathway Is Activated via Genetic and Epigenetic Alterations and Is a Therapeutic Target in Clear Cell Renal Cancer. J. Biol. Chem. 2017, 292, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Li, J.; Zheng, J.; Wei, A. The Carcinogenic Role of the Notch Signaling Pathway in the Development of Hepatocellular Carcinoma. J. Cancer 2019, 10, 1570–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, D.; Feng, F.; An, L.; Hui, F.; Dang, D.; Zhao, Q. Progressive and Prognosis Value of Notch Receptors and Ligands in Hepatocellular Carcinoma: A Systematic Review and Meta-analysis. Sci. Rep. 2017, 7, 14809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kałafut, J.; Czapiński, J.; Przybyszewska-Podstawka, A.; Czerwonka, A.; Odrzywolski, A.; Sahlgren, C.; Rivero-Müller, A. Optogenetic control of NOTCH1 signaling. Cell Commun. Signal. 2022, 20, 67. [Google Scholar] [CrossRef]
- Chen, Y.M.; Liu, Y.; Wei, H.Y.; Lv, K.Z.; Fu, P.F. Large intergenic non-coding RNA-ROR reverses Gemcitabineinduced autophagy and apoptosis in breast cancer cells. Oncotarget 2016, 7, 59604–59617. [Google Scholar] [CrossRef]
- Li, D.; Xu, D.; Zhang, Y.; Chen, P.; Xie, J. Effect of Notch1 signaling on cellular proliferation and apoptosis in human laryngeal carcinoma. World J. Surg. Oncol. 2022, 20, 262. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-L.; Chen, C.; Yang, Y.; Wang, C.; Yang, T.; Yang, X.; Liu, S.-C. Gamma secretase inhibitor enhances sensitivity to doxorubicin in MDA-MB-231 cells. Int. J. Clin. Exp. Pathol. 2015, 8, 4378–4387. [Google Scholar]
- Zang, S.; Chen, F.; Dai, J.; Guo, D.; Tse, W.; Qu, X.; Ma, D.; Ji, C. RNAi-mediated knockdown of Notch-1 leads to cell growth inhibition and enhanced chemosensitivity in human breast cancer. Oncol. Rep. 2010, 23, 893–899. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.; Song, B.; Shi, Y.; Wang, B.; Fan, S.; Yu, X.; Tang, J.; Li, L. ShRNA targeting Notch1 sensitizes breast cancer stem cell to paclitaxel. Int. J. Biochem. Cell Biol. 2013, 45, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Do, S.I.; Lee, H.J.; Kang, H.J.; Koo, B.S.; Lim, Y.C. Notch1 signaling contributes to stemness in head and neck squamous cell carcinoma. Lab. Investig. 2016, 96, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Katarkar, A.; Bottoni, G.; Clocchiatti, A.; Goruppi, S.; Bordignon, P.; Lazzaroni, F.; Gregnanin, I.; Ostano, P.; Neel, V.; Dotto, G.P. NOTCH1 gene amplification promotes expansion of Cancer Associated Fibroblast populations in human skin. Nat. Commun. 2020, 11, 5126. [Google Scholar] [CrossRef] [PubMed]
- Misiorek, J.O.; Przybyszewska-Podstawka, A.; Kałafut, J.; Paziewska, B.; Rolle, K.; Rivero-Müller, A.; Nees, M. Context Matters: NOTCH Signatures and Pathway in Cancer Progression and Metastasis. Cells 2021, 10, 94. [Google Scholar] [CrossRef]
- Aster, J.C.; Pear, W.S.; Blacklow, S.C. The Varied Roles of Notch in Cancer. Annu. Rev. Pathol. 2017, 12, 245–275. [Google Scholar] [CrossRef] [Green Version]
- Meisel, C.T.; Porcheri, C.; Mitsiadis, T.A. Cancer Stem Cells, Quo Vadis? The Notch Signaling Pathway in Tumor Initiation and Progression. Cells 2020, 9, 1879. [Google Scholar] [CrossRef]
- Lee, B.; Lee, S.; Shim, J. YTHDF2 Suppresses Notch Signaling through Post-transcriptional Regulation on Notch1. Int. J. Biol. Sci. 2021, 17, 3776–3785. [Google Scholar] [CrossRef] [PubMed]
- Salviano-Silva, A.; Berti, F.C.B.; Lobo-Alves, S.C.; de Araujo-Souza, P.S.; Boldt, A.B.W.; Malheiros, D. Interaction of Long Noncoding RNAs and Notch Signaling: Implications for Tissue Homeostasis Loss. In Notch Signaling in Embryology and Cancer; Springer: Cham, Switzerland, 2020; pp. 107–129. ISBN 978-3-030-36421-2. [Google Scholar]
- Shen, Y.; Li, C.; Zhou, L.; Huang, J. G protein-coupled oestrogen receptor promotes cell growth of non-small cell lung cancer cells via YAP1/QKI/circNOTCH1/m6A methylated NOTCH1 signalling. J. Cell. Mol. Med. 2020, 25, 284–296. [Google Scholar] [CrossRef]
- Wu, X.; Ye, W.; Gong, Y. The Role of RNA Methyltransferase METTL3 in Normal and Malignant Hematopoiesis. Front. Oncol. 2022, 12, 873903. [Google Scholar] [CrossRef]
- Antfolk, D.; Antila, C.; Kemppainen, K.; Landor, S.K.J.; Sahlgren, C. Decoding the PTM-switchboard of Notch. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118507. [Google Scholar] [CrossRef]
- Li, N.; Fassl, A.; Chick, J.; Inuzuka, H.; Li, X.; Mansour, M.R.; Liu, L.; Wang, H.; King, B.; Shaik, S.; et al. Cyclin C is a haploinsufficient tumour suppressor. Nat. Cell Biol. 2014, 16, 1080–1091. [Google Scholar] [CrossRef] [Green Version]
- Ranganathan, P.; Vasquez-Del Carpio, R.; Kaplan, F.M.; Wang, H.; Gupta, A.; VanWye, J.D.; Capobianco, A.J. Hierarchical phosphorylation within the ankyrin repeat domain defines a phosphoregulatory loop that regulates notch transcriptional activity. J. Biol. Chem. 2011, 286, 28844–28857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santio, N.M.; Landor, S.K.J.; Vahtera, L.; Ylä-Pelto, J.; Paloniemi, E.; Imanishi, S.Y.; Corthals, G.; Varjosalo, M.; Manoharan, G.B.; Uri, A.; et al. Phosphorylation of Notch1 by Pim kinases promotes oncogenic signaling in breast and prostate cancer cells. Oncotarget 2016, 7, 43220–43238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Park, S.; Kim, M.; Shin, I. Down-regulation of Notch-dependent transcription by Akt in vitro. FEBS Lett. 2008, 582, 1693–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersson, S.; Sczaniecka, M.; McLaren, L.; Russell, F.; Gladstone, K.; Hupp, T.; Wallace, M. Non-degradative ubiquitination of the Notch1 receptor by the E3 ligase MDM2 activates the Notch signalling pathway. Biochem. J. 2013, 450, 523–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and their Integrated Networks. J. Integr. Bioinform. 2019, 16, 20190027. [Google Scholar] [CrossRef]
- Du, Y.; Chen, H.; Li, X.-F.; Lu, Y.-C.; Feng, Y.-B.; Zhang, L. Molecular mechanism of Notch signaling with special emphasis on microRNAs: Implications for glioma. J. Cell. Physiol. 2019, 234, 158–170. [Google Scholar] [CrossRef] [Green Version]
- Majidinia, M.; Darband, S.G.; Kaviani, M.; Nabavi, S.M.; Jahanban-Esfahlan, R.; Yousefi, B. Cross-regulation between Notch signaling pathway and miRNA machinery in cancer. DNA Repair 2018, 66–67, 30–41. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Glassy, M.C.; Abak, A.; Hussen, B.M.; Niazi, V.; Taheri, M. The interaction between miRNAs/lncRNAs and Notch pathway in human disorders. Biomed. Pharmacother. 2021, 138, 111496. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Pan, C.-Y.; Lin, W. Overlapping protein-coding genes in human genome and their coincidental expression in tissues. Sci. Rep. 2019, 9, 13377. [Google Scholar] [CrossRef] [Green Version]
- Laurent, G.S.; Wahlestedt, C.; Kapranov, P. The Landscape of long non-coding RNA classification Georges. Trends Genet. 2016, 31, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Fernandes, J.C.R.; Acuña, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long non-coding RNAs in the regulation of gene expression: Physiology and disease. Non Coding RNA 2019, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karakas, D.; Ozpolat, B. The Role of LncRNAs in Translation. Non Coding RNA 2021, 7, 16. [Google Scholar] [CrossRef]
- Pisignano, G.; Pavlaki, I.; Murrell, A. Being in a loop: How long non-coding RNAs organise genome architecture. Essays Biochem. 2019, 63, 177–186. [Google Scholar] [CrossRef]
- Mishra, K.; Kanduri, C. Understanding Long Noncoding RNA and Chromatin Interactions: What We Know So Far. Non Coding RNA 2019, 5, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, W.-X.; Koirala, P.; Mo, Y.-Y. LncRNA-mediated regulation of cell signaling in cancer. Oncogene 2017, 36, 5661–5667. [Google Scholar] [CrossRef]
- Khorkova, O.; Hsiao, J.; Wahlestedt, C. Basic biology and therapeutic implications of lncRNA. Adv. Drug Deliv. Rev. 2015, 87, 15–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchese, F.P.; Raimondi, I.; Huarte, M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017, 18, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazave, E.; Lapébie, P.; Richards, G.S.; Brunet, F.; Ereskovsky, A.V.; Degnan, B.M.; Borchiellini, C.; Vervoort, M.; Renard, E. Origin and evolution of the Notch signalling pathway: An overview from eukaryotic genomes. BMC Evol. Biol. 2009, 9, 249. [Google Scholar] [CrossRef] [Green Version]
- Salazar, J.L.; Yamamoto, S. Integration of Drosophila and Human Genetics to Understand Notch Signaling Related Diseases; Springer: Cham, Switzerland, 2018; pp. 141–185. [Google Scholar]
- Meurette, O.; Mehlen, P. Notch Signaling in the Tumor Microenvironment. Cancer Cell 2018, 34, 536–548. [Google Scholar] [CrossRef] [Green Version]
- Lambertini, C.; Pantano, S.; Dotto, G.P. Differential Control of Notch1 Gene Transcription by Klf4 and Sp3 Transcription Factors in Normal versus Cancer-Derived Keratinocytes. PLoS ONE 2010, 5, e10369. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, P.; Lin, R.; Rong, L.; Xue, Y.; Fang, Y. LncRNA NALT interaction with NOTCH1 promoted cell proliferation in pediatric T cell acute lymphoblastic leukemia. Sci. Rep. 2015, 5, 13749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piao, H.Y.; Guo, S.; Wang, Y.; Zhang, J. Long noncoding RNA NALT1-induced gastric cancer invasion and metastasis via NOTCH signaling pathway. World J. Gastroenterol. 2019, 25, 6508–6526. [Google Scholar] [CrossRef]
- Kwon, H.; Song, K.; Han, C.; Zhang, J.; Lu, L.; Chen, W.; Wu, T. Epigenetic Silencing of miRNA-34a in Human Cholangiocarcinoma via EZH2 and DNA Methylation: Impact on Regulation of Notch Pathway. Am. J. Pathol. 2017, 187, 2288–2299. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Tzatzalos, E.; Kwan, K.Y.; Grumet, M.; Cai, L. Transcriptional Regulation of Notch1 Expression by Nkx6.1 in Neural Stem/Progenitor Cells during Ventral Spinal Cord Development. Sci. Rep. 2016, 6, 38665. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.F.; Liu, Y.H.; Wang, D.W.; Liu, T.S.; Yang, Y.; Guo, J.M.; Pan, Y.; Zhang, Y.F.; Du, H.; Li, L.; et al. Obesity-induced reduced expression of the lncRNA ROIT impairs insulin transcription by downregulation of Nkx6.1 methylation. Diabetologia 2020, 63, 811–824. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Li, C.; Huang, J.; Luo, P.; Mo, D.; Wang, H. LncRNA FEZF1-AS1 promotes non-small lung cancer cell migration and invasion through the up-regulation of NOTCH1 by serving as a sponge of miR-34a. BMC Pulm. Med. 2020, 20, 110. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Huang, T.; Liu, Z.; Sun, M.; Luo, S. LncRNA SNHG7 contributes to tumorigenesis and progression in breast cancer by interacting with miR-34a through EMT initiation and the Notch-1 pathway. Eur. J. Pharmacol. 2019, 856, 172407. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.Y.; Wang, Z.H.; Yu, Q.; Yuan, L.L.; Peng, H.L.; Xu, Y.X. LncRNA-NEAT1 promotes proliferation of T-ALL cells via miR-146b-5p/NOTCH1 signaling pathway. Pathol. Res. Pract. 2020, 216, 153212. [Google Scholar] [CrossRef]
- Rani, N.; Nowakowski, T.J.; Zhou, H.; Godshalk, S.E.; Lisi, V.; Kriegstein, A.R.; Kosik, K.S. A Primate lncRNA Mediates Notch Signaling during Neuronal Development by Sequestering miRNA. Neuron 2016, 90, 1174–1188. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wu, J.; Wu, C.; Chen, W.; Lin, R.; Zhou, Y.; Huang, X. The LINC01138 interacts with PRMT5 to promote SREBP1-mediated lipid desaturation and cell growth in clear cell renal cell carcinoma. Biochem. Biophys. Res. Commun. 2018, 507, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.P.; Ma, P.; Wang, W.Y.; Shuai, Y.; Wang, Y.F.; Yu, T.; Xia, R.; Shu, Y.Q. KLF5 and MYC modulated LINC00346 contributes to gastric cancer progression through acting as a competing endogeous RNA and indicates poor outcome. Cell Death Differ. 2019, 26, 2179–2193. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Huang, B.; Zhang, Q.; He, X.; Wei, H.; Zhang, D. NOTCH1 regulates the proliferation and migration of bladder cancer cells by cooperating with long non-coding RNA HCG18 and microRNA-34c-5p. J. Cell. Biochem. 2019, 120, 6596–6604. [Google Scholar] [CrossRef]
- Ke, J.; Shen, Z.; Hu, W.; Li, M.; Shi, Y.; Xie, Z.; Wu, D. LncRNA DCST1-AS1 was upregulated in endometrial carcinoma and may sponge miR-92a-3p to upregulate notch1. Cancer Manag. Res. 2020, 12, 1221–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ounzain, S.; Micheletti, R.; Arnan, C.; Plaisance, I.; Cecchi, D.; Schroen, B.; Reverter, F.; Alexanian, M.; Gonzales, C.; Ng, S.Y.; et al. CARMEN, a human super enhancer-associated long noncoding RNA controlling cardiac specification, differentiation and homeostasis. J. Mol. Cell. Cardiol. 2015, 89, 98–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaisance, I.; Perruchoud, S.; Fernandez-Tenorio, M.; Gonzales, C.; Ounzain, S.; Ruchat, P.; Nemir, M.; Niggli, E.; Pedrazzini, T. Cardiomyocyte Lineage Specification in Adult Human Cardiac Precursor Cells Via Modulation of Enhancer-Associated Long Noncoding RNA Expression. JACC Basic Transl. Sci. 2016, 1, 472–493. [Google Scholar] [CrossRef] [Green Version]
- Kolenda, T.; Guglas, K.; Kopczyńska, M.; Teresiak, A.; Bliźniak, R.; Mackiewicz, A.; Lamperska, K.; Mackiewicz, J. Oncogenic Role of ZFAS1 lncRNA in Head and Neck Squamous Cell Carcinomas. Cells 2019, 8, 366. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Lin, Z.; Zuo, C.; Fu, Y. Upregulation of miR-150-5p alleviates LPS-induced inflammatory response and apoptosis of RAW264.7 macrophages by targeting Notch1. Open Life Sci. 2020, 15, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Liu, H.R. Down-regulation of long noncoding RNA DLX6-AS1 defines good prognosis and inhibits proliferation and metastasis in human epithelial ovarian cancer cells via Notch signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 3243–3252. [Google Scholar] [CrossRef]
- An, Y.; Chen, X.M.; Yang, Y.; Mo, F.; Jiang, Y.; Sun, D.L.; Cai, H.H. LncRNA DLX6-AS1 promoted cancer cell proliferation and invasion by attenuating the endogenous function of miR-181b in pancreatic cancer 11 Medical and Health Sciences 1112 Oncology and Carcinogenesis. Cancer Cell Int. 2018, 18, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, R.; Huang, Y.; Wang, J.; Liu, Y. Knockdown of lncRNA DLX6-AS1 inhibits cell proliferation, migration and invasion while promotes apoptosis by downregulating PRR11 expression and upregulating miR-144 in non-small cell lung cancer. Biomed. Pharmacother. 2019, 109, 1851–1859. [Google Scholar] [CrossRef]
- Li, X.; Zhang, H.; Wu, X. Long noncoding RNA DLX6-AS1 accelerates the glioma carcinogenesis by competing endogenous sponging miR-197-5p to relieve E2F1. Gene 2019, 686, 1–7. [Google Scholar] [CrossRef]
- Petrovic, J.; Zhou, Y.; Fasolino, M.; Goldman, N.; Schwartz, G.W.; Mumbach, M.R.; Nguyen, S.C.; Rome, K.S.; Sela, Y.; Zapataro, Z.; et al. Oncogenic Notch Promotes Long-Range Regulatory Interactions within Hyperconnected 3D Cliques. Mol. Cell 2019, 73, 1174–1190.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Xiao, Z.D.; Han, L.; Zhang, J.; Lee, S.W.; Wang, W.; Lee, H.; Zhuang, L.; Chen, J.; Lin, H.K.; et al. LncRNA NBR2 engages a metabolic checkpoint by regulating AMPK under energy stress. Nat. Cell Biol. 2016, 18, 431–442. [Google Scholar] [CrossRef]
- Gao, Y.P.; Li, Y.; Li, H.J.; Zhao, B. LncRNA NBR2 inhibits EMT progression by regulating Notch 1 pathway in NSCLC. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7950–7958. [Google Scholar] [CrossRef] [PubMed]
- Ellisen, L.W.; Bird, J.; West, D.C.; Soreng, A.L.; Reynolds, T.C.; Smith, S.D.; Sklar, J. TAN-1, the human homolog of the Drosophila notch gene, is broken by chromosomal translocations in T lymphoblastic neoplasms. Cell 1991, 66, 649–661. [Google Scholar] [CrossRef]
- Trimarchi, T.; Bilal, E.; Ntziachristos, P.; Fabbri, G.; Dalla-Favera, R.; Tsirigos, A.; Aifantis, I. Genome-wide mapping and characterization of notch-regulated long noncoding RNAs in acute leukemia. Cell 2014, 158, 593–606. [Google Scholar] [CrossRef] [Green Version]
- Qian, J.; Garg, A.; Li, F.; Shen, Q.; Xiao, K. lncRNA LUNAR1 accelerates colorectal cancer progression by targeting the miR 495 3p/MYCBP axis. Int. J. Oncol. 2020, 57, 1157–1168. [Google Scholar] [CrossRef]
- Chen, Q.; Chen, X.F.; Li, D.E.; Yang, L.; Xu, L.P.; Hu, Y.; Jiang, J.S. Long non-coding RNA MACC1-AS1 promoted pancreatic carcinoma progression through activation of PAX8/NOTCH1 signaling pathway. J. Exp. Clin. Cancer Res. 2019, 38, 344. [Google Scholar] [CrossRef]
- Cai, Z.; Zhao, B.; Deng, Y.; Shangguan, S.; Zhou, F.; Zhou, W.; Li, X.; Li, Y.; Chen, G. Notch signaling in cerebrovascular diseases (Review). Mol. Med. Rep. 2016, 14, 2883–2898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malashicheva, A.; Kostina, A.; Kostareva, A.; Irtyuga, O.; Uspensky, V. Notch signaling in the pathogenesis of thoracic aortic aneurysms: A bridge between embryonic and adult states. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1866, 165631. [Google Scholar] [CrossRef] [PubMed]
- MacGrogan, D.; Münch, J.; de la Pompa, J.L. Notch and interacting signalling pathways in cardiac development, disease, and regeneration. Nat. Rev. Cardiol. 2018, 15, 685–704. [Google Scholar] [CrossRef]
- Ho, D.M.; Artavanis-Tsakonas, S.; Louvi, A. The Notch pathway in CNS homeostasis and neurodegeneration. WIREs Dev. Biol. 2020, 9, e358. [Google Scholar] [CrossRef]
- Smith, K.A.; Voiriot, G.; Tang, H.; Fraidenburg, D.R.; Song, S.; Yamamura, H.; Yamamura, A.; Guo, Q.; Wan, J.; Pohl, N.M.; et al. Notch Activation of Ca2+ Signaling in the Development of Hypoxic Pulmonary Vasoconstriction and Pulmonary Hypertension. Am. J. Respir. Cell Mol. Biol. 2015, 53, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Turnpenny, P.D.; Alman, B.; Cornier, A.S.; Giampietro, P.F.; Offiah, A.; Tassy, O.; Pourquié, O.; Kusumi, K.; Dunwoodie, S. Abnormal vertebral segmentation and the notch signaling pathway in man. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2007, 236, 1456–1474. [Google Scholar] [CrossRef] [PubMed]
- Servián-Morilla, E.; Takeuchi, H.; Lee, T.V.; Clarimon, J.; Mavillard, F.; Area-Gómez, E.; Rivas, E.; Nieto-González, J.L.; Rivero, M.C.; Cabrera-Serrano, M.; et al. A POGLUT 1 mutation causes a muscular dystrophy with reduced Notch signaling and satellite cell loss. EMBO Mol. Med. 2016, 8, 1289–1309. [Google Scholar] [CrossRef] [PubMed]
- Karaca, E.; Yuregir, O.O.; Bozdogan, S.T.; Aslan, H.; Pehlivan, D.; Jhangiani, S.N.; Akdemir, Z.C.; Gambin, T.; Bayram, Y.; Atik, M.M.; et al. Rare variants in the notch signaling pathway describe a novel type of autosomal recessive Klippel-Feil syndrome. Am. J. Med. Genet. Part A 2015, 167, 2795–2799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adami, G.; Rossini, M.; Gatti, D.; Orsolini, G.; Idolazzi, L.; Viapiana, O.; Scarpa, A.; Canalis, E. Hajdu Cheney Syndrome; report of a novel NOTCH2 mutation and treatment with denosumab. Bone 2016, 92, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Mašek, J.; Andersson, E.R. The developmental biology of genetic Notch disorders. Development 2017, 144, 1743–1763. [Google Scholar] [CrossRef] [Green Version]
- Arreola, A.; Payne, L.B.; Julian, M.H.; de Cubas, A.A.; Daniels, A.B.; Taylor, S.; Zhao, H.; Darden, J.; Bautch, V.L.; Rathmell, W.K.; et al. Von Hippel-Lindau mutations disrupt vascular patterning and maturation via Notch. JCI Insight 2018, 3, e92193. [Google Scholar] [CrossRef]
- Mitchell, E.; Gilbert, M.; Loomes, K.M. Alagille Syndrome. Clin. Liver Dis. 2018, 22, 625–641. [Google Scholar] [CrossRef]
- Wasson, C.W.; Abignano, G.; Hermes, H.; Malaab, M.; Ross, R.L.; Jimenez, S.A.; Chang, H.Y.; Feghali-Bostwick, C.A.; Del Galdo, F. Long non-coding RNA HOTAIR drives EZH2-dependent myofibroblast activation in systemic sclerosis through miRNA 34a-dependent activation of NOTCH. Ann. Rheum. Dis. 2020, 79, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, D.; Meng, N. Effects of RUNX3 mediated Notch signaling pathway on biological characteristics of colorectal cancer cells. Int. J. Oncol. 2017, 50, 2059–2068. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yang, X.; Jiang, A.; Wang, W.; Li, J.; Wen, J. Methylation-dependent transcriptional repression of RUNX3 by KCNQ1OT1 regulates mouse cardiac microvascular endothelial cell viability and inflammatory response following myocardial infarction. FASEB J. 2019, 33, 13145–13160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasson, C.W.; Ross, R.L.; Wells, R.; Corinaldesi, C.; Georgiou, I.C.; Riobo-Del Galdo, N.A.; Del Galdo, F. Long non-coding RNA HOTAIR induces GLI2 expression through Notch signalling in systemic sclerosis dermal fibroblasts. Arthritis Res. Ther. 2020, 22, 286. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sun, M.; Xia, R.; Zhang, E.; Liu, X.; Zhang, Z.; Xu, T.; De, W.; Liu, B.; Wang, Z. LincHOTAIR epigenetically silences miR34a by binding to PRC2 to promote the epithelial-to-mesenchymal transition in human gastric cancer. Cell Death Dis. 2015, 6, e1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Liu, H.Y.; Han, Y.L.; Wang, L.; Zhai, D.D.; Ma, T.; Zhang, M.J.; Liang, C.Z.; Shen, Y. Silence of lncRNA XIST represses myocardial cell apoptosis in rats with acute myocardial infarction through regulating MIR-449. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8566–8572. [Google Scholar] [CrossRef] [PubMed]
- Hadji, F.; Boulanger, M.C.; Guay, S.P.; Gaudreault, N.; Amellah, S.; Mkannez, G.; Bouchareb, R.; Marchand, J.T.; Nsaibia, M.J.; Guauque-Olarte, S.; et al. Altered DNA Methylation of Long Noncoding RNA H19 in Calcific Aortic Valve Disease Promotes Mineralization by Silencing NOTCH1. Circulation 2016, 134, 1848–1862. [Google Scholar] [CrossRef]
- Wang, J.; Cao, B.; Zhao, H.; Gao, Y.; Luo, Y.; Chen, Y.; Feng, J. Long noncoding RNA H19 prevents neurogenesis in ischemic stroke through p53/Notch1 pathway. Brain Res. Bull. 2019, 150, 111–117. [Google Scholar] [CrossRef]
- Wan, Y.; Yang, Z.Q. LncRNA NEAT1 affects inflammatory response by targeting miR-129-5p and regulating Notch signaling pathway in epilepsy. Cell Cycle 2020, 19, 419–431. [Google Scholar] [CrossRef]
- Wei, R.; Chen, Y.; Zhao, Z.; Gu, Q.; Wu, J. LncRNA FAM83H-AS1 induces nucleus pulposus cell growth via targeting the Notch signaling pathway. J. Cell. Physiol. 2019, 234, 22163–22171. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.B.; Tang, C.; Jin, X.; Liu, S.H.; Pi, W. Increased expression of lncRNA SNHG12 predicts a poor prognosis of nasopharyngeal carcinoma and regulates cell proliferation and metastasis by modulating Notch signal pathway. Cancer Biomark. 2018, 23, 603–613. [Google Scholar] [CrossRef]
- Chen, M.; Xia, Z.; Chen, C.; Hu, W.; Yuan, Y. LncRNA MALAT1 promotes epithelial-to-mesenchymal transition of esophageal cancer through Ezh2-Notch1 signaling pathway. Anti Cancer Drugs 2018, 29, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jin, X.; Wang, Z.; Zhang, X.; Liu, S.; Liu, G. Downregulation of SNHG1 suppresses cell proliferation and invasion by regulating Notch signaling pathway in esophageal squamous cell cancer. Cancer Biomark. 2017, 21, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zuo, J.J.; Li, F.; Xu, Y.C.; Zheng, A.Y.; Tao, Z.Z. LncRNA SNHG1 promotes cell proliferation in laryngeal cancer via Notch1 signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 12, 6562–6569. [Google Scholar] [CrossRef]
- Lu, S.; Dong, W.; Zhao, P.; Liu, Z. LncRNA FAM83H-AS1 is associated with the prognosis of colorectal carcinoma and promotes cell proliferation by targeting the notch signaling pathway. Oncol. Lett. 2018, 15, 1861–1868. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Yang, Z.; Long, F.; Luo, L.; Yang, B.; Zhu, R.; Sang, X.; Cao, G.; Wang, K. Long noncoding RNA MALAT1 mediates stem cell-like properties in human colorectal cancer cells by regulating miR-20b-5p/Oct4 axis. J. Cell. Physiol. 2019, 234, 20816–20828. [Google Scholar] [CrossRef]
- Zhang, H.F.; Li, W.; Han, Y.D. LINC00261 suppresses cell proliferation, invasion and Notch signaling pathway in hepatocellular carcinoma. Cancer Biomark. 2018, 21, 575–582. [Google Scholar] [CrossRef]
- Bai, L.; Wang, A.; Zhang, Y.; Xu, X.; Zhang, X. Knockdown of MALAT1 enhances chemosensitivity of ovarian cancer cells to cisplatin through inhibiting the Notch1 signaling pathway. Exp. Cell Res. 2018, 366, 161–171. [Google Scholar] [CrossRef]
- Guo, Q.; Qian, Z.; Yan, D.; Li, L.; Huang, L. LncRNA-MEG3 inhibits cell proliferation of endometrial carcinoma by repressing Notch signaling. Biomed. Pharmacother. 2016, 82, 589–594. [Google Scholar] [CrossRef]
- Wu, Z.; Huang, W.; Chen, Y.; Chen, B.; Liu, R.; Bai, P.; Xing, J. LINC01638 lncRNA promotes the proliferation, migration and invasion of prostate carcinoma cells by interacting with Notch1. Cancer Biomark. 2019, 25, 161–168. [Google Scholar] [CrossRef]
- Zhu, Y.; Tong, Y.; Wu, J.; Liu, Y.; Zhao, M. Knockdown of LncRNA GHET1 suppresses prostate cancer cell proliferation by inhibiting HIF-1α/Notch-1 signaling pathway via KLF2. BioFactors 2019, 45, 364–373. [Google Scholar] [CrossRef]
- Tang, L.X.; Su, S.F.; Wan, Q.; He, P.; Xhang, Y.; Cheng, X.M. Novel long non-coding RNA LBX2-AS1 indicates poor prognosis and promotes cell proliferation and metastasis through Notch signaling in non-small cell lung cancer. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7419–7429. [Google Scholar] [CrossRef]
- Qi, H.; Wang, S.; Wu, J.; Yang, S.; Gray, S.; Ng, C.S.H.; Du, J.; Underwood, M.J.; Li, M.Y.; Chen, G.G. EGFR-AS1/HIF2A regulates the expression of FOXP3 to impact the cancer stemness of smoking-related non-small cell lung cancer. Ther. Adv. Med. Oncol. 2019, 11, 1758835919855228. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhao, H.; Li, J.; Zhang, A.; Wang, H. Downregulation of long non-coding RNA LET predicts poor prognosis and increases Notch signaling in non-small cell lung cancer. Oncotarget 2018, 9, 1156–1168. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.Z.; Cai, L.; Li, B. MEG3 long non-coding RNA prevents cell growth and metastasis of osteosarcoma. Bratisl. Med. J. 2017, 118, 632–636. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Wu, B.; Li, Z.; He, P.; Wang, B.; Cai, A.; Zhang, X. LncRNA NBR2 inhibits epithelial-mesenchymal transition by regulating Notch1 signaling in osteosarcoma cells. J. Cell. Biochem. 2019, 120, 2015–2027. [Google Scholar] [CrossRef]
- Li, Z.; Tang, Y.; Xing, W.; Dong, W.; Wang, Z. LncRNA, CRNDE promotes osteosarcoma cell proliferation, invasion and migration by regulating Notch1 signaling and epithelial-mesenchymal transition. Exp. Mol. Pathol. 2018, 104, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Vander Roest, M.; Krapp, C.; Thorvaldsen, J.L.; Bartolomei, M.S.; Merryman, W.D. H19 is not hypomethylated or upregulated with age or sex in the aortic valves of mice. Physiol. Rep. 2019, 7, e14244. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Chen, L.; Tang, Y.; Yuan, P.; Yan, J.; Zheng, Y.; Huang, L.; Li, Z.; Sun, Y.; Han, S.; et al. Lnc-RP5 Regulates the miR-129-5p/Notch1/PFV Internal Promoter Axis to Promote the Expression of the Prototype Foamy Virus Transactivator Tas. Virol. Sin. 2019, 35, 73–82. [Google Scholar] [CrossRef]
- Zhang, K.; Han, X.; Zhang, Z.; Zheng, L.; Hu, Z.; Yao, Q.; Cui, H.; Shu, G.; Si, M.; Li, C.; et al. The liver-enriched lnc-LFAR1 promotes liver fibrosis by activating TGFβ and Notch pathways. Nat. Commun. 2017, 8, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Fei, T.; Verhaak, R.G.W.; Su, Z.; Zhang, Y.; Brown, M.; Chen, Y.; Liu, X.S. Integrative genomic analyses reveal clinically relevant long noncoding RNAs in human cancer. Nat. Struct. Mol. Biol. 2013, 20, 908–913. [Google Scholar] [CrossRef] [Green Version]
- Pardini, B.; Sabo, A.A.; Birolo, G.; Calin, G.A. Noncoding rnas in extracellular fluids as cancer biomarkers: The new frontier of liquid biopsies. Cancers 2019, 11, 1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeuschner, P.; Linxweiler, J.; Junker, K. Non-coding RNAs as biomarkers in liquid biopsies with a special emphasis on extracellular vesicles in urological malignancies. Expert Rev. Mol. Diagn. 2020, 20, 151–167. [Google Scholar] [CrossRef]
- Bolha, L.; Ravnik-Glavač, M.; Glavač, D. Long Noncoding RNAs as Biomarkers in Cancer. Dis. Markers 2017, 2017, 7243968. [Google Scholar] [CrossRef] [Green Version]
- Chandra Gupta, S.; Nandan Tripathi, Y. Potential of long non-coding RNAs in cancer patients: From biomarkers to therapeutic targets. Int. J. Cancer 2017, 140, 1955–1967. [Google Scholar] [CrossRef]
- Deng, J.; Tang, J.; Wang, G.; Zhu, Y.S. Long non-coding RNA as potential biomarker for prostate cancer: Is it making a difference? Int. J. Environ. Res. Public Health 2017, 14, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCleland, M.L.; Mesh, K.; Lorenzana, E.; Chopra, V.S.; Segal, E.; Watanabe, C.; Haley, B.; Mayba, O.; Yaylaoglu, M.; Gnad, F.; et al. CCAT1 is an enhancer-templated RNA that predicts BET sensitivity in colorectal cancer. J. Clin. Investig. 2016, 126, 639–652. [Google Scholar] [CrossRef] [Green Version]
- Barrows, J.K.; Lin, B.; Quaas, C.E.; Fullbright, G.; Wallace, E.N.; Long, D.T. BRD4 promotes resection and homology-directed repair of DNA double-strand breaks. Nat. Commun. 2022, 13, 3016. [Google Scholar] [CrossRef] [PubMed]
- Donati, B.; Lorenzini, E.; Ciarrocchi, A. BRD4 and Cancer: Going beyond transcriptional regulation. Mol. Cancer 2018, 17, 164. [Google Scholar] [CrossRef]
- Kulikowski, E.; Rakai, B.D.; Wong, N.C.W. Inhibitors of bromodomain and extra-terminal proteins for treating multiple human diseases. Med. Res. Rev. 2021, 41, 223–245. [Google Scholar] [CrossRef]
- Hang, Q.; Sun, R.; Jiang, C.; Li, Y. Notch 1 promotes cisplatin-resistant gastric cancer formation by upregulating lncRNA AK022798 expression. Anticancer Drugs 2015, 26, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zheng, M.; Wang, H. Circular RNA hsa_circ_0072309 inhibits proliferation and invasion of breast cancer cells via targeting miR-492. Cancer Manag. Res. 2019, 11, 1033–1041. [Google Scholar] [CrossRef] [Green Version]
- Smid, M.; Wilting, S.M.; Uhr, K.; Rodríguez-González, F.G.; De Weerd, V.; Prager-Van Der Smissen, W.J.C.; Van Der Vlugt-Daane, M.; Van Galen, A.; Nik-Zainal, S.; Butler, A.; et al. The circular RNome of primary breast cancer. Genome Res. 2019, 29, 356–366. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, Y.; Qi, L.; Ding, L.; Jiang, H.; Yu, H. NFIX Circular RNA Promotes Glioma Progression by Regulating miR-34a-5p via Notch Signaling Pathway. Front. Mol. Neurosci. 2018, 11, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Li, Z.; Zhang, M.; Wang, B.; Ye, J.; Zhang, Y.; Tang, D.; Ma, D.; Jin, W.; Li, X.; et al. Circ-ASH2L promotes tumor progression by sponging miR-34a to regulate Notch1 in pancreatic ductal adenocarcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, F.; Maillard, I. Therapeutic Targeting of Notch Signaling: From Cancer to Inflammatory Disorders. Front. Cell Dev. Biol. 2021, 9, 649205. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kałafut, J.; Czerwonka, A.; Czapla, K.; Przybyszewska-Podstawka, A.; Hermanowicz, J.M.; Rivero-Müller, A.; Borkiewicz, L. Regulation of Notch1 Signalling by Long Non-Coding RNAs in Cancers and Other Health Disorders. Int. J. Mol. Sci. 2023, 24, 12579. https://doi.org/10.3390/ijms241612579
Kałafut J, Czerwonka A, Czapla K, Przybyszewska-Podstawka A, Hermanowicz JM, Rivero-Müller A, Borkiewicz L. Regulation of Notch1 Signalling by Long Non-Coding RNAs in Cancers and Other Health Disorders. International Journal of Molecular Sciences. 2023; 24(16):12579. https://doi.org/10.3390/ijms241612579
Chicago/Turabian StyleKałafut, Joanna, Arkadiusz Czerwonka, Karolina Czapla, Alicja Przybyszewska-Podstawka, Justyna Magdalena Hermanowicz, Adolfo Rivero-Müller, and Lidia Borkiewicz. 2023. "Regulation of Notch1 Signalling by Long Non-Coding RNAs in Cancers and Other Health Disorders" International Journal of Molecular Sciences 24, no. 16: 12579. https://doi.org/10.3390/ijms241612579