

Isoalantolactone Suppresses Glycolysis and Resensitizes Cisplatin-Based Chemotherapy in Cisplatin-Resistant Ovarian Cancer Cells

Abstract
:1. Introduction
2. Results
2.1. Isoalantolactone Markedly Inhibits the Growth of A2780cisR and SNU-8cisR OC Cells
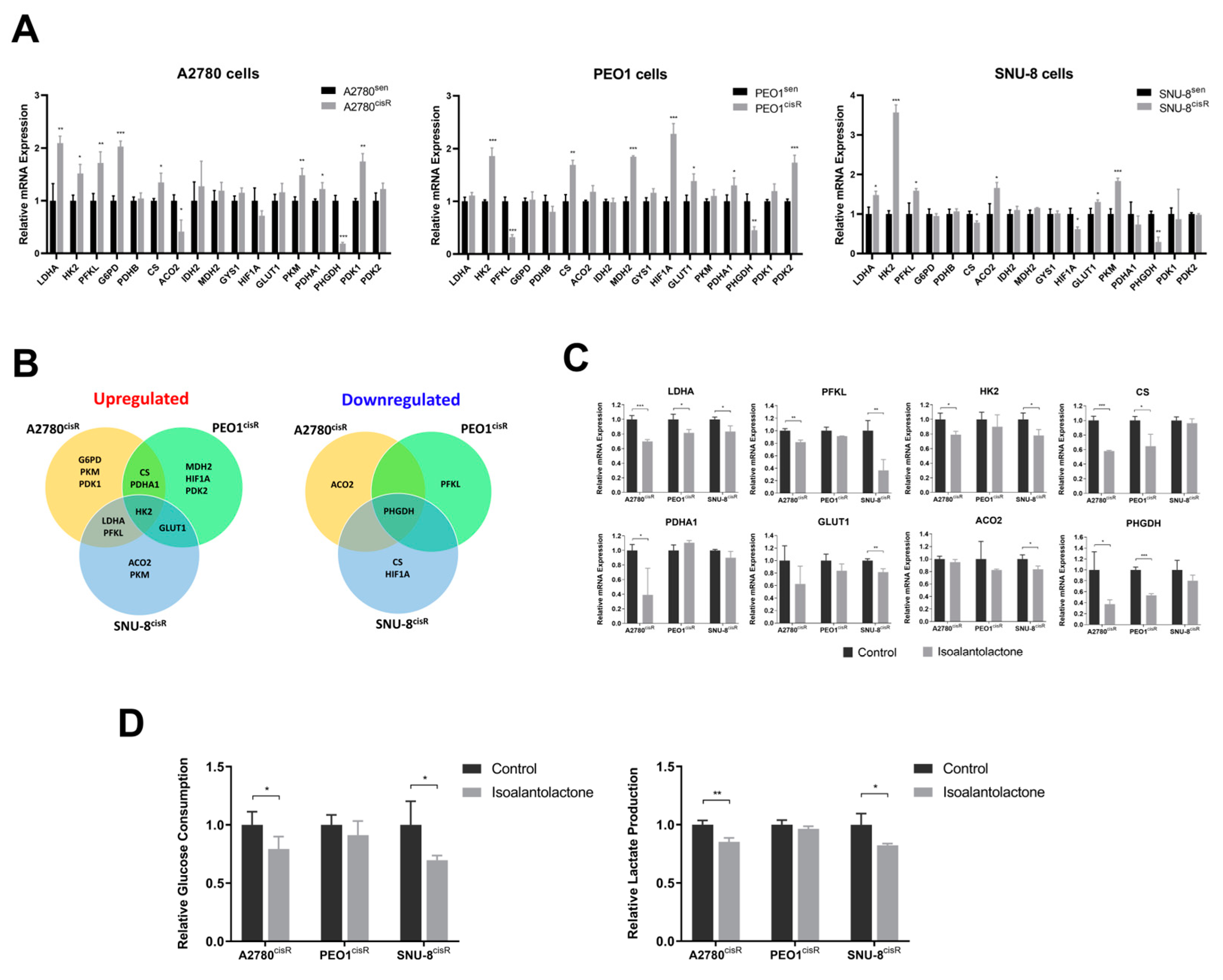
2.2. Isoalantolactone Inhibits Elevated Glycolysis in A2780cisR and SNU-8cisR OC Cells
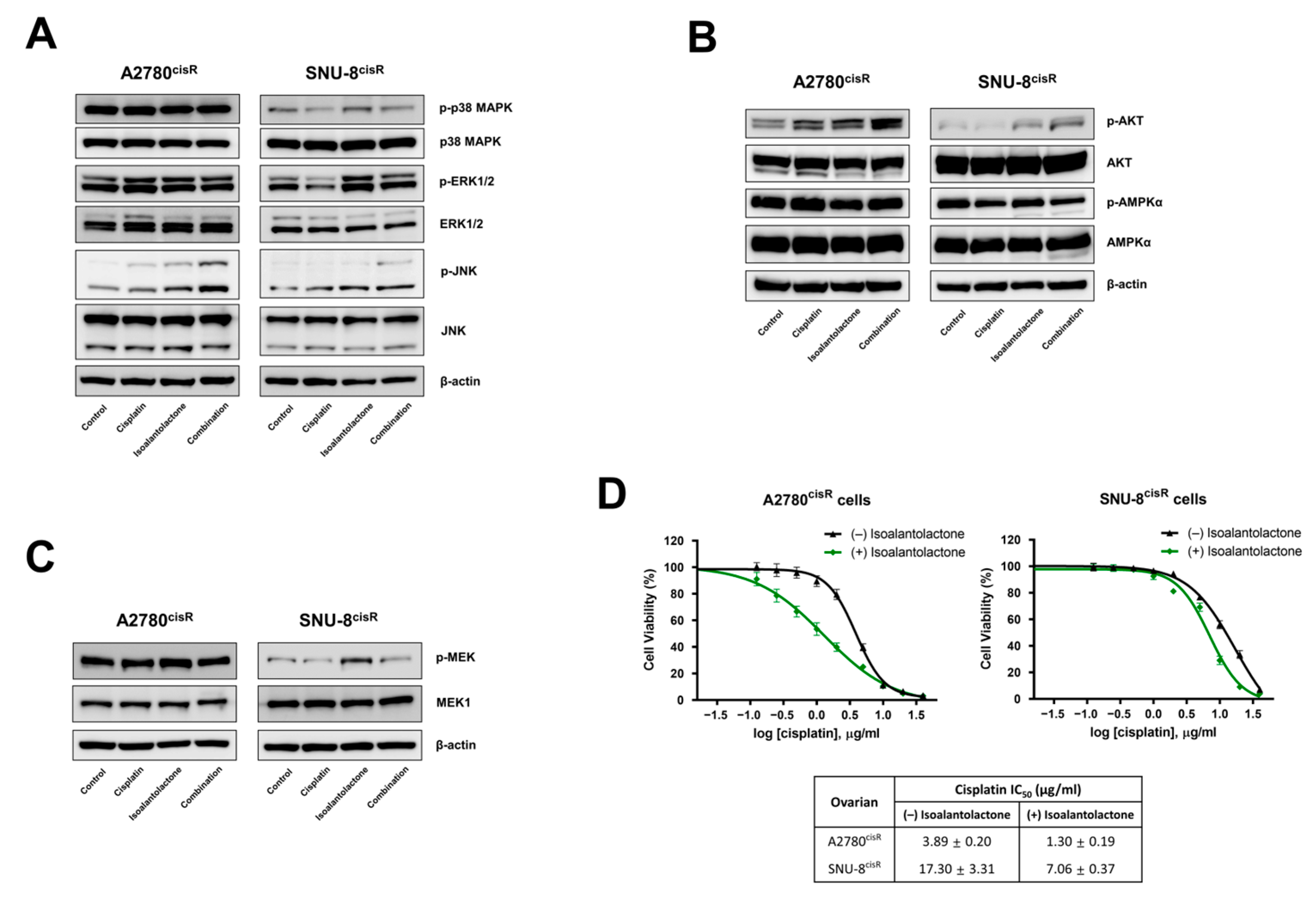
2.3. Isoalantolactone Increases the Sensitivity of A2780cisR and SNU-8cisR OC Cells to Cisplatin via the Apoptotic Pathway
2.4. Isoalantolactone Potentiates Cisplatin-Induced Apoptosis by Regulating the Survival Signaling Pathway
2.5. Isoalantolactone–Cisplatin Treatment Inhibits A2780cisR Xenograft Tumor Growth
2.6. Network Pharmacology Analysis Predicts the Target Pathways of Isoalantolactone-Regulated Glycolysis for OC Treatment
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Apoptosis Analysis
4.5. Quantitative RT-PCR
4.6. Metabolic Assays
4.7. Western Blotting
4.8. Measurement of Caspase-3/7 Activity
4.9. Tumor-Bearing Mice and Treatment
4.10. Systematic Pharmacological Analysis of Isoalantolactone
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Torre, L.A.; Islami, F.; Siegel, R.L.; Ward, E.M.; Jemal, A. Global cancer in women: Burden and trends. Cancer Epidemiol. Biomark. Prev. 2017, 26, 444–457. [Google Scholar] [CrossRef] [Green Version]
- Chandra, A.; Pius, C.; Nabeel, M.; Nair, M.; Vishwanatha, J.K.; Ahmad, S.; Basha, R. Ovarian cancer: Current status and strategies for improving therapeutic outcomes. Cancer Med. 2019, 8, 7018–7031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guarneri, V.; Piacentini, F.; Barbieri, E.; Conte, P.F. Achievements and unmet needs in the management of advanced ovarian cancer. Gynecol. Oncol. 2010, 117, 152–158. [Google Scholar] [CrossRef]
- Brown, A.; Kumar, S.; Tchounwou, P.B. Cisplatin-based chemotherapy of human cancers. J. Cancer Sci. Ther. 2019, 11, 97. [Google Scholar] [PubMed]
- Song, M.; Cui, M.; Liu, K. Therapeutic strategies to overcome cisplatin resistance in ovarian cancer. Eur. J. Med. Chem. 2022, 232, 114205. [Google Scholar] [CrossRef]
- Koberle, B.; Tomicic, M.T.; Usanova, S.; Kaina, B. Cisplatin resistance: Preclinical findings and clinical implications. Biochim. Biophys. Acta 2010, 1806, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Rebucci, M.; Michiels, C. Molecular aspects of cancer cell resistance to chemotherapy. Biochem. Pharmacol. 2013, 85, 1219–1226. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, X.; Fu, J.; Xu, W.; Yuan, J. The role of tumour metabolism in cisplatin resistance. Front. Mol. Biosci. 2021, 8, 691795. [Google Scholar] [CrossRef]
- Hay, N. Reprogramming glucose metabolism in cancer: Can it be exploited for cancer therapy? Nat. Rev. Cancer 2016, 16, 635–649. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Suo, C.; Li, S.T.; Zhang, H.; Gao, P. Metabolic reprogramming for cancer cells and their microenvironment: Beyond the Warburg effect. Biochim. Biophys. Acta Rev. Cancer 2018, 1870, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Gao, W.; Zhang, Y.; Wu, S.; Liu, Y.; Deng, X.; Xie, L.; Yang, J.; Yu, H.; Su, J.; et al. ABT737 reverses cisplatin resistance by targeting glucose metabolism of human ovarian cancer cells. Int. J. Oncol. 2018, 53, 1055–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varghese, E.; Samuel, S.M.; Liskova, A.; Samec, M.; Kubatka, P.; Busselberg, D. Targeting glucose metabolism to overcome resistance to anticancer chemotherapy in breast cancer. Cancers 2020, 12, 2252. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Mahalanobish, S.; Saha, S.; Ghosh, S.; Sil, P.C. Natural products: An upcoming therapeutic approach to cancer. Food Chem. Toxicol. 2019, 128, 240–255. [Google Scholar] [CrossRef]
- Zhang, X.; Qiu, H.; Li, C.; Cai, P.; Qi, F. The positive role of traditional Chinese medicine as an adjunctive therapy for cancer. Bio Sci. Trends 2021, 15, 283–298. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.R.; Chang, C.H.; Hsu, C.F.; Tsai, M.J.; Cheng, H.; Leong, M.K.; Sung, P.J.; Chen, J.C.; Weng, C.F. Natural compounds as potential adjuvants to cancer therapy: Preclinical evidence. Br. J. Pharmacol. 2020, 177, 1409–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Tan, H.Y.; Chen, F.; Wang, N.; Feng, Y. Targeting cancer metabolism to resensitize chemotherapy: Potential development of cancer chemosensitizers from traditional Chinese medicines. Cancers 2020, 12, 404. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.H.; Song, Y.D.; Wu, Y.X.; He, H.Q.; Yu, T.H.; Hu, Y.D.; Zhang, D.P.; Jiang, H.C.; Yu, K.K.; Li, X.Z.; et al. Isoalantolactone suppresses LPS-induced inflammation by inhibiting TRAF6 ubiquitination and alleviates acute lung injury. Acta Pharmacol. Sin. 2019, 40, 64–74. [Google Scholar] [CrossRef] [Green Version]
- He, G.; Zhang, X.; Chen, Y.; Chen, J.; Li, L.; Xie, Y. Isoalantolactone inhibits LPS-induced inflammation via NF-κB inactivation in peritoneal macrophages and improves survival in sepsis. Biomed. Pharmacother. 2017, 90, 598–607. [Google Scholar] [CrossRef]
- Wu, M.; Zhang, H.; Hu, J.; Weng, Z.; Li, C.; Li, H.; Zhao, Y.; Mei, X.; Ren, F.; Li, L. Isoalantolactone inhibits UM-SCC-10A cell growth via cell cycle arrest and apoptosis induction. PLoS ONE 2013, 8, e76000. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Zhang, G.; Zhang, Y.; Hua, P.; Song, G.; Sun, M.; Li, X.; Tong, T.; Li, B.; Zhang, X. Isoalantolactone induces intrinsic apoptosis through p53 signaling pathway in human lung squamous carcinoma cells. PLoS ONE 2017, 12, e0181731. [Google Scholar] [CrossRef] [Green Version]
- Schultze, S.M.; Hemmings, B.A.; Niessen, M.; Tschopp, O. PI3K/AKT, MAPK and AMPK signalling: Protein kinases in glucose homeostasis. Expert Rev. Mol. Med. 2012, 14, e1. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.R.; Chua, K.N.; Sim, W.J.; Ng, H.C.; Bi, C.; Ho, J.; Nga, M.E.; Pang, Y.H.; Ong, W.R.; Soo, R.A.; et al. MEK inhibition overcomes cisplatin resistance conferred by SOS/MAPK pathway activation in squamous cell carcinoma. Mol. Cancer Ther. 2015, 14, 1750–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao da, C.; Xiao, P.G. Network pharmacology: A Rosetta Stone for traditional Chinese medicine. Drug Dev. Res. 2014, 75, 299–312. [Google Scholar] [CrossRef]
- Cocetta, V.; Ragazzi, E.; Montopoli, M. Links between cancer metabolism and cisplatin resistance. Int. Rev. Cell Mol. Biol. 2020, 354, 107–164. [Google Scholar] [PubMed]
- Catanzaro, D.; Gaude, E.; Orso, G.; Giordano, C.; Guzzo, G.; Rasola, A.; Ragazzi, E.; Caparrotta, L.; Frezza, C.; Montopoli, M. Inhibition of glucose-6-phosphate dehydrogenase sensitizes cisplatin-resistant cells to death. Oncotarget 2015, 6, 30102–30114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, M.; Chen, X.S.; Li, L.Y.; Wu, H.Z.; Zeng, D.; Wang, X.L.; Zhang, Y.; Xiao, S.S.; Cheng, Y. Inhibition of AXL enhances chemosensitivity of human ovarian cancer cells to cisplatin via decreasing glycolysis. Acta Pharmacol. Sin. 2021, 42, 1180–1189. [Google Scholar] [CrossRef]
- Tan, V.P.; Miyamoto, S. HK2/hexokinase-II integrates glycolysis and autophagy to confer cellular protection. Autophagy 2015, 11, 963–964. [Google Scholar] [CrossRef] [Green Version]
- Suh, D.H.; Kim, M.A.; Kim, H.; Kim, M.K.; Kim, H.S.; Chung, H.H.; Kim, Y.B.; Song, Y.S. Association of overexpression of hexokinase II with chemoresistance in epithelial ovarian cancer. Clin. Exp. Med. 2014, 14, 345–353. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Zhang, M.; Cong, Q.; Zhang, M.X.; Zhang, M.Y.; Lu, Y.Y.; Xu, C.J. Hexokinase 2 confers resistance to cisplatin in ovarian cancer cells by enhancing cisplatin-induced autophagy. Int. J. Biochem. Cell Biol. 2018, 95, 9–16. [Google Scholar] [CrossRef]
- Li, G.; Li, Y.; Wang, D.Y. Overexpression of miR-329-3p sensitizes osteosarcoma cells to cisplatin through suppression of glucose metabolism by targeting LDHA. Cell Biol. Int. 2021, 45, 766–774. [Google Scholar] [CrossRef]
- Zheng, C.; Yu, X.; Liang, Y.; Zhu, Y.; He, Y.; Liao, L.; Wang, D.; Yang, Y.; Yin, X.; Li, A.; et al. Targeting PFKL with penfluridol inhibits glycolysis and suppresses esophageal cancer tumorigenesis in an AMPK/FOXO3a/BIM-dependent manner. Acta Pharm. Sin. B 2022, 12, 1271–1287. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.Y.; Zhang, Q.; Zhang, B.; Yang, B.; Lin, N.M. Active ingredients of Inula helenium L. exhibits similar anti-cancer effects as isoalantolactone in pancreatic cancer cells. Nat. Prod. Res. 2020, 34, 2539–2544. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Yang, P. Isoalantolactone exerts anticancer effects on human HEC-1-B endometrial cancer cells via induction of ROS mediated apoptosis and inhibition of MEK/ERK signalling pathway. Acta Biochim. Pol. 2022, 69, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Shao, R.; Zheng, P.; Zhang, T.; Qiu, C.; Sui, H.; Li, S.; Jin, L.; Pan, H.; Jin, X.; et al. Isoalantolactone enhances the antitumor activity of doxorubicin by inducing reactive oxygen species and DNA damage. Front. Oncol. 2022, 12, 813854. [Google Scholar] [CrossRef]
- Kim, M.Y.; Lee, H.; Ji, S.Y.; Kim, S.Y.; Hwangbo, H.; Park, S.H.; Kim, G.Y.; Park, C.; Leem, S.H.; Hong, S.H.; et al. Induction of apoptosis by isoalantolactone in human hepatocellular carcinoma Hep3B cells through activation of the ROS-dependent JNK signaling pathway. Pharmaceutics 2021, 13, 1627. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.S.; Chen, C.; Liu, Z.; Liu, S.L.; Guo, J.H.; Zhang, C.; Zhang, Q.W.; Gao, F.H. Isoalantolactone mediates the degradation of BCR-ABL protein in imatinib-resistant CML cells by down-regulating survivin. Cell Cycle 2023, 22, 1407–1420. [Google Scholar] [CrossRef]
- Parker, R.J.; Eastman, A.; Bostick-Bruton, F.; Reed, E. Acquired cisplatin resistance in human ovarian cancer cells is associated with enhanced repair of cisplatin-DNA lesions and reduced drug accumulation. J. Clin. Investig. 1991, 87, 772–777. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Feng, Q.; Kim, J.M.; Schneiderman, D.; Liston, P.; Li, M.; Vanderhyden, B.; Faught, W.; Fung, M.F.; Senterman, M.; et al. Human ovarian cancer and cisplatin resistance: Possible role of inhibitor of apoptosis proteins. Endocrinology 2001, 142, 370–380. [Google Scholar] [CrossRef]
- Li, F.L.; Liu, J.P.; Bao, R.X.; Yan, G.; Feng, X.; Xu, Y.P.; Sun, Y.P.; Yan, W.; Ling, Z.Q.; Xiong, Y.; et al. Acetylation accumulates PFKFB3 in cytoplasm to promote glycolysis and protects cells from cisplatin-induced apoptosis. Nat. Commun. 2018, 9, 508. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Zhang, Y.; Chen, T.; Wang, Y.; Xue, J.; Zhang, Y.; Xiao, W.; Mo, X.; Lu, Y. Efficacy of RNAi targeting of pyruvate kinase M2 combined with cisplatin in a lung cancer model. J. Cancer Res. Clin. Oncol. 2011, 137, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Loar, P.; Wahl, H.; Kshirsagar, M.; Gossner, G.; Griffith, K.; Liu, J.R. Inhibition of glycolysis enhances cisplatin-induced apoptosis in ovarian cancer cells. Am. J. Obstet. Gynecol. 2010, 202, 371.e1–371.e8. [Google Scholar] [CrossRef] [PubMed]
- Achkar, I.W.; Abdulrahman, N.; Al-Sulaiti, H.; Joseph, J.M.; Uddin, S.; Mraiche, F. Cisplatin based therapy: The role of the mitogen activated protein kinase signaling pathway. J. Transl. Med. 2018, 16, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, D.J.; Wang, J.; Zhou, J.Y.; Wu, G.S. Role of the Akt/mTOR survival pathway in cisplatin resistance in ovarian cancer cells. Biochem. Biophys. Res. Commun. 2010, 394, 600–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brozovic, A.; Osmak, M. Activation of mitogen-activated protein kinases by cisplatin and their role in cisplatin-resistance. Cancer Lett. 2007, 251, 1–16. [Google Scholar] [CrossRef]
- Banerjee, S.; Kaye, S.B. New strategies in the treatment of ovarian cancer: Current clinical perspectives and future potential. Clin. Cancer Res. 2013, 19, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Moserle, L.; Jimenez-Valerio, G.; Casanovas, O. Antiangiogenic therapies: Going beyond their limits. Cancer Discov. 2014, 4, 31–41. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | Forward Sequences | Reverse Sequences |
|---|---|---|
| LDHA | GGA TCT CCA ACA TGG CAG CCT T | AGA CGG CTT TCT CCC TCT TGC T |
| HK2 | GAG TTT GAC CTG GAT GTG GTT GC | CCT CCA TGT AGC AGG CAT TGC T |
| PFKL | AAG AAG TAG GCT GGC ACG ACG T | GCG GAT GTT CTC CAC AAT GGA C |
| G6PD | CTG TTC CGT GAG GAC CAG ATC T | TGA AGG TGA GGA TAA CGC AGG C |
| PDHB | TGT AAC TGT GGA AGG AGG CTG G | CAT CAG CAC CAG TGA CAC GAA C |
| CS | CAC AGG GTA TCA GCC GAA CCA A | CCA ATA CCG CTG CCT TCT CTG T |
| ACO2 | CAA TCG TCA CCT CCT ACA ACA GG | GTC TCT GGG TTG AAC TTG AGG G |
| IDH2 | AGA TGG CAG TGG TGT CAA GGA G | CTG GAT GGC ATA CTG GAA GCA G |
| MDH2 | CTG GAC ATC GTC AGA GCC AAC A | GGA TGA TGG TCT TCC CAG CAT G |
| GYS1 | CCG CTA TGA GTT CTC CAA CAA GG | AGA AGG CAA CCA CTG TCT GCT C |
| HIF1A | TAT GAG CCA GAA GAA CTT TTA GGC | CAC CTC TTT TGG CAA GCA TCC TG |
| GLUT1 | TTG CAG GCT TCT CCA ACT GGA C | CAG AAC CAG GAG CAC AGT GAA G |
| PKM | ATG GCT GAC ACA TTC CTG GAG C | CCT TCA ACG TCT CCA CTG ATC G |
| PDHA1 | GGA TGG TGA ACA GCA ATC TTG CC | TCG CTG GAG TAG ATG TGG TAG C |
| PHGDH | CTT ACC AGT GCC TTC TCT CCA C | GCT TAG GCA GTT CCC AGC ATT C |
| PDK1 | CAT GTC ACG CTG GGT AAT GAG G | CTC AAC ACG AGG TCT TGG TGC A |
| PDK2 | TGC CTA CGA CAT GGC TAA GCT C | GAC GTA GAC CAT GTG AAT CGG C |
| ACTB | CAC CAT TGG CAA TGA GCG GTT C | AGG TCT TTG CGG ATG TCC ACG T |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chun, J. Isoalantolactone Suppresses Glycolysis and Resensitizes Cisplatin-Based Chemotherapy in Cisplatin-Resistant Ovarian Cancer Cells. Int. J. Mol. Sci. 2023, 24, 12397. https://doi.org/10.3390/ijms241512397
Chun J. Isoalantolactone Suppresses Glycolysis and Resensitizes Cisplatin-Based Chemotherapy in Cisplatin-Resistant Ovarian Cancer Cells. International Journal of Molecular Sciences. 2023; 24(15):12397. https://doi.org/10.3390/ijms241512397
Chicago/Turabian StyleChun, Jaemoo. 2023. "Isoalantolactone Suppresses Glycolysis and Resensitizes Cisplatin-Based Chemotherapy in Cisplatin-Resistant Ovarian Cancer Cells" International Journal of Molecular Sciences 24, no. 15: 12397. https://doi.org/10.3390/ijms241512397