
Daidzein Inhibits Human Platelet Activation by Downregulating Thromboxane A2 Production and Granule Release, Regardless of COX-1 Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introductions
2. Results
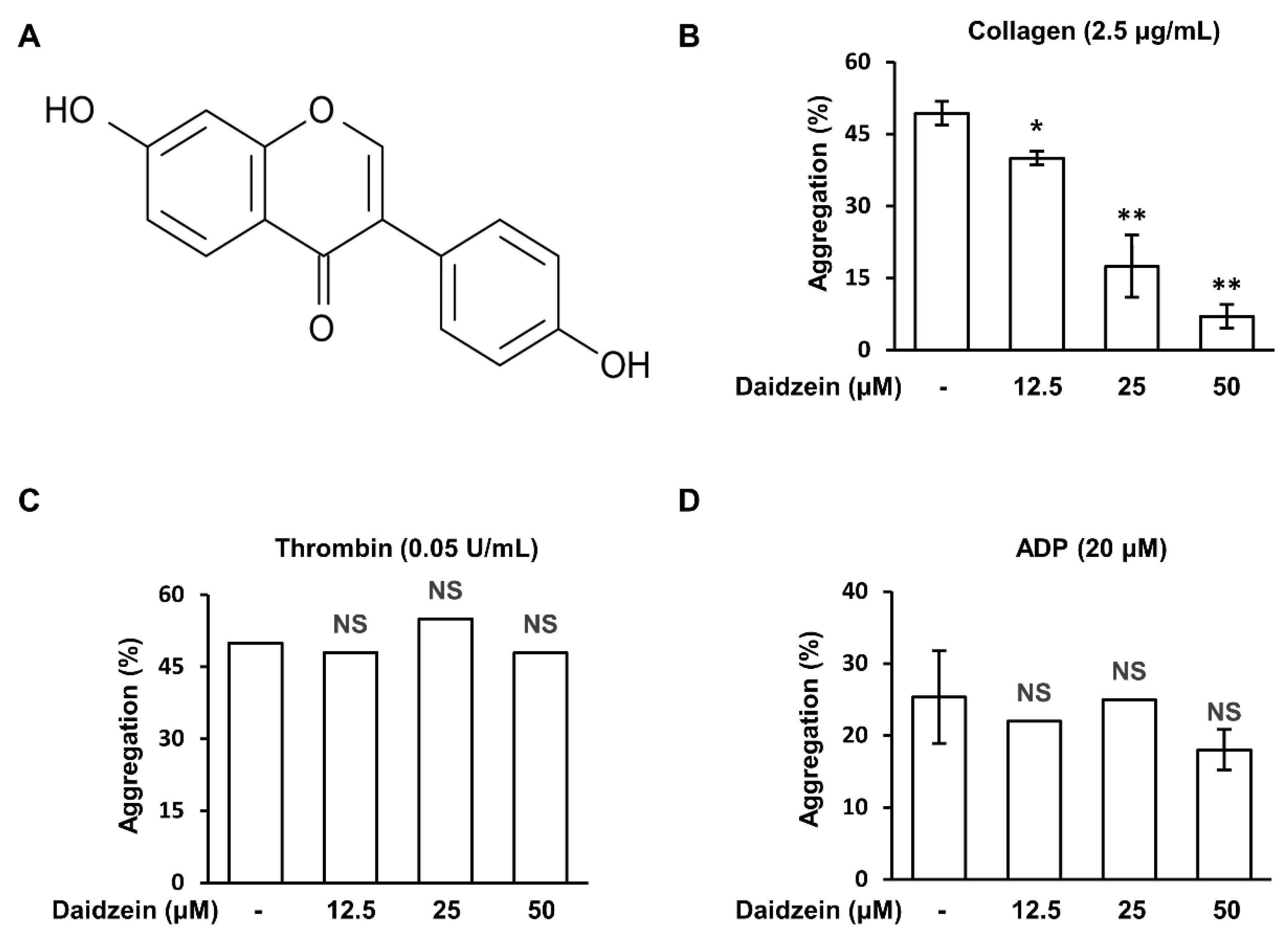
2.1. Effects of Daidzein on Human Platelet Aggregation induced by Different Agonists
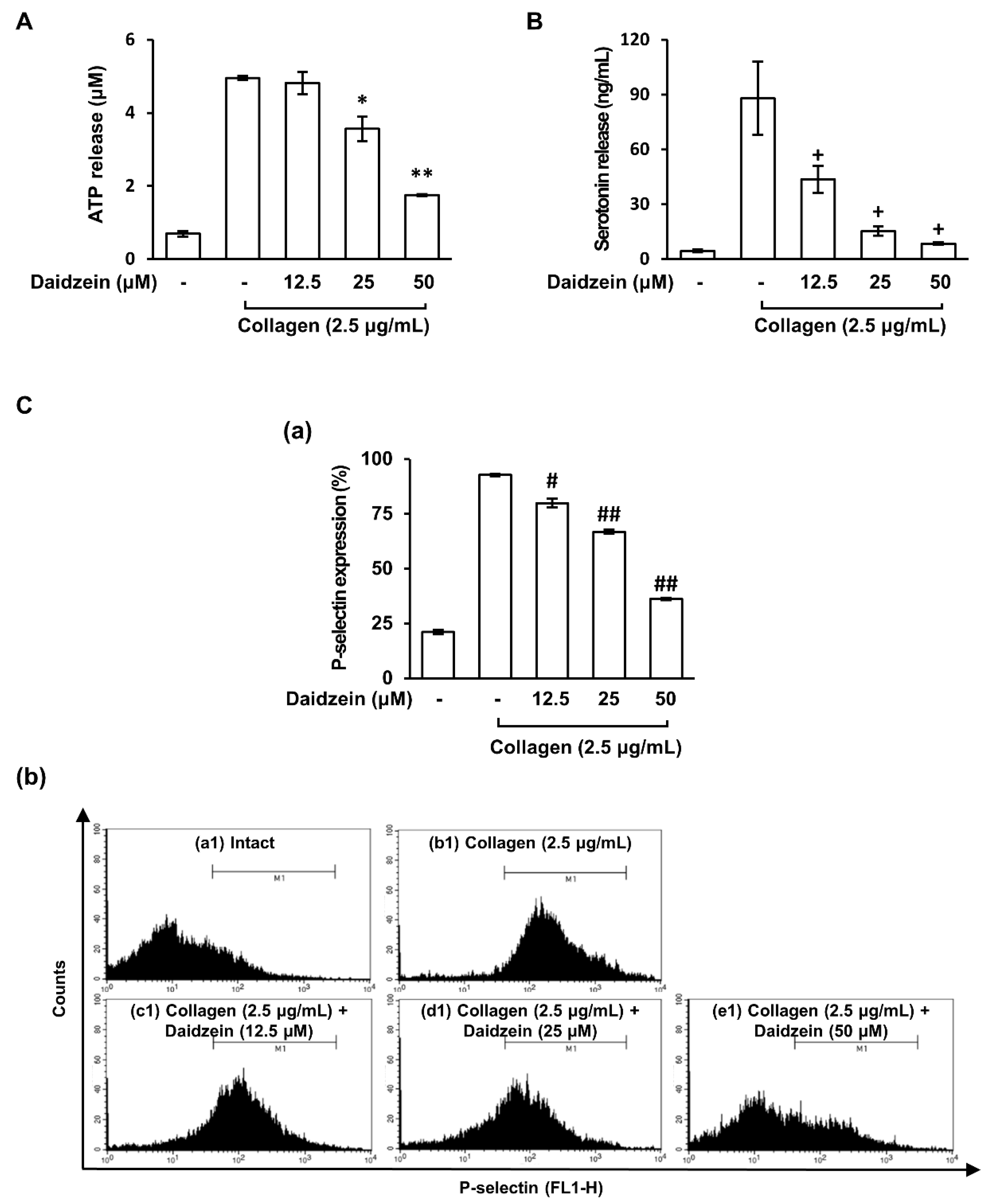
2.2. Effect of Daidzein on Granule Release by Collagen-Activated Platelets
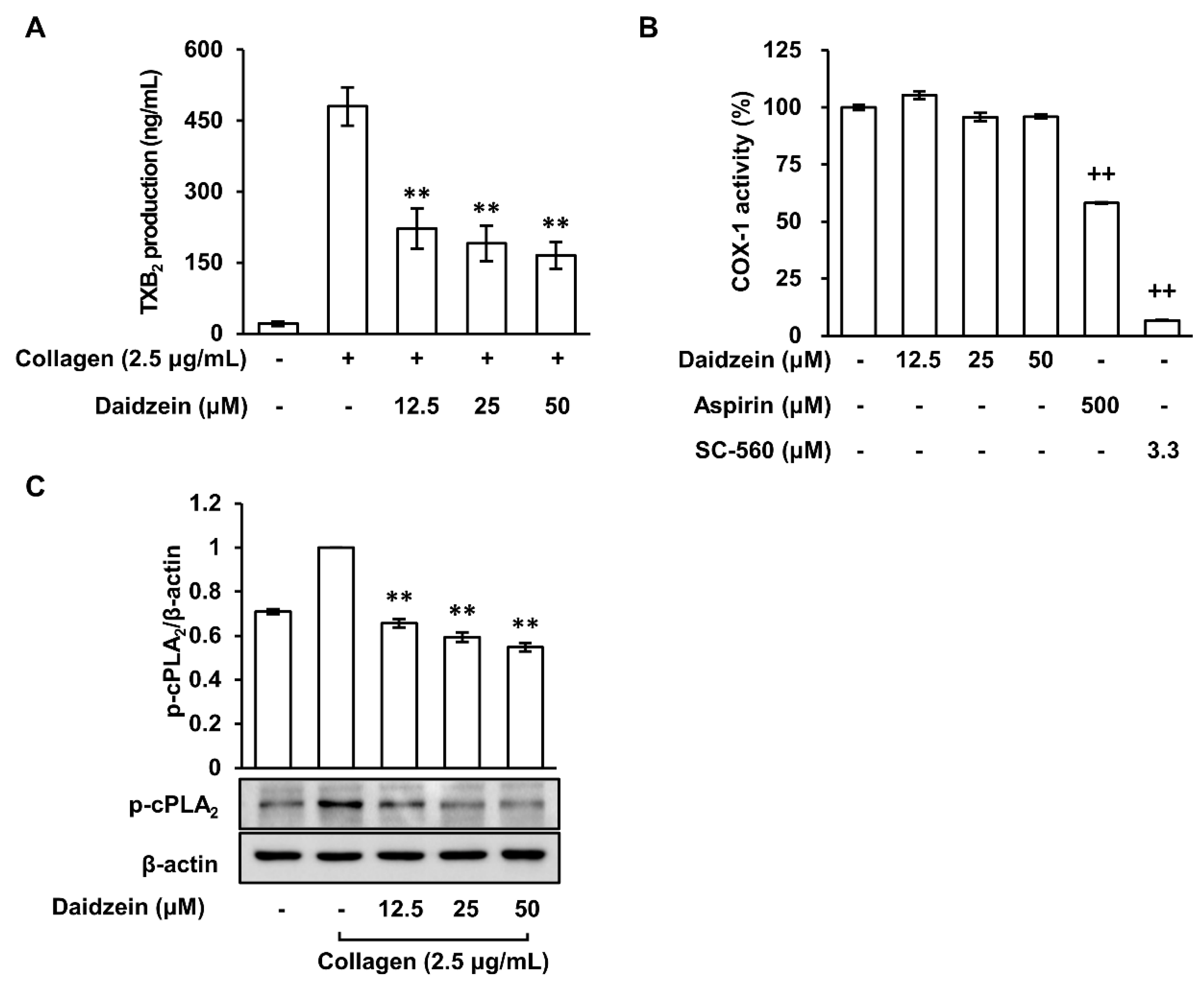
2.3. Daidzein Inhibited Thromboxane A2 Production Regardless of COX-1 Activity in Collagen-Induced Platelets
2.4. Inhibitory Effect of Daidzein on the Phosphorylation of Cytosolic Phospholipase A2
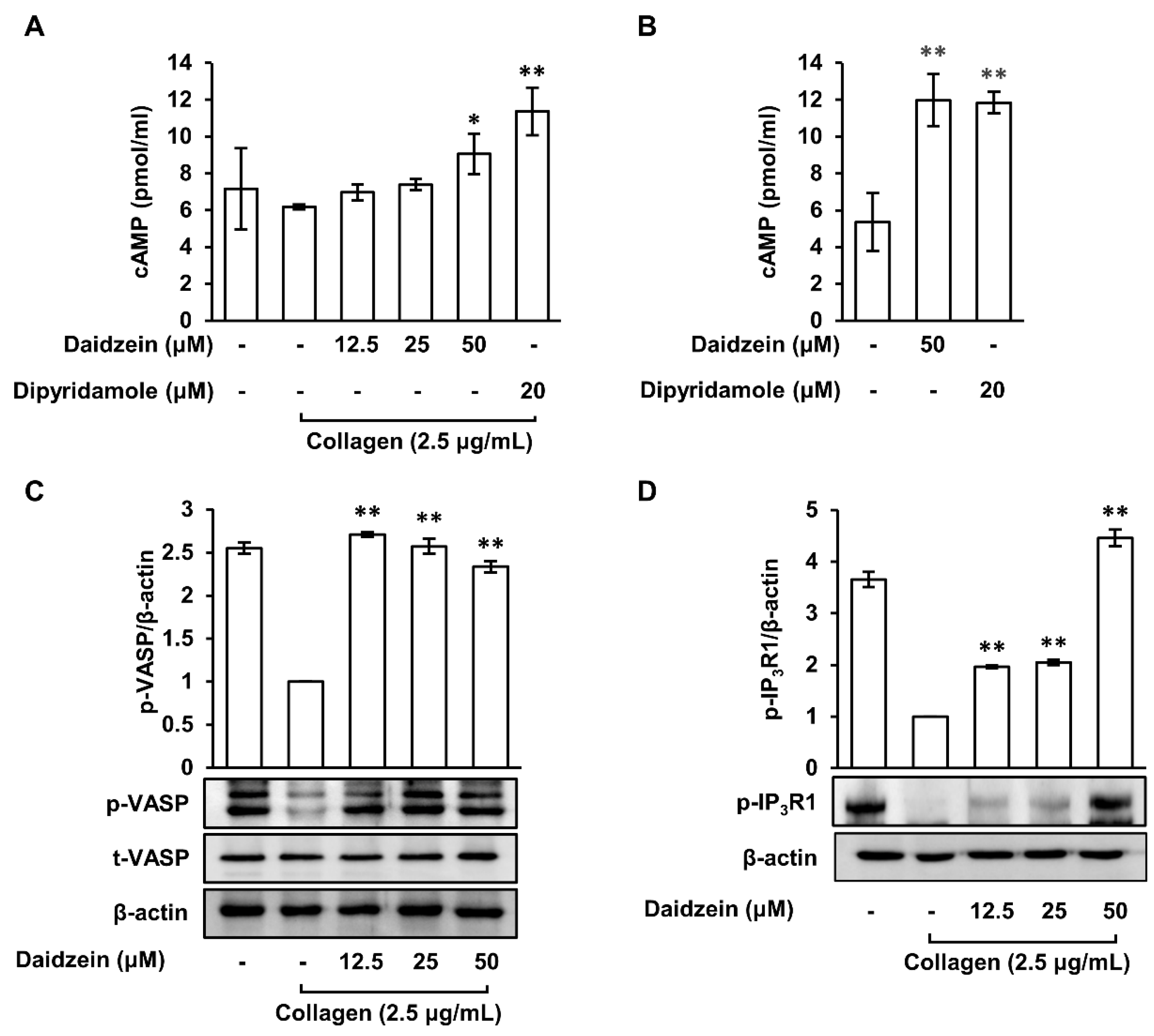
2.5. Effects of Daidzein on Cyclic Adenosine Monophosphate Levels and Phosphodiesterase Activity in Collagen-Induced Platelets
2.6. Effects of Daidzein on the Phosphorylations of VASP (Ser157) and IP3R1 in Collagen-Activated Platelets
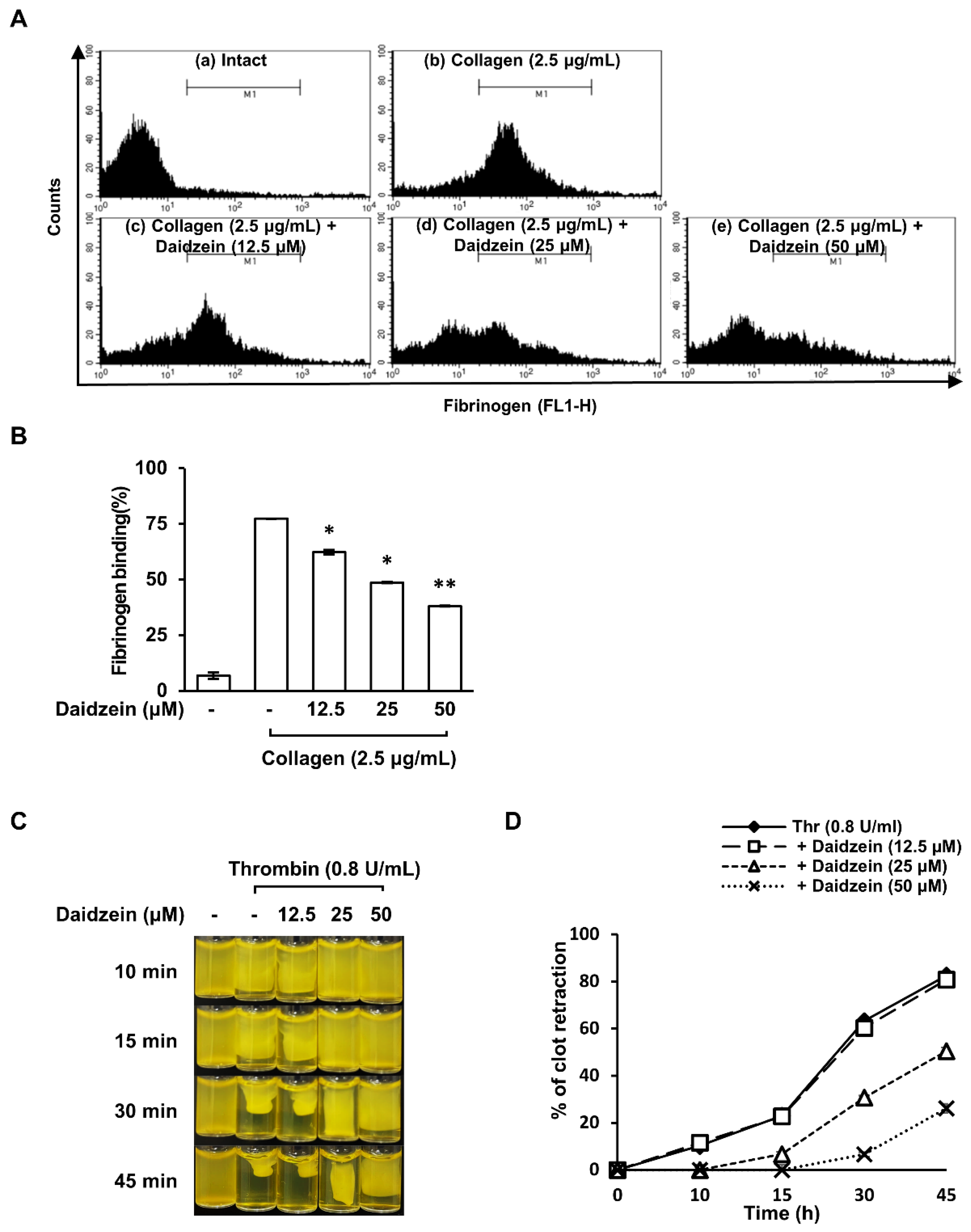
2.7. Effect of Daidzein on Fibrinogen Binding to Integrin αIIbβ3 on Collagen-Induced Platelets
2.8. Inhibitory Effect of Daidzein on Clot Retraction
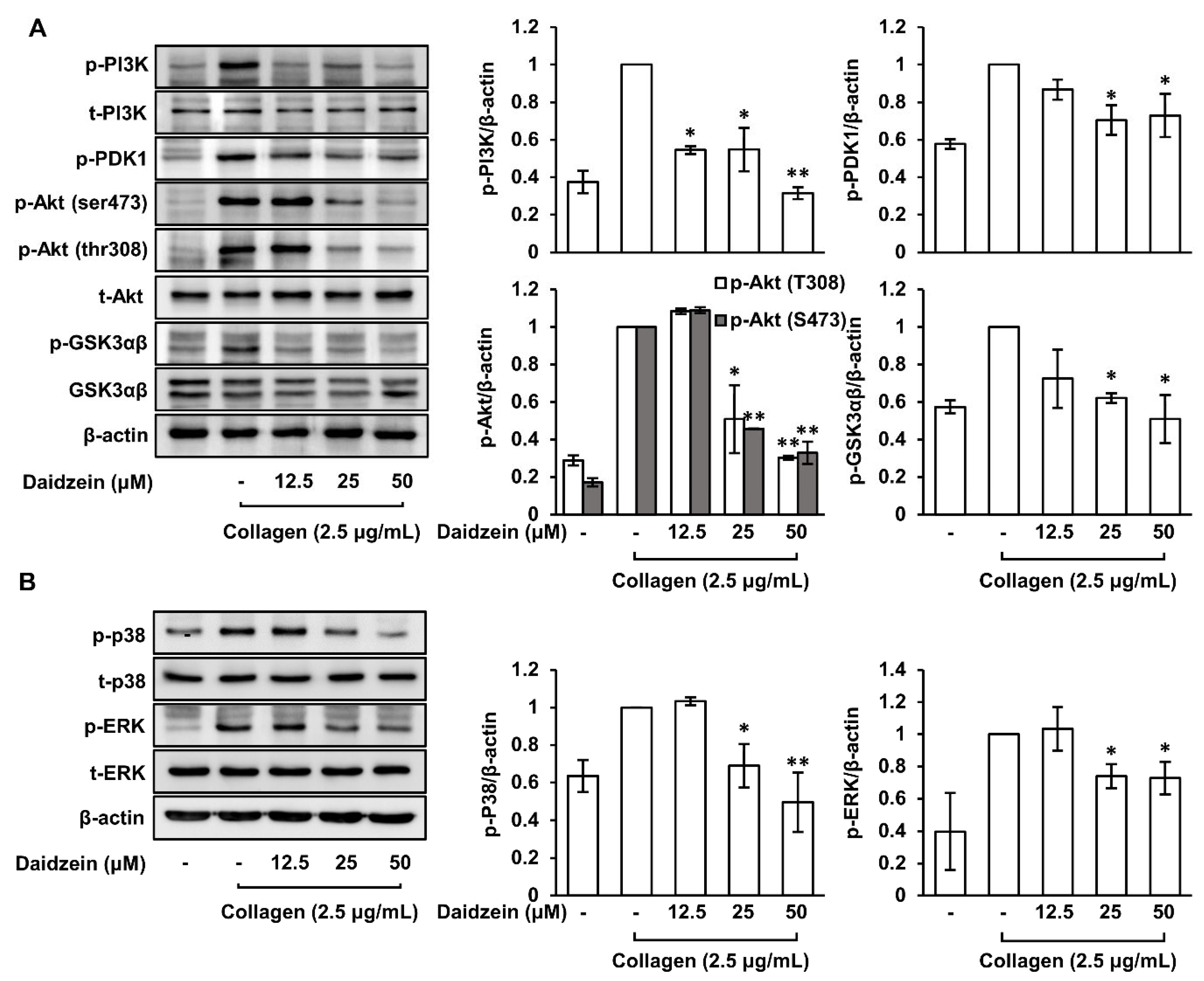
2.9. Inhibitory Effects of Daidzein on the Phosphorylations of PI3K/PDK1/Akt/GSK3αβ and MAPK (p38 and ERK) Pathways
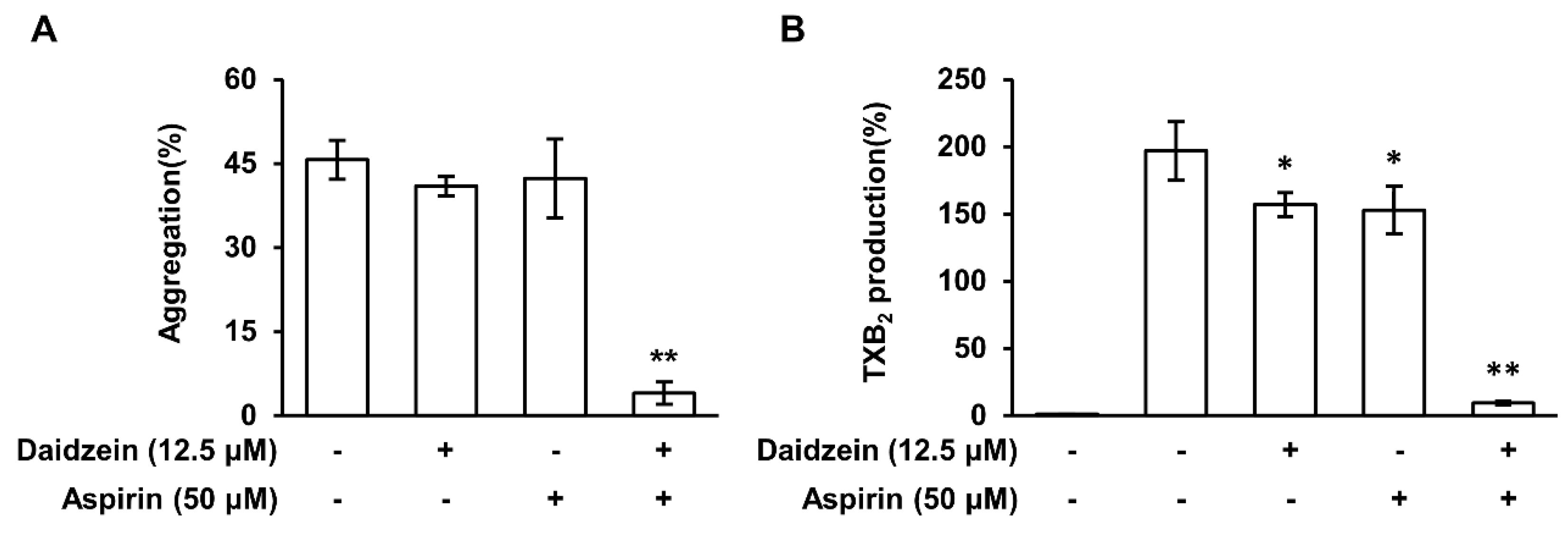
2.10. The Synergistic Effect of Daidzein and Aspirin
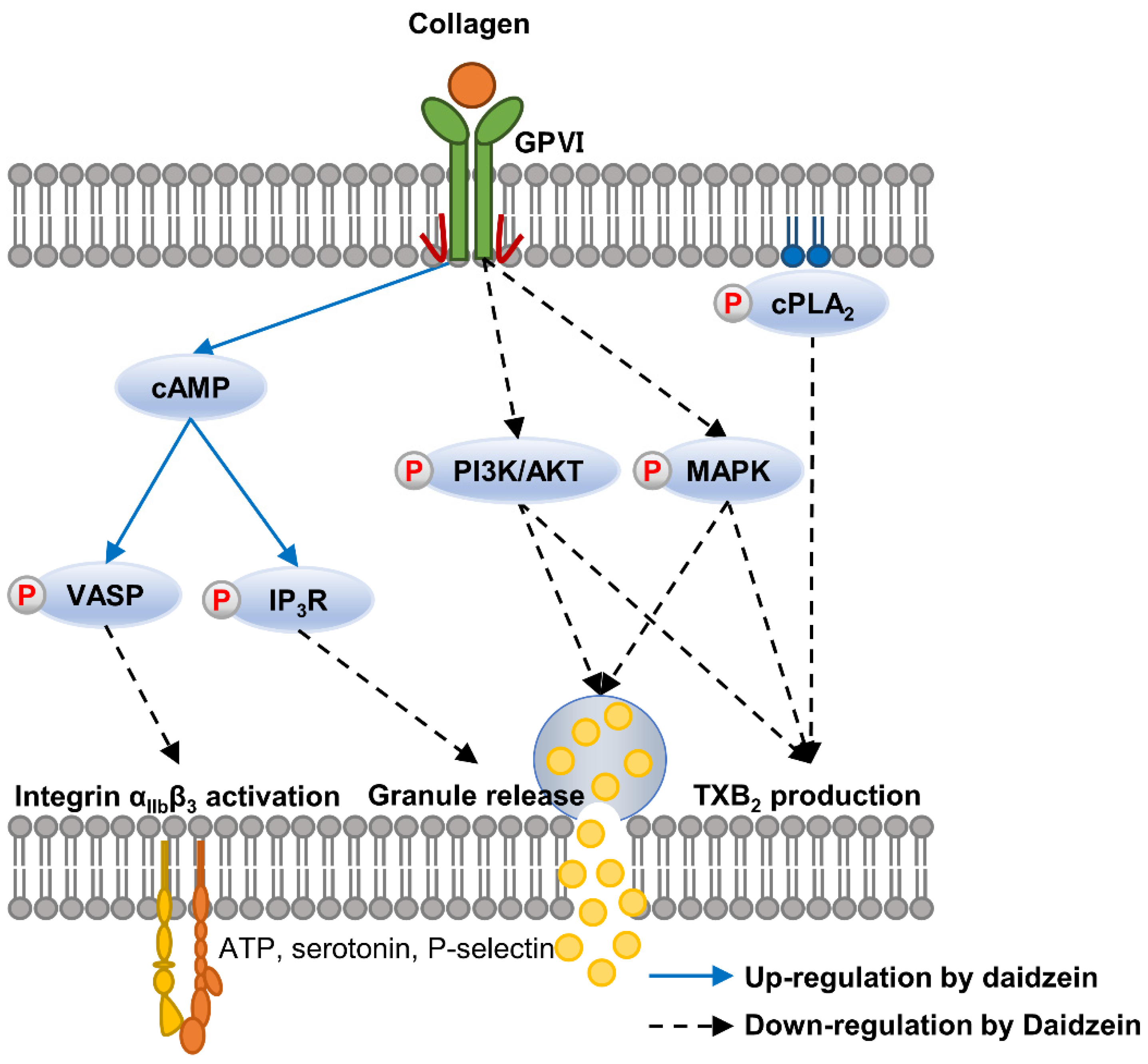
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Washed Human Platelets
4.3. Determination of Human Platelet Aggregation
4.4. Determination of ATP and Serotonin Release
4.5. Determination of TXA2 Production
4.6. Determination of COX-1 Activity
4.7. Determination of cAMP Levels
4.8. Flow Cytometry Analysis
4.9. Clot Retraction Assay
4.10. Immunoblotting Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Payne, R.A. Cardiovascular Risk. Br. J. Clin. Pharmacol. 2012, 74, 396–410. [Google Scholar] [CrossRef]
- Willoughby, S.; Holmes, A.; Loscalzo, J. Platelets and Cardiovascular Disease. Eur. J. Cardiovasc. Nurs. 2002, 1, 273–288. [Google Scholar] [CrossRef]
- Jackson, S.P. Arterial Thrombosis-Insidious, Unpredictable and Deadly. Nat. Med. 2011, 17, 1423–1436. [Google Scholar] [CrossRef]
- Yun, S.H.; Sim, E.H.; Goh, R.Y.; Park, J.I.; Han, J.Y. Platelet Activation: The Mechanisms and Potential Biomarkers. Biomed Res. Int. 2016, 2016, 9060143. [Google Scholar] [CrossRef]
- Born, G. Platelets: Past, Present and Future. Handb. Exp. Pharmacol. 2012, 210, 59–85. [Google Scholar]
- Jung, S.H.; Han, J.H.; Park, H.S.; Lee, J.J.; Yang, S.Y.; Kim, Y.H.; Heo, K.S.; Myung, C.S. Inhibition of Collagen-Induced Platelet Aggregation by the Secobutanolide Secolincomolide a from Lindera Obtusiloba Blume. Front. Pharmacol. 2017, 8, 560. [Google Scholar] [CrossRef]
- Jandrot-Perrus, M.; Busfield, S.; Lagrue, A.H.; Xiong, X.; Debili, N.; Chickering, T.; Le Couedic, J.P.; Goodearl, A.; Dussault, B.; Fraser, C.; et al. Cloning, Characterization, and Functional Studies of Human and Mouse Glycoprotein VI: A Platelet-Specific Collagen Receptor from the Immunoglobulin Superfamily. Blood 2000, 96, 1798–1807. [Google Scholar] [CrossRef]
- Flaumenhaft, R.; Sharda, A. The Life Cycle of Platelet Granules. F1000Research 2018, 7, 236. [Google Scholar]
- Golebiewska, E.M.; Poole, A.W. Platelet Secretion: From Haemostasis to Wound Healing and Beyond. Blood Rev. 2015, 29, 153–162. [Google Scholar] [CrossRef]
- Blair, P.; Flaumenhaft, R. Platelet α–Granules: Basic Biology and Clinical Correlates. Blood Rev. 2009, 23, 177–189. [Google Scholar] [CrossRef]
- Amalia, L. The Role of Platelet-Selectin as a Marker of Thrombocyte Aggregation on Cerebral Sinus Venous Thrombosis. J. Blood Med. 2022, 13, 267–274. [Google Scholar] [CrossRef]
- Khan, S.A.; Ilies, M.A. The Phospholipase A2 Superfamily: Structure, Isozymes, Catalysis, Physiologic and Pathologic Roles. Int. J. Mol. Sci. 2023, 24, 1353. [Google Scholar] [CrossRef]
- Szczuko, M.; Kozioł, I.; Kotlęga, D.; Brodowski, J.; Drozd, A. The Role of Thromboxane in the Course and Treatment of Ischemic Stroke: Review. Int. J. Mol. Sci. 2021, 22, 11644. [Google Scholar] [CrossRef]
- Yoon, S.; Kwon, H.; Shin, J.; Rhee, M.H.; Park, C.; Lee, D. Anti-Thrombotic Effects of Artesunate through Regulation of cAMP and PI3K / MAPK Pathway on Human Platelets. Int. J. Mol. Sci. 2022, 23, 1586. [Google Scholar] [CrossRef]
- Varga-Szabo, D.; Braun, A.; Nieswandt, B. Calcium Signaling in Platelets. J. Thromb. Haemost. 2009, 7, 1057–1066. [Google Scholar] [CrossRef]
- Huang, J.; Li, X.; Shi, X.; Zhu, M.; Wang, J.; Huang, S.; Huang, X.; Wang, H.; Li, L.; Deng, H.; et al. Platelet Integrin αIIbβ3: Signal Transduction, Regulation, and Its Therapeutic Targeting. J. Hematol. Oncol. 2019, 12, 26. [Google Scholar] [CrossRef]
- Bennett, J.S. Structure and Function of the Platelet Integrin αIIbβ3. J. Clin. Investig. 2005, 115, 3363–3369. [Google Scholar] [CrossRef]
- Tucker, K.L.; Sage, T.; Gibbins, J.M. Clot Retraction. Methods Mol. Biol. 2012, 788, 101–107. [Google Scholar]
- Long, H.Z.; Cheng, Y.; Zhou, Z.W.; Luo, H.Y.; Wen, D.D.; Gao, L.C. PI3K/AKT Signal Pathway: A Target of Natural Products in the Prevention and Treatment of Alzheimer’s Disease and Parkinson’s Disease. Front. Pharmacol. 2021, 12, 648636. [Google Scholar] [CrossRef]
- Yin, H.; Stojanovic, A.; Hay, N.; Du, X. The Role of Akt in the Signaling Pathway of the Glycoprotein Ib-IX Induced Platelet Activation. Blood 2008, 111, 658–665. [Google Scholar] [CrossRef]
- Niu, H.; Chen, X.; Gruppo, R.A.; Li, D.; Wang, Y.; Zhang, L.; Wang, K.; Chai, W.; Sun, Y.; Ding, Z.; et al. Integrin αIIbβ3-Mediated PI3K/Akt Activation in Platelets. PLoS ONE 2012, 7, e47356. [Google Scholar] [CrossRef]
- Riaz, M.N. Soy Beans: Processing; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; pp. 48–53. [Google Scholar]
- Liu, M.; Wang, G.; Xu, R.; Shen, C.; Ni, H.; Lai, R. Soy Isoflavones Inhibit Both GPIB-IX Signaling and αIIbβ3 Outside-in Signaling via 14-3-3ζ in Platelet. Molecules 2021, 26, 4911. [Google Scholar] [CrossRef]
- Beretz, A.; Cazenave, J.P.; Anton, R. Inhibition of Aggregation and Secretion of Human Platelets by Quercetin and Other Flavonoids: Structure-Activity Relationships. Agents Actions 1982, 12, 382–387. [Google Scholar] [CrossRef]
- Nakashima, S.; Koike, T.; Nozawa, Y. Genistein, a protein tyrosine kinase inhibitor, inhibits thromboxane A2-mediated human platelet responses. Mol. Pharmacol. 1991, 39, 475–480. [Google Scholar]
- Dobrydneva, Y.; Williams, R.L.; Morris, G.Z.; Blackmore, P.F. Dietary Phytoestrogens and Their Synthetic Structural Analogues as Calcium Channel Blockers in Human Platelets. J. Cardiovasc. Pharmacol. 2002, 40, 399–410. [Google Scholar] [CrossRef]
- Guerrero, J.A.; Lozano, M.L.; Castillo, J.; Benavente-García, O.; Vicente, V.; Rivera, J. Flavonoids Inhibit Platelet Function through Binding to the Thromboxane A2 Receptor. J. Thromb. Haemost. 2005, 3, 369–376. [Google Scholar] [CrossRef]
- Alshehri, M.M.; Sharifi-Rad, J.; Herrera-Bravo, J.; Jara, E.L.; Salazar, L.A.; Kregiel, D.; Uprety, Y.; Akram, M.; Iqbal, M.; Martorell, M.; et al. Therapeutic Potential of Isoflavones with an Emphasis on Daidzein. Oxid. Med. Cell Longev. 2021, 2021, 6331630. [Google Scholar] [CrossRef]
- Tomić, M.; Micov, A.; Pecikoza, U.; Stepanović-Petrović, R. Clinical Uses of Nonsteroidal Anti-Inflammatory Drugs (NSAIDs) and Potential Benefits of NSAIDs Modified-Release Preparations. In Microsized Nanosized Carriers Nonsteroidal Anti-Inflammatory. Drugs Formulation Challenges Potential Benefits; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–29. [Google Scholar]
- Shin, J.H.; Kwon, H.W.; Rhee, M.H.; Park, H.J. Inhibitory Effects of Thromboxane A2 Generation by Ginsenoside Ro due to Attenuation of Cytosolic Phospholipase A2 Phosphorylation and Arachidonic Acid Release. J. Ginseng Res. 2019, 43, 236–241. [Google Scholar] [CrossRef]
- Noe, L.; Peeters, K.; Izzi, B.; Van Geet, C.; Freson, K. Regulators of Platelet cAMP Levels: Clinical and Therapeutic Implications. Curr. Med. Chem. 2010, 17, 2897–2905. [Google Scholar] [CrossRef]
- Guidetti, G.F.; Canobbio, I.; Torti, M. PI3K/Akt in Platelet Integrin Signaling and Implications in Thrombosis. Adv. Biol. Regul. 2015, 59, 36–52. [Google Scholar] [CrossRef]
- Payrastre, B.; Missy, K.; Trumel, C.; Bodin, S.; Plantavid, M.; Chap, H. The Integrin αIIbβ3 in Human Platelet Signal Transduction. Biochem. Pharmacol. 2000, 60, 1069–1074. [Google Scholar] [CrossRef]
- Flevaris, P.; Li, Z.; Zhang, G.; Zheng, Y.; Liu, J.; Du, X. Two Distinct Roles of Mitogen-Activated Protein Kinases in Platelets and a Novel Rac1-MAPK-Dependent Integrin Outside-in Retractile Signaling Pathway. Blood 2009, 113, 893–901. [Google Scholar] [CrossRef]
- Jankowski, J.; Floege, J.; Fliser, D.; Böhm, M.; Marx, N. Cardiovascular Disease in Chronic Kidney Disease Pathophysiological Insights and Therapeutic Options. Circulation 2021, 143, 1157–1172. [Google Scholar] [CrossRef]
- Periayah, M.H.; Halim, A.S.; Saad, A.Z.M. Mechanism Action of Platelets and Crucial Blood Coagulation Pathways in Hemostasis. Int. J. Hematol. Stem Cell Res. 2017, 11, 319–327. [Google Scholar]
- Wang, L.; Tang, C. Targeting Platelet in Atherosclerosis Plaque Formation: Current Knowledge and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 9760. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Padró, T.; Vilahur, G. Atherosclerosis, Platelets and Thrombosis in Acute Ischaemic Heart Disease. Eur. Hear J. Acute Cardiovasc. Care. 2012, 1, 60–74. [Google Scholar] [CrossRef] [PubMed]
- Fitch-Tewfik, J.L.; Flaumenhaft, R. Platelet Granule Exocytosis: A Comparison with Chromaffin Cells. Front. Endocrinol. 2013, 4, 77. [Google Scholar] [CrossRef]
- Gresele, P.; Momi, S.; Falcinelli, E. Anti-Platelet Therapy: Phosphodiesterase Inhibitors. Br. J. Clin. Pharmacol. 2011, 72, 634–646. [Google Scholar] [CrossRef]
- Shin, J.H.; Kwon, H.W.; Cho, H.J.; Rhee, M.H.; Park, H.J. Vasodilator-Stimulated Phosphoprotein-Phosphorylation by Ginsenoside Ro Inhibits Fibrinogen Binding to αIIbβ3 in Thrombin-Induced Human Platelets. J. Ginseng Res. 2016, 40, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Yuan, Z.; Tian, X.; Xiong, X.; Guo, F.; Lin, Z.; Qin, Z. Pimpinellin Inhibits Collagen-Induced Platelet Aggregation and Activation through Inhibiting Granule Secretion and PI3K/Akt Pathway. Front. Pharmacol. 2021, 12, 706363. [Google Scholar] [CrossRef]
- Rukoyatkina, N.; Mindukshev, I.; Walter, U.; Gambaryan, S. Dual Role of the P38 MAPK/cPLA2 Pathway in the Regulation of Platelet Apoptosis Induced by ABT-737 and Strong Platelet Agonists. Cell Death Dis. 2013, 4, e931. [Google Scholar] [CrossRef] [PubMed]
- Nam, G.S.; Kim, S.; Kwon, Y.S.; Kim, M.K.; Nam, K.S. A New Function for MAP4K4 Inhibitors during Platelet Aggregation and Platelet-Mediated Clot Retraction. Biochem. Pharmacol. 2021, 188, 114519. [Google Scholar] [CrossRef] [PubMed]
- Borer, J.S.; Simon, L.S. Cardiovascular and Gastrointestinal Effects of COX-2 Inhibitors and NSAIDs: Achieving a Balance. Arthritis Res. Ther. 2005, 7, 14–22. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, H.-J.; Nam, G.-S.; Nam, K.-S. Daidzein Inhibits Human Platelet Activation by Downregulating Thromboxane A2 Production and Granule Release, Regardless of COX-1 Activity. Int. J. Mol. Sci. 2023, 24, 11985. https://doi.org/10.3390/ijms241511985
Hong H-J, Nam G-S, Nam K-S. Daidzein Inhibits Human Platelet Activation by Downregulating Thromboxane A2 Production and Granule Release, Regardless of COX-1 Activity. International Journal of Molecular Sciences. 2023; 24(15):11985. https://doi.org/10.3390/ijms241511985
Chicago/Turabian StyleHong, Hyun-Jin, Gi-Suk Nam, and Kyung-Soo Nam. 2023. "Daidzein Inhibits Human Platelet Activation by Downregulating Thromboxane A2 Production and Granule Release, Regardless of COX-1 Activity" International Journal of Molecular Sciences 24, no. 15: 11985. https://doi.org/10.3390/ijms241511985