
DNA-Binding One Finger Transcription Factor PhDof28 Regulates Petal Size in Petunia
 , and
, and 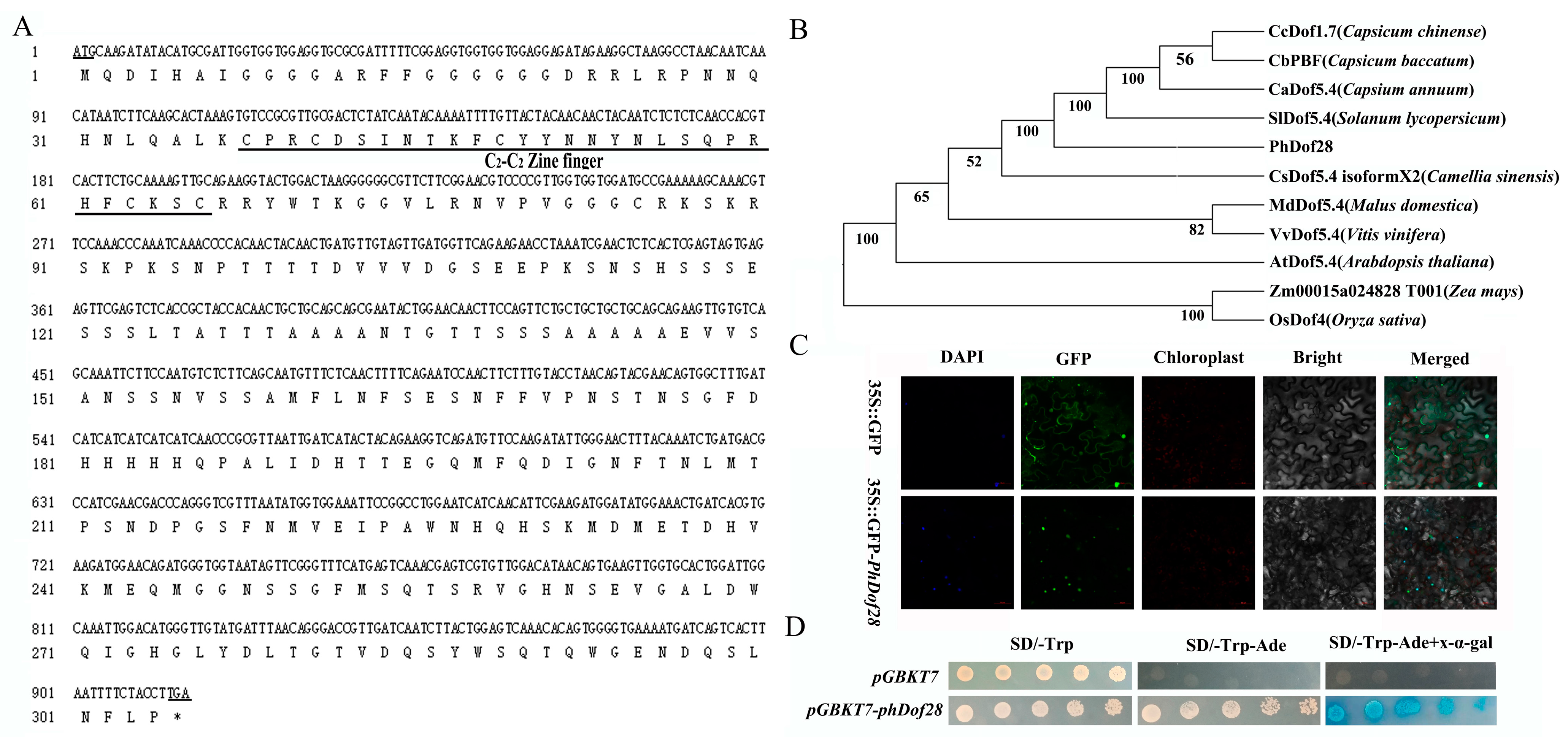{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
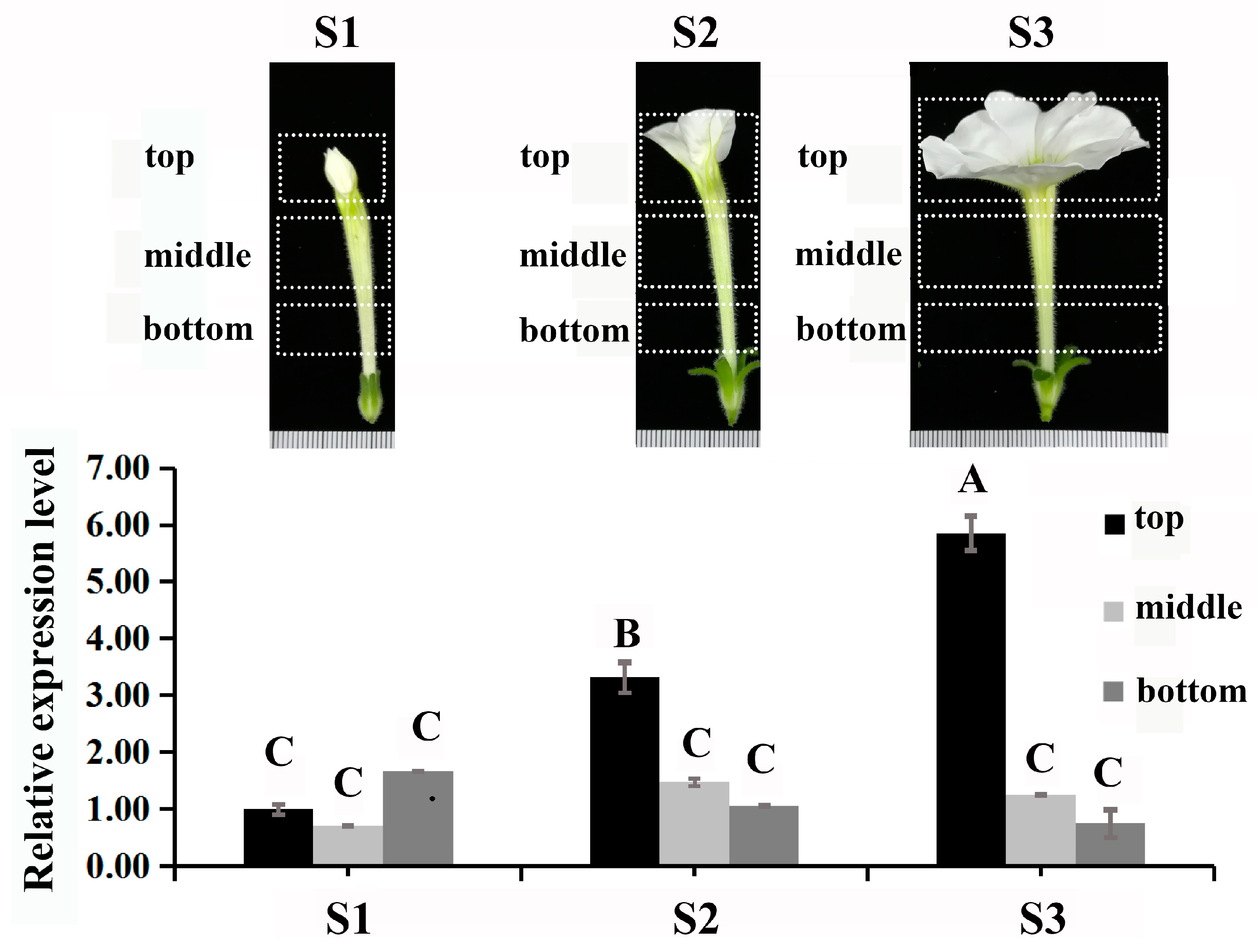
2.1. Sequence Features, Subcellular Localization, Self-Activation, and Expression of PhDof28
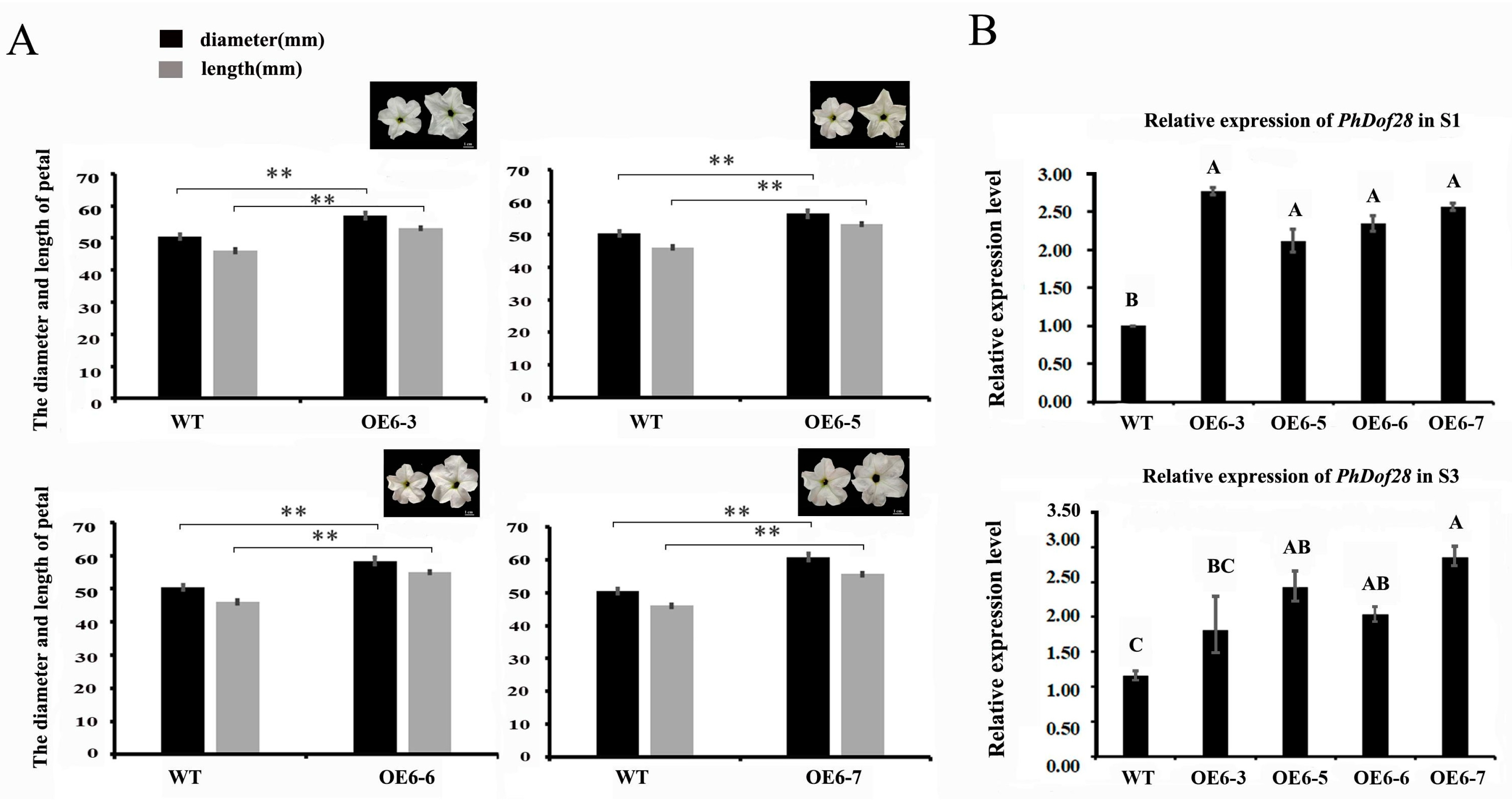
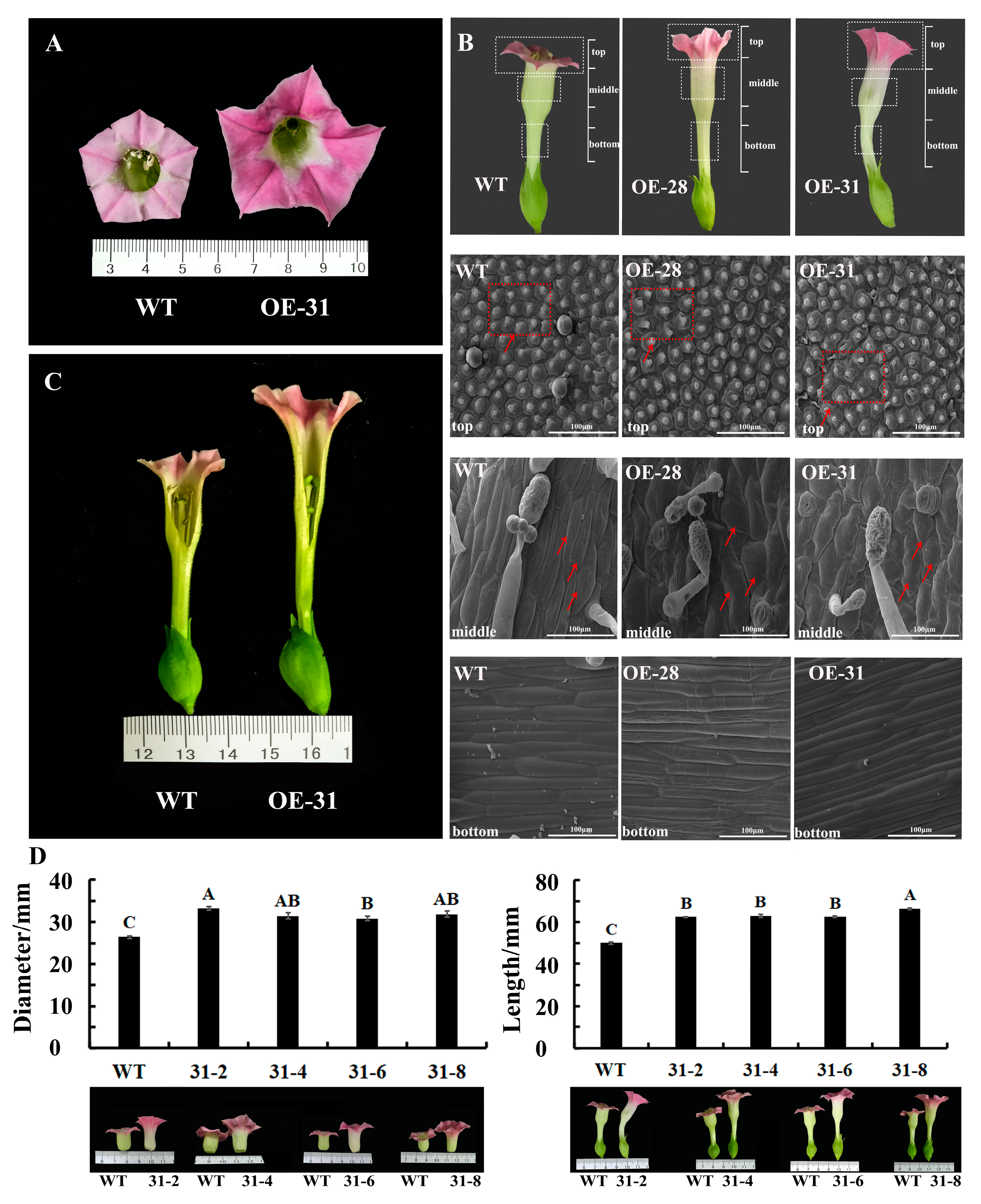
2.2. Phenotypic Characteristics of Transgenic Petunia Plants
2.3. Phenotypic Characteristics of Transgenic Tobacco Plants
2.4. The Endogenous Hormone Contents in Transgenic Petunia
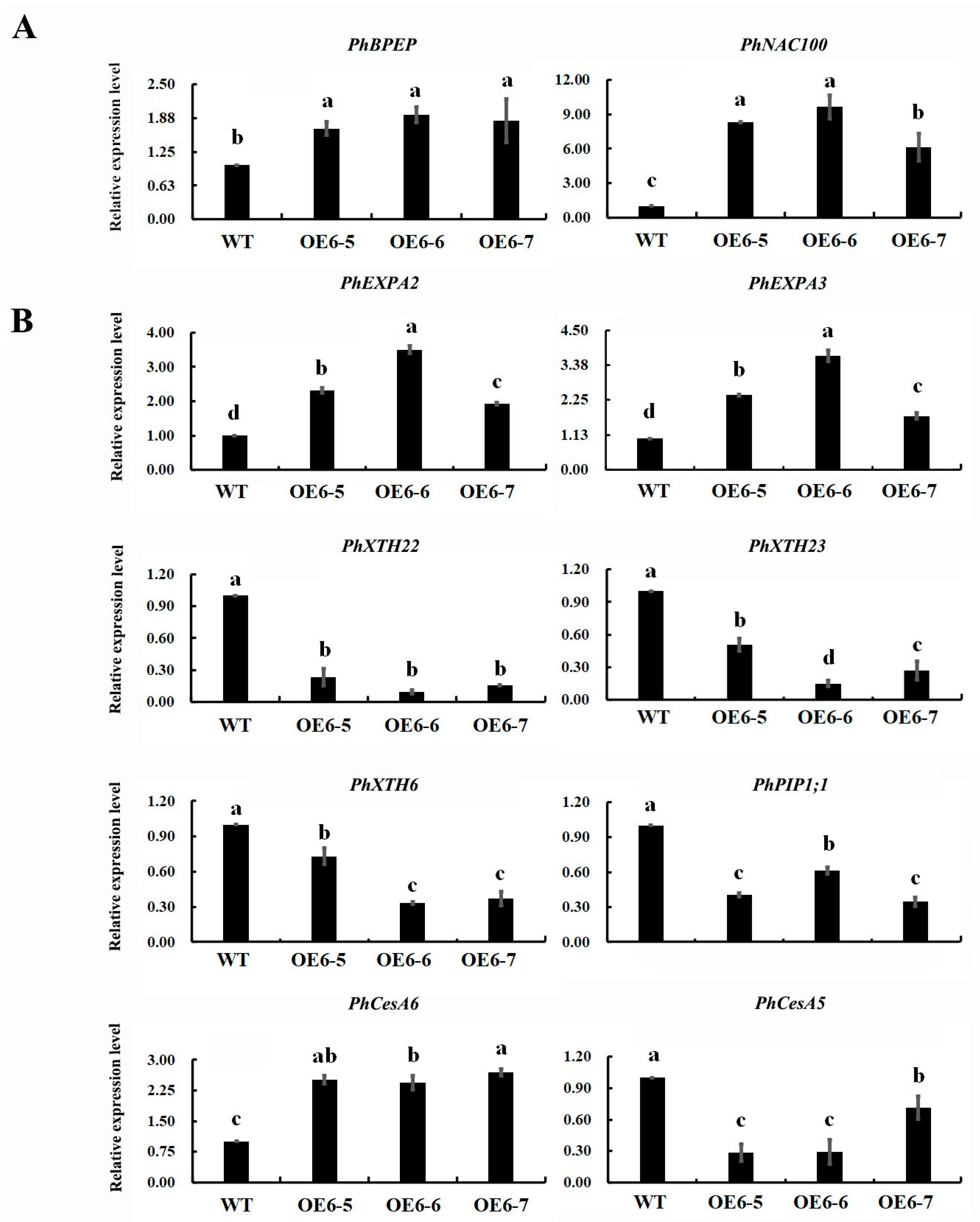
2.5. Expression Profile of Cell Expansion Associated Genes in Transgenic Petunia Petals
3. Discussion
3.1. Overexpression of PhDof28 Causes Petal Expansion in Petunia and Tobacco
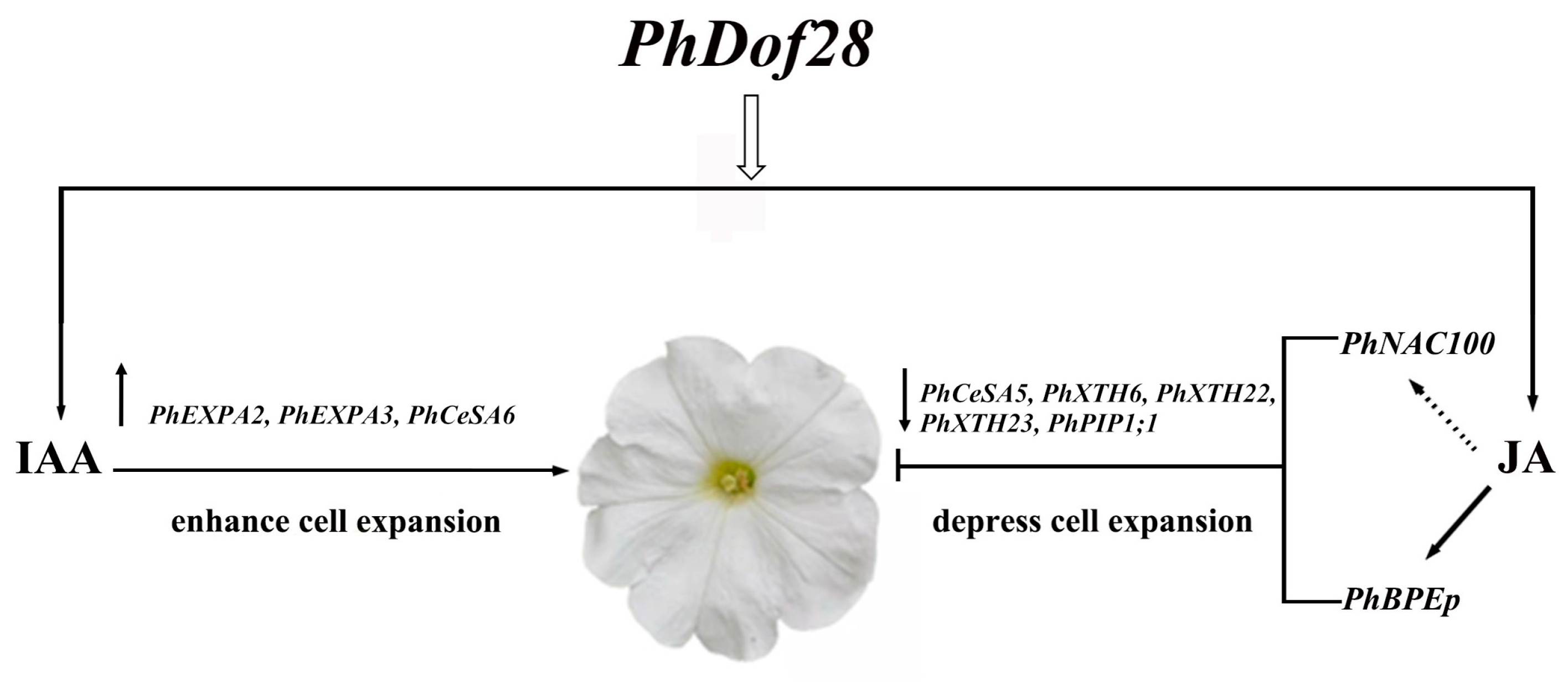
3.2. PhDof28 Regulates Petal Size by Triggering IAA and JA-Related Pathways
4. Materials and Methods
4.1. Plant Materials
4.2. qRT-PCR Analysis
4.3. Subcellular Localization and Transcriptional Self-Activation Assay
4.4. Stable Transformation of Petunia and Tobacco Plants
4.5. Scanning Electron Microscope (SEM) Analysis of Petal Cells
4.6. Measurement of Hormone Content
4.7. Screening for Petal Candidate Genes Associated with Cell Expansion
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baumann, K.; Perez-Rodriguez, M.; Bradley, D.; Venail, C.; Bailey, P.; Jin, H.; Koes, R.; Roberts, K.; Martin, C. Control of Cell and Petal Morphogenesis by R2R3 MYB Transcription Factors. Development 2007, 134, 1691–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meagher, T.R.; Gillies, A.C.M.; Costich, D.E. Genome Size, Quantitative Genetics and The Genomic Basis for Flower Size Evolution in Silene latifolia. Ann. Bot. 2005, 95, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Pei, H.X.; Ma, N.; Tian, J.; Luo, J.; Chen, J.W.; Li, J.; Zheng, Y.; Chen, X.; Fei, Z.; Gao, J.P. An NAC Transcription Factor Controls Ethylene-regulated Cell Expansion in Flower Petals. Plant Physiol. 2013, 163, 775–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepworth, J.; Lenhard, M. Regulation of Plant Lateral-organ Growth by Modulating Cell Number and Size. Curr. Opin. Plant Biol. 2014, 17, 36–42. [Google Scholar] [CrossRef]
- Huang, T.B.; Irish, V.F. Gene Networks Controlling Petal Organogenesis. J. Exp. Bot. 2016, 67, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.X.; Hussain, N.; Wang, Y.R.; Li, M.T.; Liu, L.; Qin, M.Z.; Ma, N.; Gao, J.P.; Sun, X.M. An Ethylene-inhibited NF-YC Transcription Factor RhNF-YC9 Regulates Petal Expansion in Rose. Hortic. Plant J. 2020, 6, 419–427. [Google Scholar] [CrossRef]
- Varaud, E.; Brioudes, F.; Szécsi, J.; Leroux, J.; Brown, S.; Rechenmann, C.P.; Bendahmane, M. AUXIN RESPONSE FACTOR8 Regulates Arabidopsis Petal Growth by Interacting with the bHLH Transcription Factor BIGPETALp. Plant Cell 2011, 23, 973–983. [Google Scholar] [CrossRef] [Green Version]
- Brioudes, F.; Joly, C.; Szécsi, J.; Varaud, E.; Leroux, J.; Bellvert, F.; Bertrand, C.; Bendahmane, M. Jasmonate Controls Late Development Stages of Petal Growth in Arabidopsis thaliana. Plant J. 2009, 60, 1070–1080. [Google Scholar] [CrossRef]
- Luo, J.; Ma, N.; Pei, H.X.; Chen, J.W.; Li, J.; Gao, J.P. A DELLA Gene, RhGAI1, is a Direct Target of EIN3 and Mediates Ethylene-regulated Rose Petal Cell Expansion via Repressing the Expression of RhCesA2. J. Exp. Bot. 2013, 64, 5075–5084. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Guan, Y.X.; Ding, L.; Li, P.R.; Zhao, W.Q.; Jiang, J.F.; Chen, S.M.; Chen, F.D. The CmTCP20 Gene Regulates Petal Elongation Growth in Chrysanthemum morifolium. Plant Sci. 2019, 280, 248–257. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, N.N.; Deng, X.; Liu, D.M.; Li, M.F.; Cui, D.D.; Hu, Y.K.; Yan, Y.M. Genome-wide Analysis of Wheat DNA-binding with One Finger (Dof) Transcription Factor Genes: Evolutionary Characteristics and Diverse Abiotic Stress Responses. BMC Genom. 2020, 21, 276–294. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Chen, Q.J.; Zhu, Y.D.; Li, T.Z. Identification of MdDof Genes in Apple and Analysis of Their Response to Biotic or Abiotic Stress. Funct. Plant Biol. 2018, 45, 528–541. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.C.; Hou, D.; Liu, J.; Li, X.Y.; Xie, L.H.; Ma, Y.J.; Gao, J. Characterization of Moso Bamboo (Phyllostachys edulis) Dof Transcription Factors in Floral Development and Abiotic Stress Responses. Genome 2018, 61, 151–156. [Google Scholar] [CrossRef]
- Ahmad, M.; Rim, Y.; Chen, H.; Kim, J.Y. Functional Characterization of Arabidopsis Dof Transcription Factor AtDof4.1. Russ. J. Plant Physiol. 2013, 60, 116–123. [Google Scholar] [CrossRef]
- Xu, P.P.; Chen, H.Y.; Ying, L.; Cai, W.M. AtDOF5.4/OBP4, a DOF Transcription Factor Gene that Negatively Regulates Cell Cycle Progression and Cell Expansion in Arabidopsis thaliana. Sci. Rep. 2016, 6, 27705–27718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, Y.Z.; Du, J.H.; Li, Y.; Thomas, H.R.; Frank, M.H.; Wang, L.G.; Hu, H.R. Insight into The Petunia Dof Transcription Factor Family Reveals a New Regulator of Male-sterility. Ind. Crops Prod. 2021, 161, 113196. [Google Scholar] [CrossRef]
- Norikoshi, R.; Shibata, T.; Ichimura, K. Cell Division and Expansion in Petals During Flower Development and Opening in Eustoma grandiflorum. Hortic. J. 2016, 85, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.F.; Chen, C.B.; Chen, J.Y.; Chen, J.X.; Hua, Q.Z.; Shah, K.; Zhang, Z.K.; Zhao, J.T.; Hu, G.B.; Chen, J.Y.; et al. Betalain Biosynthesis in Red Pulp Pitaya is Regulated via HuMYB132: A R-R Type MYB Transcription Factor. BMC Plant Biol. 2023, 23, 28–42. [Google Scholar] [CrossRef]
- Sagawa, J.M.; Stanley, L.E.; Lafountain, A.M.; Frank, H.A.; Liu, C.; Yuan, Y.W. An R2R3-MYB Transcription Factor Regulates Carotenoid Pigmentation in Mimulus lewisii Flowers. New Phytol. 2016, 209, 1049–1057. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Shi, Y.Q.; Sun, M.D.; Hu, X.Z.; Hao, M.Y.; Shu, Y.; Zhou, X.R.; Hu, Q.; Li, C.; Mei, D.S. Functional Differentiation of BnVTE4 Gene Homologous Copies in α-tocopherol Biosynthesis Revealed by CRISPR/Cas9 Editing. Front. Plant Sci. 2022, 13, 850914–850923. [Google Scholar] [CrossRef]
- Zhai, K.H.; Zhao, G.W.; Jiang, H.Y.; Sun, C.X.; Ren, J.Y. Overexpression of Maize ZmMYB59 Gene Plays a Negative Regulatory Role in Seed Germination in Nicotiana tabacum and Oryza sativa. Front. Plant Sci. 2020, 11, 564665–564674. [Google Scholar] [CrossRef]
- Wu, R.; Ding, Y.Q.; Li, C.Y.; Wu, B.K.; Huang, Z.J.; Li, Z.N.; Wang, X.M.; Zhao, G.W. An R2R3-type Transcription Factor OsMYBAS1 Regulates Seed Germination Under Artificial Accelerated Aging in Transgenic Rice (Oryza sativa L.). Agronomy 2022, 12, 1955. [Google Scholar] [CrossRef]
- Su, Y.H.; Liu, Y.B.; Bai, B.; Zhang, X.S. Establishment of Embryonic Shoot-root Axis is Involved in Auxin and Cytokinin Response during Arabidopsis Somatic Embryogenesis. Front. Plant Sci. 2015, 5, 781–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majda, M.; Robert, S. The Role of Auxin in Cell Wall Expansion. Int. J. Mol. Sci. 2018, 19, 951–972. [Google Scholar] [CrossRef] [Green Version]
- Du, M.M.; Edgar, P.S.; William, M.G. Rapid Auxin-mediated Cell Expansion. Annu Rev. Plant Biol. 2020, 71, 379–402. [Google Scholar] [CrossRef]
- Rose, J.K.C.; Braam, J.; Fry, S.C.; Nishitani, K. The XTH Family of Enzymes Involved in Xyloglucan Endotransglucosylation and Endohydrolysis: Current Perspectives and A New Unifying Nomenclature. Plant Cell Physiol. 2002, 43, 1421–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutwil, M.; Debolt, S.; Persson, S. Cellulose Synthesis: A Complex Complex. Curr. Opin. Plant Biol. 2008, 11, 252–257. [Google Scholar] [CrossRef]
- Stintzi, A.; Browse, J. The Arabidopsis Male-sterile Mutant, opr3, Lacks the 12-oxophytodienoic Acid Reductase Required for Jasmonate Synthesis. Proc. Natl. Acad. Sci. USA 2000, 97, 10625–10630. [Google Scholar] [CrossRef]
- Szécsi, J.; Joly, C.; Bordji, K.; Varaud, E.; Cock, J.M.; Dumas, C.; Bendahmane, M. BIGPETALp, A bHLH Transcription Factor is Involved in The control of Arabidopsis Petal Size. EMBO J. 2006, 25, 3912–3920. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.K.; Mangalam, A.K.; Dwivedi, S.; Naik, S. Primer Premier: Program for Design of Degenerate Primers from A Protein Sequence. Biotechniques 1998, 24, 318–319. [Google Scholar] [CrossRef] [Green Version]
- Yue, Y.Z.; Yang, D.; Sun, J.; Peng, H.; Yin, C.Q.; Guo, R.; Ning, G.G.; Hu, H.R. A Novel PhLRR Gene Promoter is Sufficient for Engineering Male Sterility in Petunia. Plant Mol. Biol. Rep. 2016, 34, 940–977. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2020, 172, 103990. [Google Scholar] [CrossRef]
- Huang, X.; Yue, Y.Z.; Sun, J.; Peng, H.; Yang, Z.N.; Bao, M.Z.; Hu, H.R. Characterization of A Fertility-related SANT/MYB gene (PhRL) from Petunia. Sci. Hortic. 2015, 183, 152–159. [Google Scholar] [CrossRef]
- Gu, H.; Ding, W.J.; Shi, T.T.; Ouyang, Q.X.; Yang, X.L.; Yue, Y.Z.; Wang, L.G. Integrated Transcriptome and Endogenous Hormone Analysis Provides New Insights into Callus Proliferation in Osmanthus fragrans. Sci. Rep. 2022, 12, 7609–7622. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.L.; Ding, W.J.; Yue, Y.Z.; Xu, C.; Wang, X.; Wang, L.G. Cloning and Expression Analysis of Three Critical Triterpenoid Pathway Genes in Osmanthus fragrans. Electron. J. Biotechnol. 2018, 36, 1–8. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, Y.; Zhu, W.; Shen, H.; Wang, H.; Du, J.; Wang, L.; Hu, H. DNA-Binding One Finger Transcription Factor PhDof28 Regulates Petal Size in Petunia. Int. J. Mol. Sci. 2023, 24, 11999. https://doi.org/10.3390/ijms241511999
Yue Y, Zhu W, Shen H, Wang H, Du J, Wang L, Hu H. DNA-Binding One Finger Transcription Factor PhDof28 Regulates Petal Size in Petunia. International Journal of Molecular Sciences. 2023; 24(15):11999. https://doi.org/10.3390/ijms241511999
Chicago/Turabian StyleYue, Yuanzheng, Wuwei Zhu, Huimin Shen, Hongtao Wang, Juhua Du, Lianggui Wang, and Huirong Hu. 2023. "DNA-Binding One Finger Transcription Factor PhDof28 Regulates Petal Size in Petunia" International Journal of Molecular Sciences 24, no. 15: 11999. https://doi.org/10.3390/ijms241511999