
Genome-Wide Identification and Expression Analysis of the Sucrose Synthase Gene Family in Sweet Potato and Its Two Diploid Relatives



Abstract
:1. Introduction
2. Results
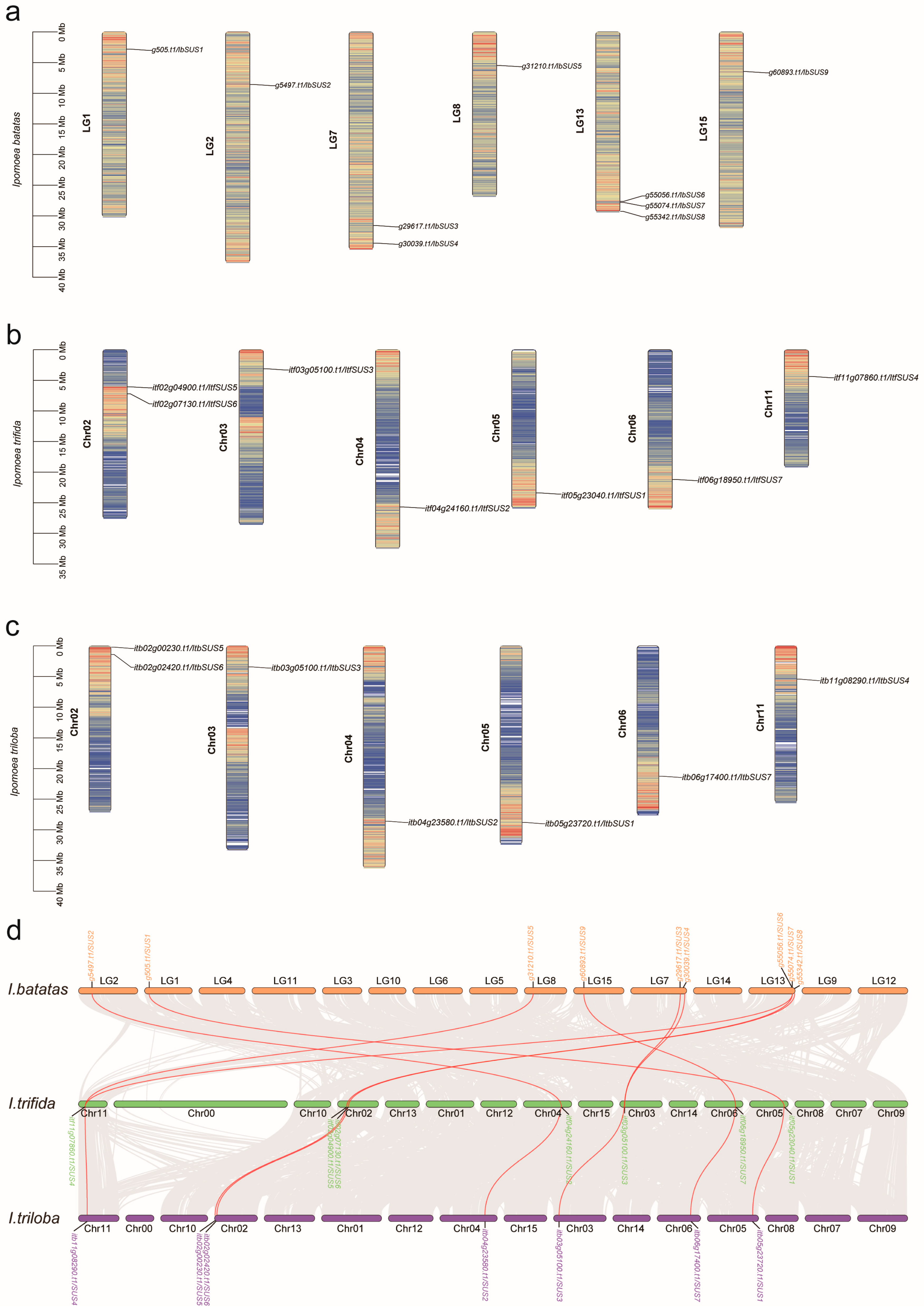
2.1. Identification of SUSs in Sweet Potato and Its Two Diploid Wild Relatives
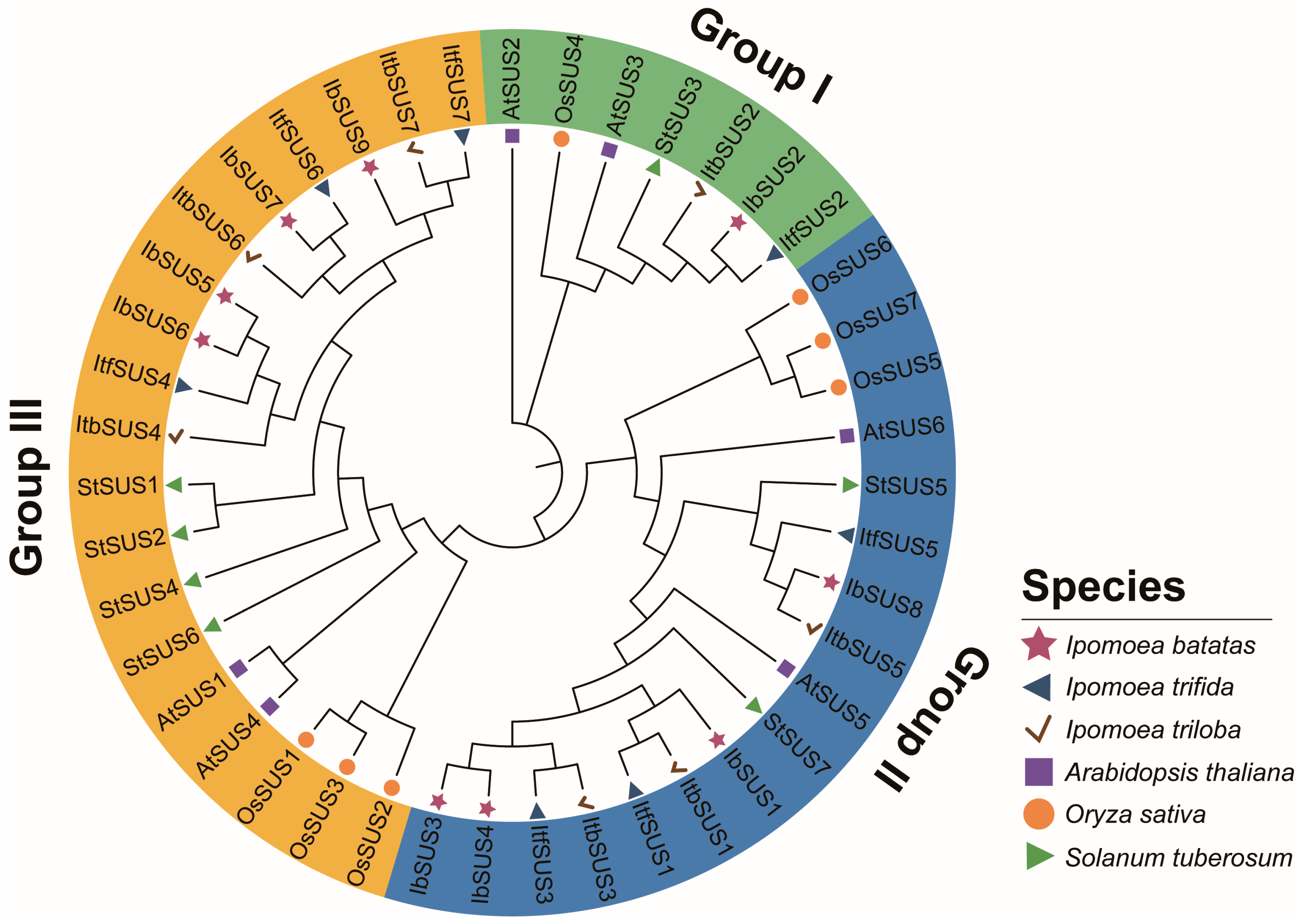
2.2. Phylogenetic Relationship Analysis of SUSs in Sweet Potato and Its Two Diploid Wild Relatives
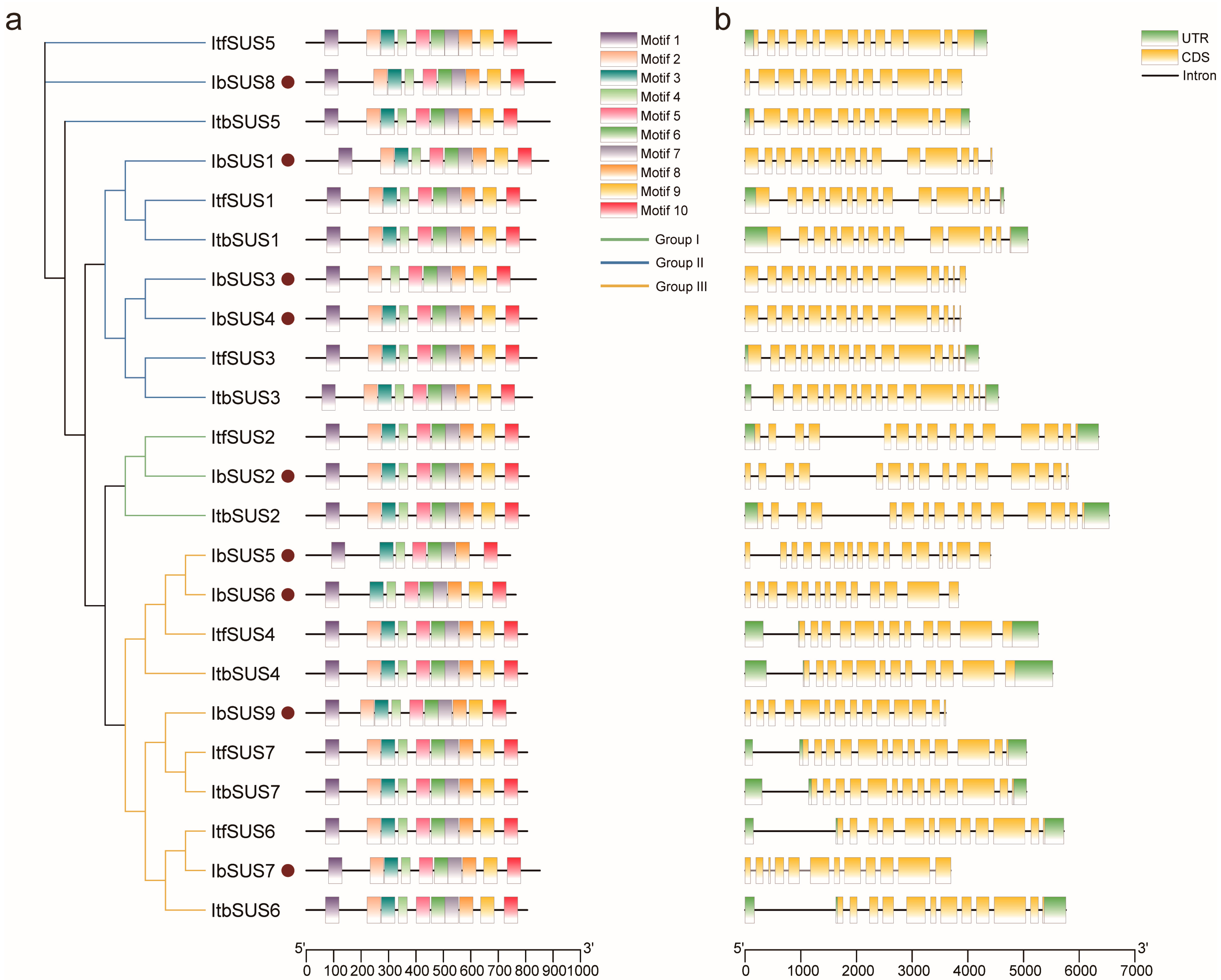
2.3. Conserved Motif and Exon–Intron Structure Analysis of SUSs in Sweet Potato and Its Two Diploid Wild Relatives
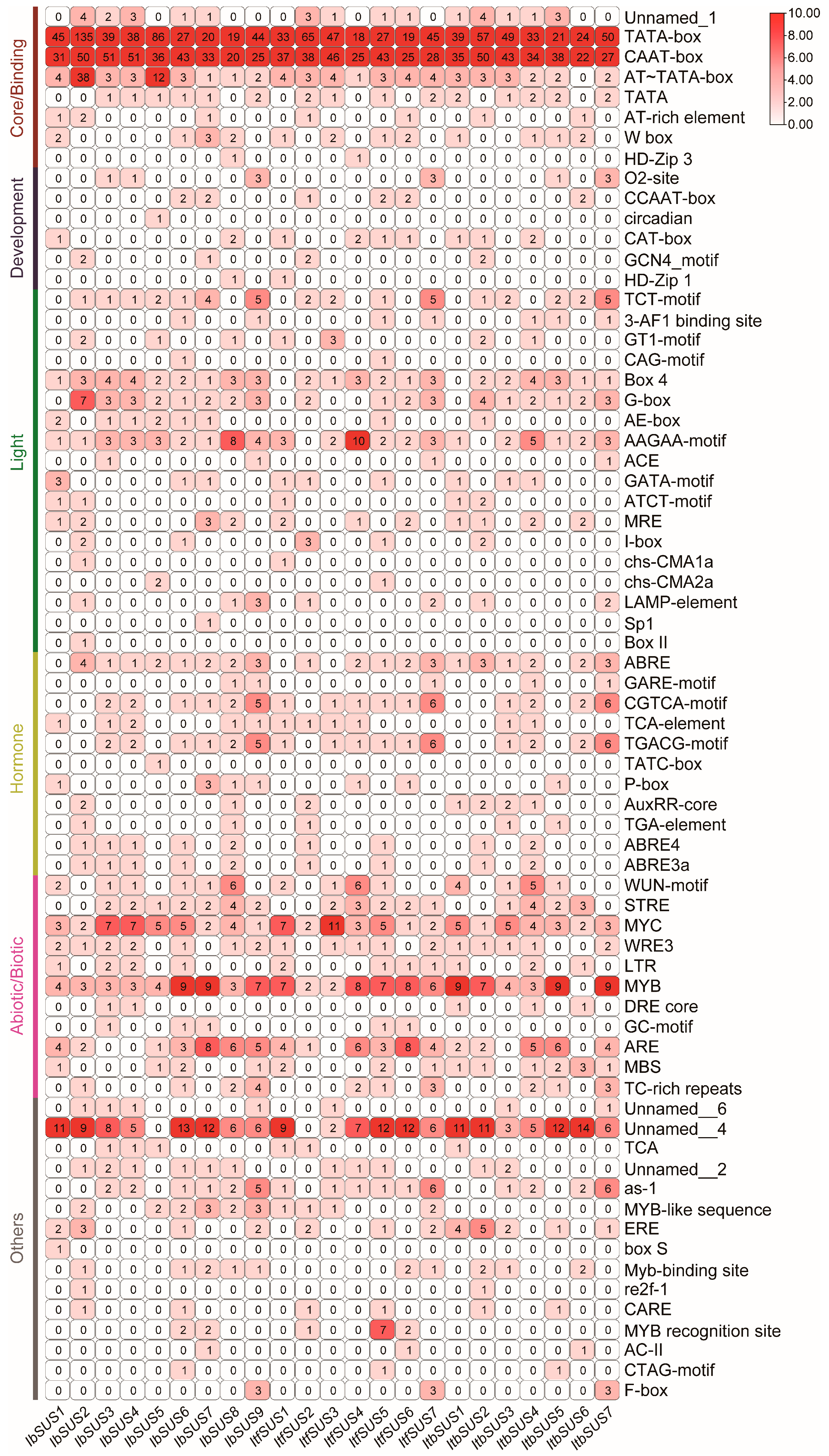
2.4. Cis-Element Analysis in the Promoters of SUSs in Sweet Potato and Its Two Diploid Wild Relatives
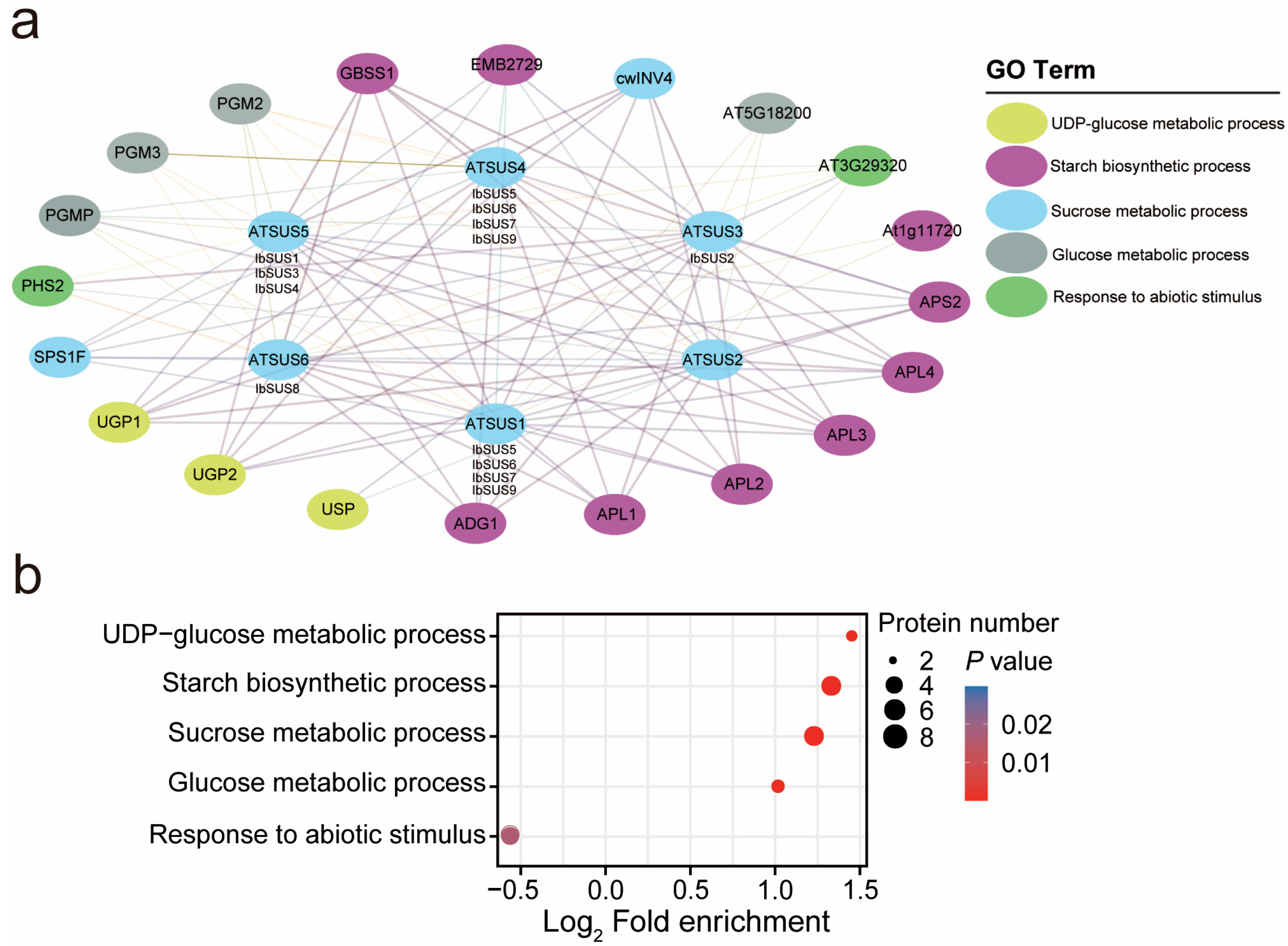
2.5. Protein Interaction Network of IbSUSs in Sweet Potato
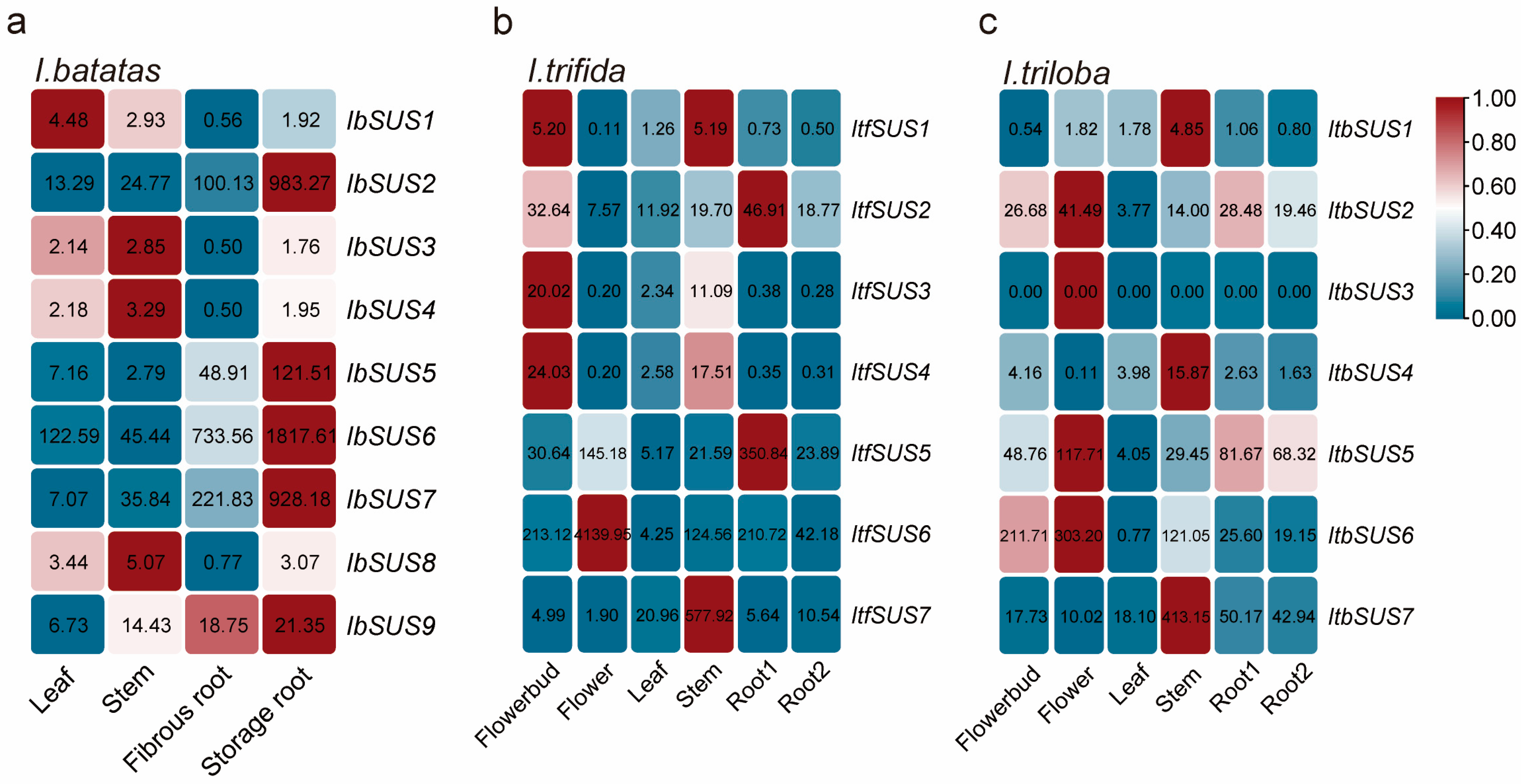
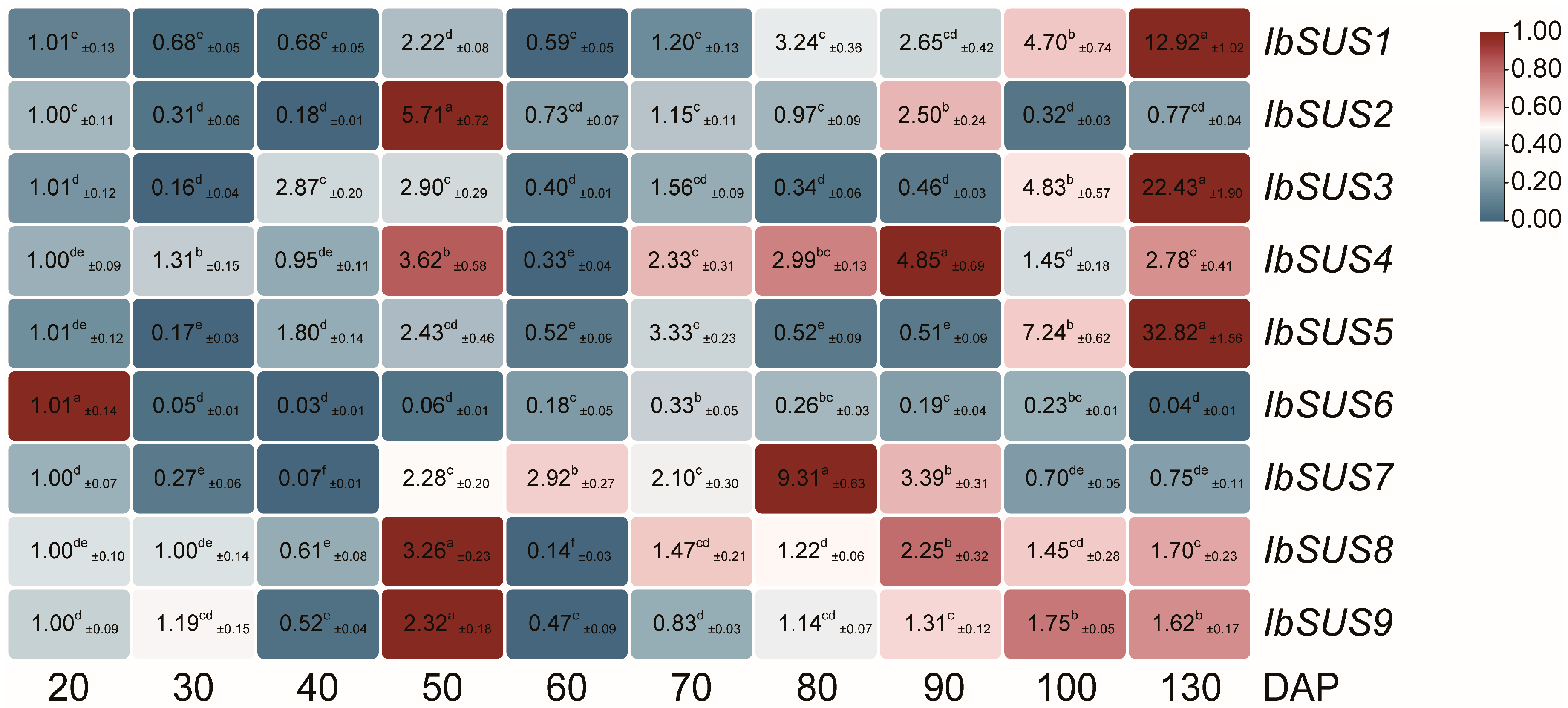
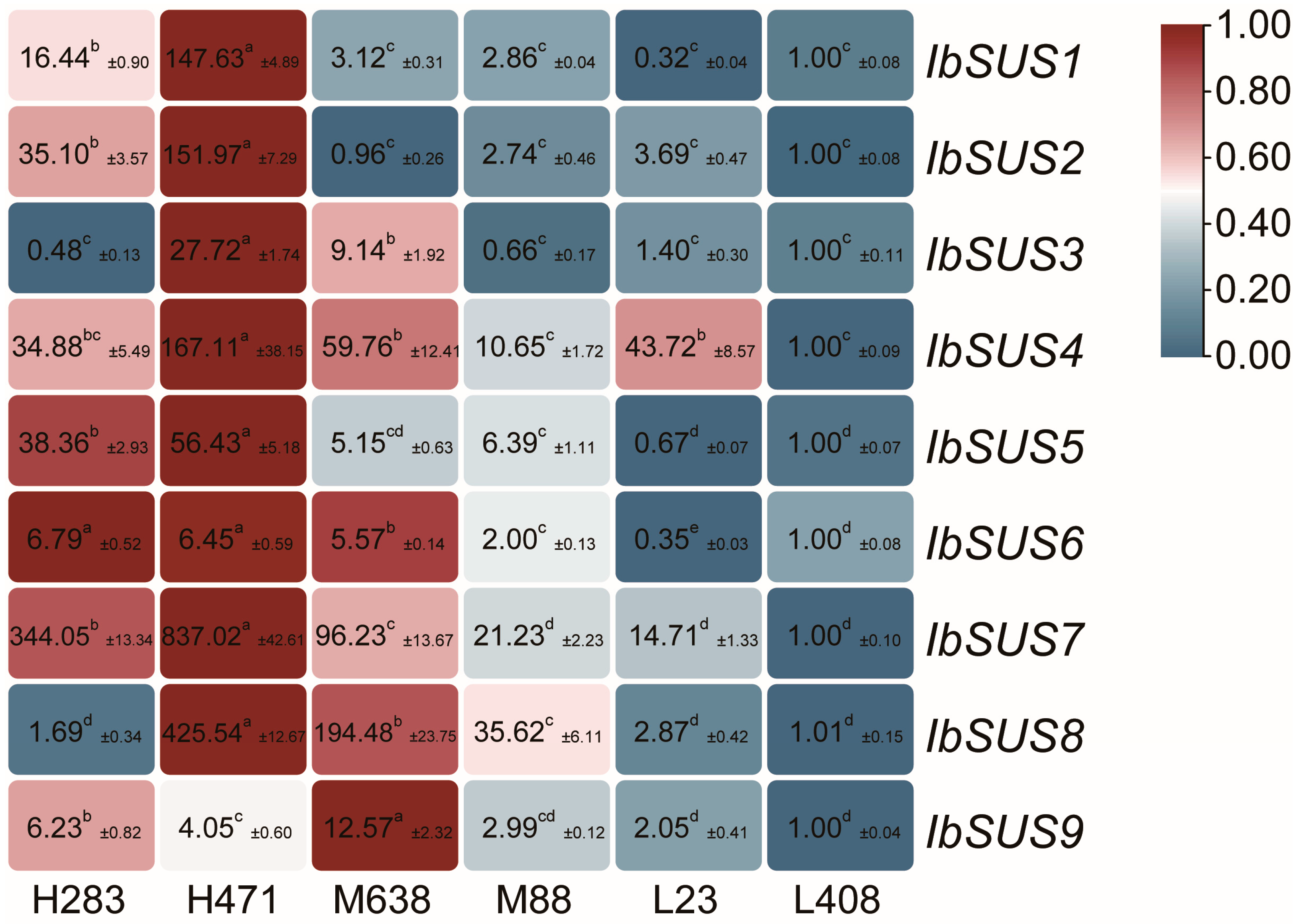
2.6. Expression Analysis of SUSs in Sweet Potato and Its Two Diploid Wild Relatives
2.6.1. Expression Analysis in Various Tissues
2.6.2. Expression Analysis at Different Developmental Stages of Storage Roots in Sweet Potato
2.6.3. Expression Analysis in Sweet Potato Lines with Different Starch Contents
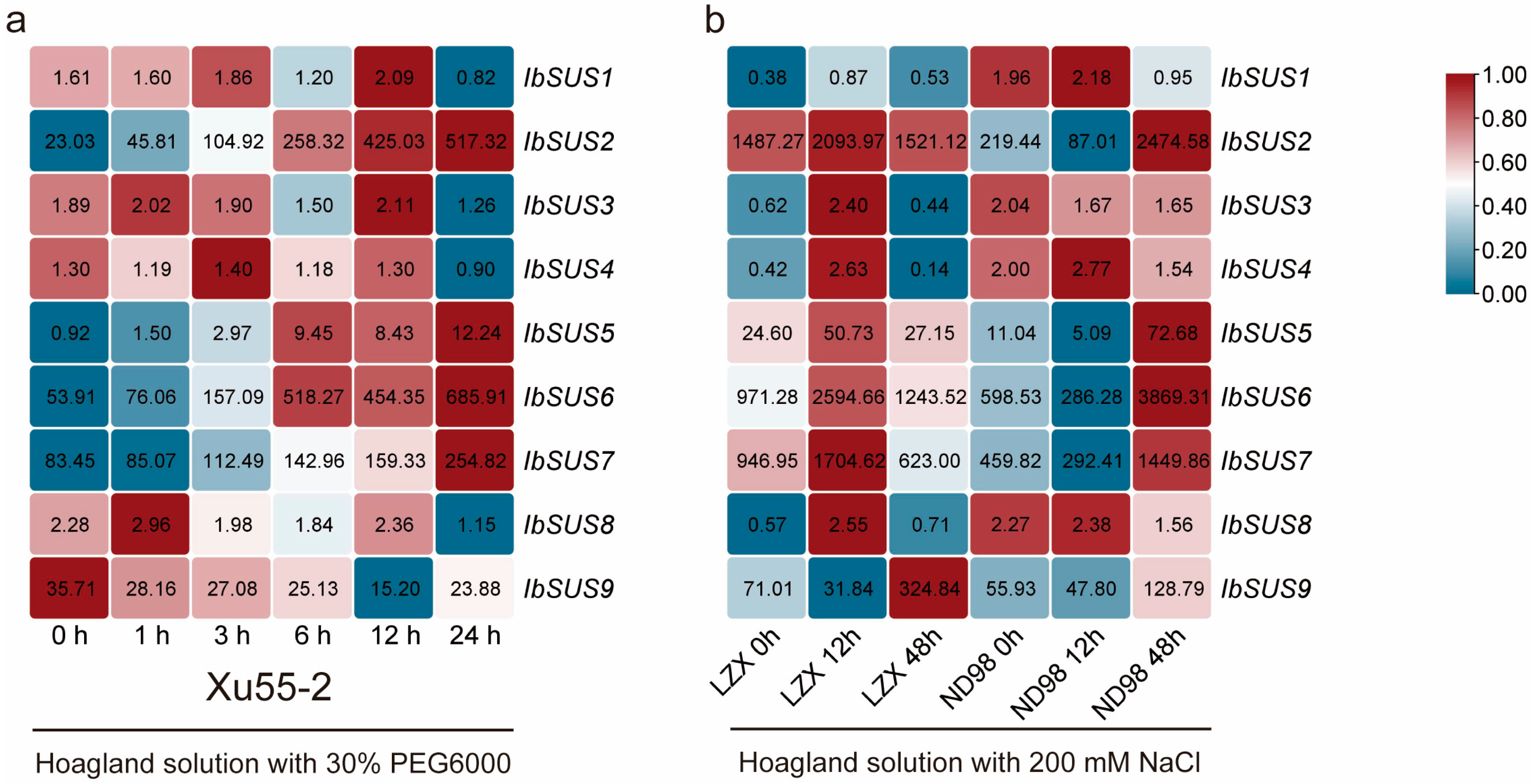
2.6.4. Expression Analysis under Drought and Salt Stresses
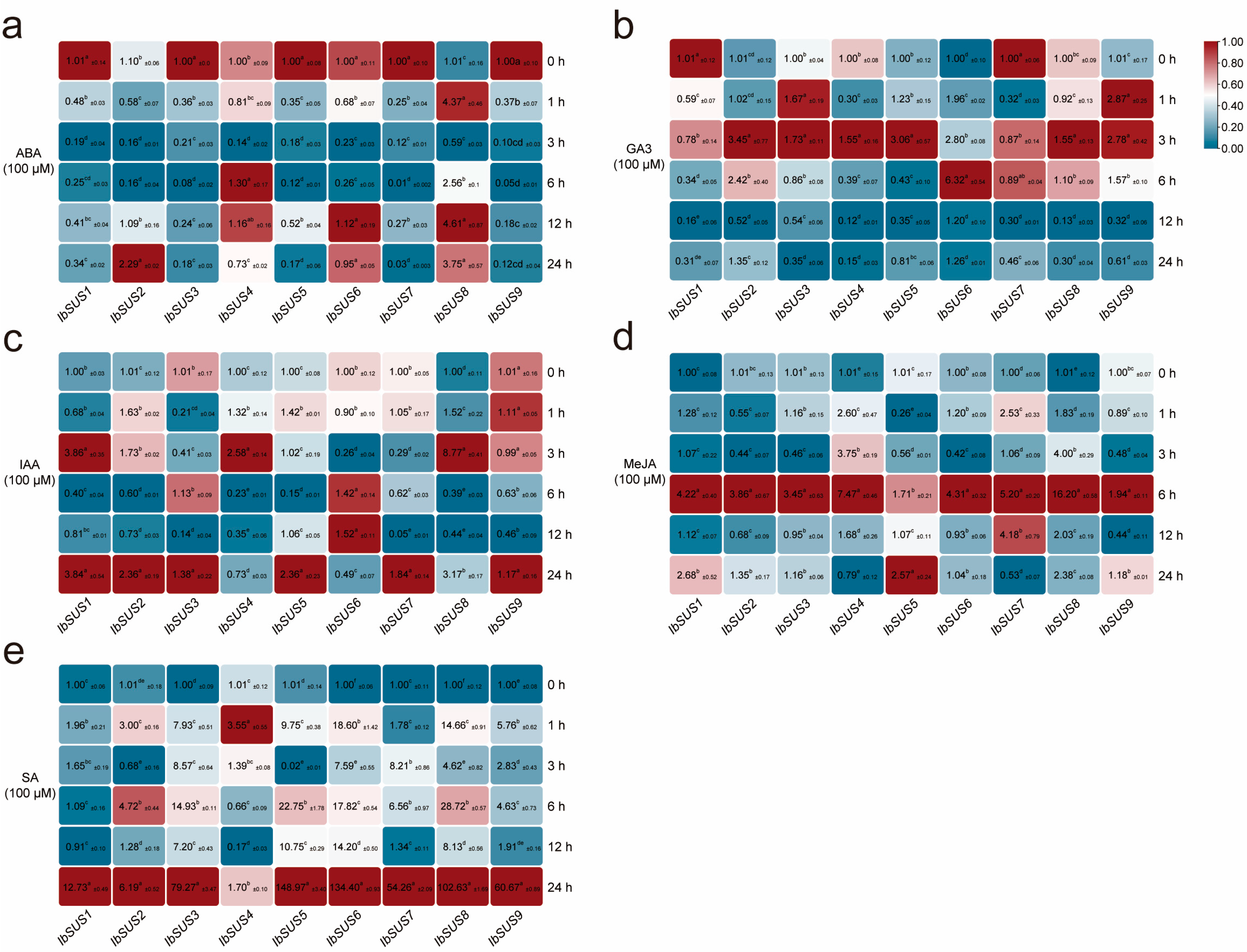
2.6.5. Expression Analysis in Response to Hormones
3. Discussion
3.1. Evolution of SUSs in Sweet Potato and Its Two Diploid Wild Relatives
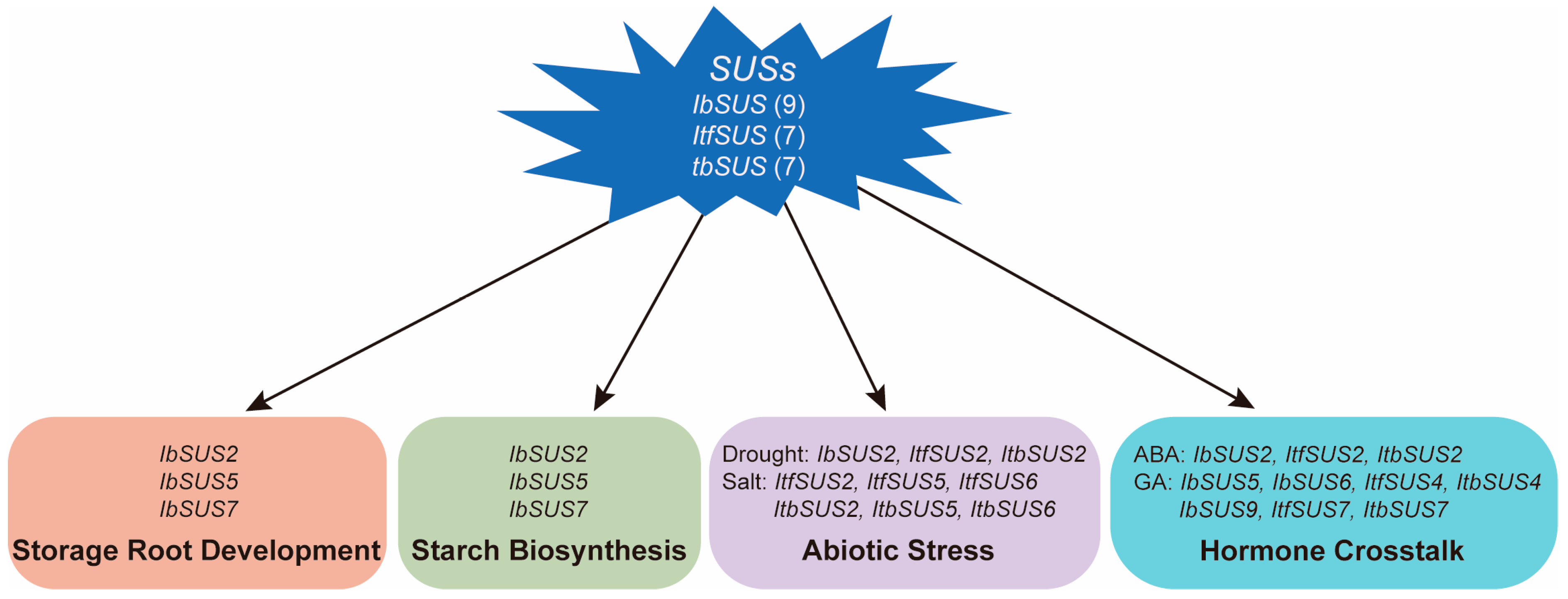
3.2. IbSUSs Are Involved in Storage Root Development and Starch Biosynthesis in Sweet Potato
3.3. SUSs Regulate Response to Drought and Salt Stresses in Sweet Potato and Its Two Diploid Wild Relatives
3.4. SUSs Participate in Hormone Crosstalk in Sweet Potato and Its Two Diploid Wild Relatives
4. Materials and Methods
4.1. Identification of SUSs
4.2. Chromosomal Distribution of SUSs
4.3. Property Prediction of SUSs
4.4. Phylogenetic Analysis of SUSs
4.5. Domain Identification and Conserved Motif Analysis of SUSs
4.6. Exon–Intron Structure and Promoter Analyses of SUSs
4.7. Protein Interaction Network and GO Enrichment Analysis of SUSs and Regulated Proteins
4.8. Transcriptome Analysis
4.9. qRT-PCR Analysis of SUSs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruan, Y.L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.D.; Wang, M.Q.; Li, X.P.; Jiu, S.T.; Wang, C.; Fang, J.G. Genome-wide analysis of the sucrose synthase gene family in grape (Vitis vinifera): Structure, evolution and expression profiles. Genes 2017, 8, 111. [Google Scholar] [CrossRef] [Green Version]
- Stein, O.; Granot, D. An overview of sucrose synthases in plants. Front. Plant Sci. 2019, 10, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, D.Y.; Gonzales-Vigil, E.; Mansfield, S.D. Arabidopsis sucrose synthase localization indicates a primary role in sucrose translocation in phloem. J. Exp. Bot. 2020, 71, 1858–1869. [Google Scholar] [CrossRef]
- Mathan, J.; Singh, A.; Ranjan, A. Sucrose transport and metabolism control carbon partitioning between stem and grain in rice. J. Exp. Bot. 2021, 72, 4355–4372. [Google Scholar] [CrossRef]
- Koch, K. Sucrose metabolism: Regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr. Opin. Plant Biol. 2004, 7, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.D.H.; Paul, D.; Kim, F.; Marilyn, P.; Wellner, N.; Lunn, J.; Feil, R.; Simpsona, C.; Maule, A.J.; Smith, A.M. Normal growth of Arabidopsis requires cytosolic invertase but not sucrose synthase. Proc. Natl. Acad. Sci. USA 2009, 106, 13124–13129. [Google Scholar]
- Zheng, Y.; Anderson, S.; Zhang, Y.; Garavito, R.M. The structure of sucrose synthase-1 from Arabidopsis thaliana and its functional implications. J. Biol. Chem. 2011, 286, 36108–36118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmolzer, K.; Gutmann, A.; Diricks, M.; Desmet, T.; Nidetzky, B. Sucrose synthase: A unique glycosyltransferase for biocatalytic glycosylation process development. Biotechnol. Adv. 2016, 34, 88–111. [Google Scholar] [CrossRef]
- Angeles-Núñez, J.G.; Tiessen, A. Regulation of ATSUS2 and ATSUS3 by glucose and the transcription factor LEC2 in different tissues and at different stages of arabidopsis seed development. Plant Mol. Biol. 2012, 78, 377–392. [Google Scholar] [CrossRef]
- Baud, S.; Vaultier, M.N.; Rochat, C. Structure and expression profile of the sucrose synthase multigene family in Arabidopsis. J. Exp. Bot. 2004, 55, 397–409. [Google Scholar] [CrossRef]
- Duncan, K.A.; Hardin, S.C.; Huber, S.C. The three maize sucrose synthase isoforms differ in distribution, localization, and phosphorylation. Plant Cell Physiol. 2006, 47, 959–971. [Google Scholar] [CrossRef] [Green Version]
- Hirose, T.; Scofield, G.N.; Terao, T. An expression analysis profile for the entire sucrose synthase gene family in rice. Plant Sci. 2008, 174, 534–543. [Google Scholar] [CrossRef]
- Zou, C.; Lu, C.; Shang, H.; Jing, X.; Cheng, H.; Zhang, Y.; Song, G. Genome-wide analysis of the Sus gene family in cotton. J. Integr. Plant Biol. 2013, 55, 643–653. [Google Scholar] [CrossRef]
- An, X.; Chen, Z.; Wang, J.; Ye, M.; Ji, L.; Wang, J.; Liao, W.; Ma, H. Identification and characterization of the Populus sucrose synthase gene family. Gene 2014, 539, 58–67. [Google Scholar] [CrossRef]
- Fu, H.; Park, W.D. Sink- and vascular-associated sucrose synthase functions are encoded by different gene classes in potato. Plant Cell 1995, 7, 1369–1385. [Google Scholar]
- Biemelt, S.; Hajirezaei, M.R.; Melzer, M.; Albrecht, G.; Sonnewald, U. Sucrose synthase activity does not restrict glycolysis in roots of transgenic potato plants under hypoxic conditions. Planta 1999, 210, 41–49. [Google Scholar] [CrossRef]
- Chourey, P.S.; Taliercio, E.W.; Carlson, S.J.; Ruan, Y.L. Genetic evidence that the two isozymes of sucrose synthase present in developing maize endosperm are critical, one for cell wall integrity and the other for starch biosynthesis. Mol. Gen. Genet. 1998, 259, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Ricard, B.; Toai, T.V.; Chourey, P.; Saglio, P. Evidence for the critical role of sucrose synthase for anoxic tolerance of maize roots using a double mutant. Plant Physiol. 1998, 116, 1323–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.T.; Wang, J.C.; Zhang, Z.Y.; Wu, Y.R. Transactivation of Sus1 and Sus2 by Opaque2 is an essential supplement to sucrose synthase-mediated endosperm filling in maize. Plant Biotechnol. J. 2020, 18, 1897–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fallahi, H.; Scofield, G.N.; Badger, M.R.; Chow, W.S.; Furbank, R.T.; Ruan, Y.L. Localization of sucrose synthase in developing seed and siliques of Arabidopsis thaliana reveals diverse roles for SUS during development. J. Exp. Bot. 2008, 59, 3283–3295. [Google Scholar] [CrossRef] [Green Version]
- Baroja-Fernández, E.; Muñoz, F.J.; Li, J.; Bahaji, A.; Almagro, G.; Montero, M.; Etxeberria, E.; Hidalgo, M.; Sesma, M.T.; Pozueta-Romero, J. Sucrose synthase activity in the sus1/sus2/sus3/sus4 Arabidopsis mutant is sufficient to support normal cellulose and starch production. Proc. Natl. Acad. Sci. USA 2012, 109, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Piro, L.; Flütsch, S.; Santelia, D. Arabidopsis Sucrose Synthase 3 (SUS3) regulates starch accumulation in guard cells at the end of day. Plant Signal. Behav. 2023, 18, e2171614. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.M.; Brill, E.; Llewellyn, D.J.; Furbank, R.T.; Ruan, Y.L. Overexpression of a potato sucrose synthase gene in cotton accelerates leaf expansion, reduces seed abortion, and enhances fiber production. Mol. Plant 2012, 5, 430–441. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, Y.; Cheng, F.; Zhang, S.P.; Zheng, Y.; Li, Y.; Li, X.B. Comparative phosphoproteomic analysis reveals that phosphorylation of sucrose synthase GhSUS2 by Ca2+-dependent protein kinases GhCPK84/93 affects cotton fiber development. J. Exp. Bot. 2023, 74, 1836–1852. [Google Scholar] [CrossRef] [PubMed]
- Takehara, K.; Murata, K.; Yamaguchi, T.; Yamaguchi, K.; Chaya, G.; Kido, S.; Iwasaki, Y.; Ogiwara, H.; Ebitani, T.; Miura, K. Thermo-responsive allele of sucrose synthase 3 (Sus3) provides high temperature tolerance during the ripening stage in rice (Oryza sativa L.). Breed. Sci. 2018, 68, 336–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Wang, L.L.; Hu, S.L.; Luo, X.G.; Cao, Y. Overexpression of the bamboo sucrose synthase gene (BeSUS5) improves cellulose production, cell wall thickness and fiber quality in transgenic poplar. Tree Genet. Genomes 2020, 16, 75. [Google Scholar] [CrossRef]
- Liu, Q.C. Improvement for agronomically important traits by gene engineering in sweetpotato. Breed. Sci. 2017, 67, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, Q.; Zhai, H.; Gao, S.; Yang, L.; Wang, Z.; Xu, Y.; Huo, J.; Ren, Z.; Zhao, N.; et al. IbBBX24 promotes the jasmonic acid pathway and enhances fusarium wilt resistance in sweet potato. Plant Cell 2020, 32, 1102–1123. [Google Scholar] [CrossRef]
- Xue, L.Y.; Wei, Z.H.; Zhai, H.; Xing, S.H.; Wang, Y.X.; He, S.Z.; Gao, S.P.; Zhao, N.; Zhang, H.; Liu, Q.C. The IbPYL8-IbbHLH66-IbbHLH118 complex mediates the abscisic acid-dependent drought response in sweet potato. New Phytol. 2022, 236, 2151–2171. [Google Scholar] [CrossRef]
- Lai, Y.C.; Wang, S.Y.; Gao, H.Y.; Nguyen, K.M.; Nguyen, C.H.; Shih, M.C.; Lin, K.H. Physicochemical properties of starches and expression and activity of starch biosynthesis-related genes in sweet potatoes. Food Chem. 2016, 199, 556–564. [Google Scholar] [CrossRef]
- Yang, J.; Moeinzadeh, M.H.; Kuhl, H.; Helmuth, J.; Xiao, P.; Haas, S.; Liu, G.; Zheng, J.; Sun, Z.; Fan, W.; et al. Haplotype-resolved sweet potato genome traces back its hexaploidization history. Nat. Plants 2017, 3, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Lau, K.H.; Cao, Q.; Hamilton, J.P.; Sun, H.; Zhou, C.; Eserman, L.; Gemenet, D.C.; Olukolu, B.A.; Wang, H.; et al. Genome sequences of two diploid wild relatives of cultivated sweetpotato reveal targets for genetic improvement. Nat. Commun. 2018, 9, 4580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Xin, S.; Huang, Z.; Zhu, Z.; Si, C.; Chen, Y.; Zhu, G.; Liu, Y. Identification of key sucrose-degrading enzymes and the relevant gene family members in the development of sweetpotato storage roots. Chin. J. Trop. Crops 2022, 43, 1535–1544. [Google Scholar]
- Zheng, C.X.; Jiang, Z.C.; Meng, Y.S.; Yu, J.; Yang, X.S.; Zhang, H.; Zhao, N.; He, S.Z.; Gao, S.P.; Zhai, H.; et al. Construction of a high-density SSR genetic linkage map and identification of QTL for storage-root yield and dry-matter content in sweetpotato. Crop J. 2023, 11, 963–967. [Google Scholar] [CrossRef]
- Zhu, H.; Zhou, Y.Y.; Zhai, H.; He, S.Z.; Zhao, N.; Liu, Q.C. Transcriptome profiling reveals insights into the molecular mechanism of drought tolerance in sweetpotato. J. Integr. Agric. 2019, 18, 9–23. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Q.; Zhai, H.; Li, Y.; Wang, X.; Liu, Q.; He, S. Transcript profile analysis reveals important roles of jasmonic acid signalling pathway in the response of sweet potato to salt stress. Sci. Rep. 2017, 7, 202–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Frugoli, J.A.; McPeek, M.A.; Thomas, T.L.; McClung, C.R. Intron loss and gain during evolution of the catalase gene family in angiosperms. Genetics 1998, 149, 355–365. [Google Scholar] [CrossRef]
- Lecharny, A.; Boudet, N.; Gy, I.; Aubourg, S.; Kreis, M. Introns in, introns out in plant gene families: A genomic approach of the dynamics of gene structure. J. Struct. Funct. Genom. 2003, 3, 111–116. [Google Scholar] [CrossRef]
- Zrenner, R.; Salanoubat, M.; Willmitzer, L.; Sonnewald, U. Evidence of the crucial role of sucrose synthase for sink strength using transgenic potato plants (Solanum tuberosum L.). Plant J. 1995, 7, 97–107. [Google Scholar] [CrossRef]
- Baroja-Fernández, E.; Muñoz, F.J.; Montero, M.; Etxeberria, E.; Sesma, M.T.; Ovecka, M.; Bahaji, A.; Ezquer, I.; Li, J.; Prat, S.; et al. Enhancing sucrose synthase activity in transgenic potato (Solanum tuberosum L.) tubers results in increased levels of starch, ADP glucose and UDP glucose and total yield. Plant Cell Physiol. 2009, 50, 1651–1662. [Google Scholar] [CrossRef] [PubMed]
- D’Aoust, M.A.; Yelle, S.; Nguyen-Quoc, B. Antisense inhibition of tomato fruit sucrose synthase decreases fruit setting and the sucrose unloading capacity of young fruit. Plant Cell 1999, 11, 2407–2418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.W.; Wang, H.Y.; Li, X.; Sui, X.L.; Zhang, Z.X. Downregulating cucumber sucrose synthase 4 (CsSUS4) suppresses the growth and development of flowers and fruits. Plant Cell Physiol. 2018, 60, 752–764. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.H.; Tang, C.R.; Fang, Y.J.; Yang, M.; Zhou, B.H.; Qi, J.; Zhang, Y. Structure and expression profile of the sucrose synthase gene family in the rubber tree: Indicative of roles in stress response and sucrose utilization in the laticifers. FEBS J. 2014, 281, 291–305. [Google Scholar] [CrossRef] [Green Version]
- Barrero, S.C.; Hernando, A.S.; Gonzalez, M.P.; Carbonero, P. Structure, expression profile and subcellular localisation of four different sucrose synthase genes from barley. Planta 2011, 234, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Goren, S.; Lugassi, N.; Stein, O.; Yeselson, Y.; Schaffer, A.A.; David-Schwartz, R.; Granot, D. Suppression of sucrose synthase affects auxin signaling and leaf morphology in tomato. PLoS ONE 2017, 12, e0182334. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis Tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Kohl, M.; Wiese, S.; Warscheid, B. Cytoscape: Software for visualization and analysis of biological networks. Methods Mol. Biol. 2011, 696, 291–303. [Google Scholar] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C-T method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Genomic Length (bp) | CDS (bp) | Protein Size (aa) | MW (kDa) | pI | Instability | GRAVY | Subcellular Location |
|---|---|---|---|---|---|---|---|---|---|
| g505 | IbSUS1 | 4689 | 2649 | 882 | 99.99 | 6.21 | 40.26 | −0.312 | cytoplasm |
| g5497 | IbSUS2 | 6116 | 2436 | 811 | 92.13 | 6.02 | 38.17 | −0.275 | chloroplast |
| g29617 | IbSUS3 | 4187 | 2514 | 837 | 94.75 | 6.14 | 36.59 | −0.351 | cytoplasm |
| g30039 | IbSUS4 | 4091 | 2520 | 839 | 94.95 | 6.33 | 38.73 | −0.33 | cytoplasm |
| g31210 | IbSUS5 | 4954 | 2232 | 743 | 84.78 | 6.11 | 36.16 | −0.202 | plastid |
| g55056 | IbSUS6 | 4794 | 2292 | 763 | 87.82 | 6.17 | 33.53 | −0.298 | mitochondria |
| g55074 | IbSUS7 | 5591 | 2556 | 851 | 97.49 | 6.16 | 38.91 | −0.256 | mitochondria |
| g55342 | IbSUS8 | 4125 | 2721 | 906 | 101.58 | 6.99 | 39.82 | −0.214 | chloroplast |
| g60893 | IbSUS9 | 4931 | 2292 | 763 | 87.7 | 6.18 | 35.82 | −0.261 | cytoplasm |
| itf05g23040 | ItfSUS1 | 4648 | 2511 | 836 | 94.86 | 6.02 | 38.89 | −0.326 | cytoplasm |
| itf04g24160 | ItfSUS2 | 6346 | 2436 | 811 | 92.21 | 5.99 | 37.62 | −0.281 | chloroplast |
| itf03g05100 | ItfSUS3 | 4194 | 2520 | 839 | 94.88 | 6.22 | 39.2 | −0.336 | cytoplasm |
| itf11g07860 | ItfSUS4 | 5265 | 2418 | 805 | 92.60 | 5.93 | 34.11 | −0.279 | cytoplasm |
| itf02g04900 | ItfSUS5 | 4342 | 2679 | 892 | 99.89 | 6.40 | 38.81 | −0.248 | chloroplast |
| itf02g07130 | ItfSUS6 | 5723 | 2418 | 805 | 92.49 | 5.95 | 35.03 | −0.25 | cytoplasm |
| itf06g18950 | ItfSUS7 | 5052 | 2418 | 805 | 92.71 | 5.96 | 33.65 | −0.27 | cytoplasm |
| itb05g23720 | ItbSUS1 | 5077 | 2508 | 835 | 94.88 | 6.10 | 38.94 | −0.332 | cytoplasm |
| itb04g23580 | ItbSUS2 | 6536 | 2436 | 811 | 92.09 | 6.02 | 38 | −0.277 | chloroplast |
| itb03g05100 | ItbSUS3 | 4547 | 2472 | 823 | 93.27 | 6.30 | 38.87 | −0.325 | chloroplast |
| itb11g08290 | ItbSUS4 | 5524 | 2418 | 805 | 92.58 | 6.02 | 33.8 | −0.271 | cytoplasm |
| itb02g00230 | ItbSUS5 | 4028 | 2664 | 887 | 99.29 | 6.51 | 37.07 | −0.236 | chloroplast |
| itb02g02420 | ItbSUS6 | 5760 | 2418 | 805 | 92.46 | 5.93 | 35.3 | −0.241 | cytoplasm |
| itb06g17400 | ItbSUS7 | 5052 | 2418 | 805 | 92.74 | 5.96 | 33.76 | −0.267 | cytoplasm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Z.; Zhang, H.; Gao, S.; Zhai, H.; He, S.; Zhao, N.; Liu, Q. Genome-Wide Identification and Expression Analysis of the Sucrose Synthase Gene Family in Sweet Potato and Its Two Diploid Relatives. Int. J. Mol. Sci. 2023, 24, 12493. https://doi.org/10.3390/ijms241512493
Jiang Z, Zhang H, Gao S, Zhai H, He S, Zhao N, Liu Q. Genome-Wide Identification and Expression Analysis of the Sucrose Synthase Gene Family in Sweet Potato and Its Two Diploid Relatives. International Journal of Molecular Sciences. 2023; 24(15):12493. https://doi.org/10.3390/ijms241512493
Chicago/Turabian StyleJiang, Zhicheng, Huan Zhang, Shaopei Gao, Hong Zhai, Shaozhen He, Ning Zhao, and Qingchang Liu. 2023. "Genome-Wide Identification and Expression Analysis of the Sucrose Synthase Gene Family in Sweet Potato and Its Two Diploid Relatives" International Journal of Molecular Sciences 24, no. 15: 12493. https://doi.org/10.3390/ijms241512493