
NCOA7 Regulates Growth and Metastasis of Clear Cell Renal Cell Carcinoma via MAPK/ERK Signaling Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
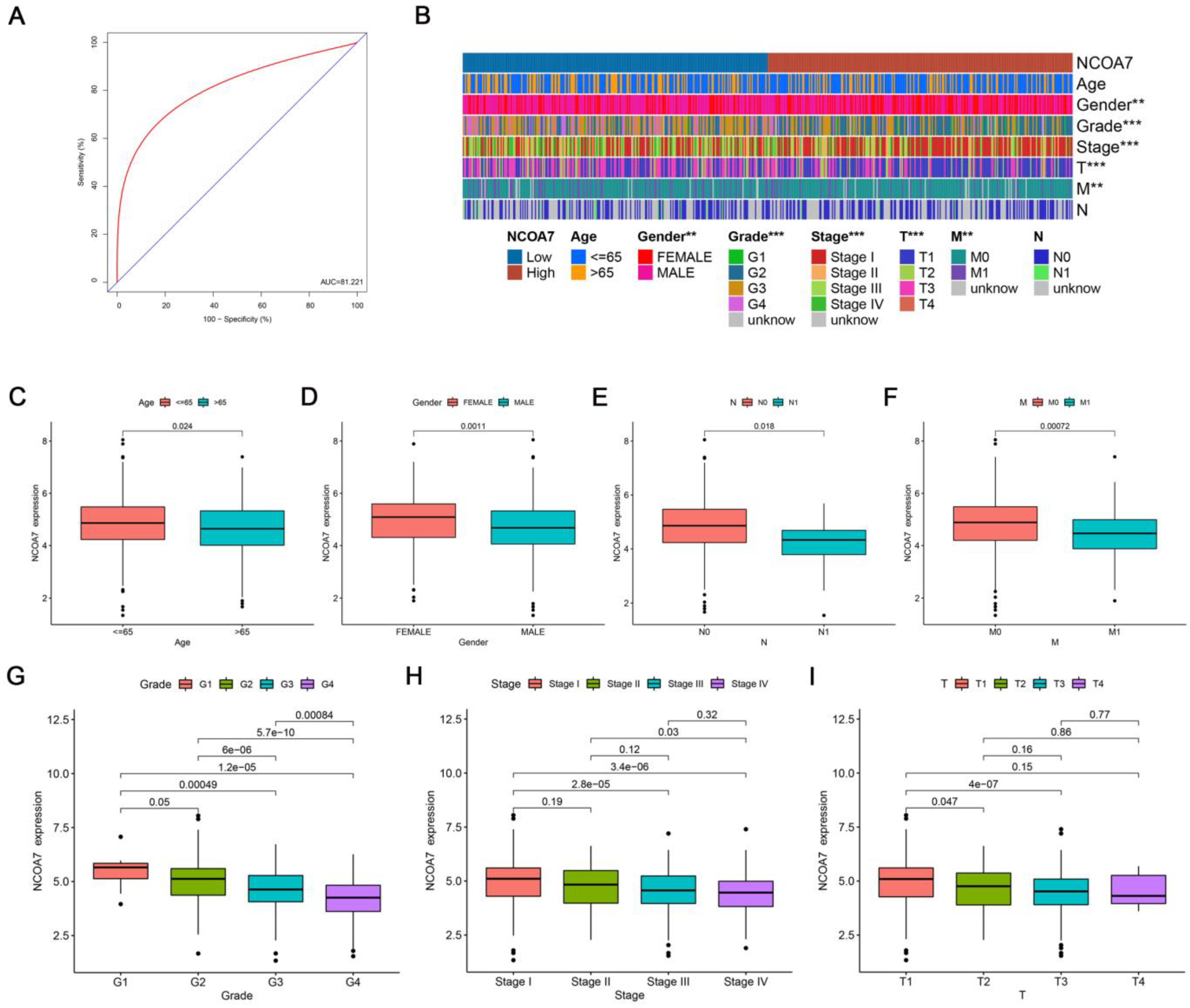
2.1. Analysis of the Expression Level, Prognostic Value, and Diagnosis of NCOA7 in Three Subtypes of RCC
2.2. Correlation between the Expression Level of NCOA7 Regarding Clinicopathological Features and Prognosis
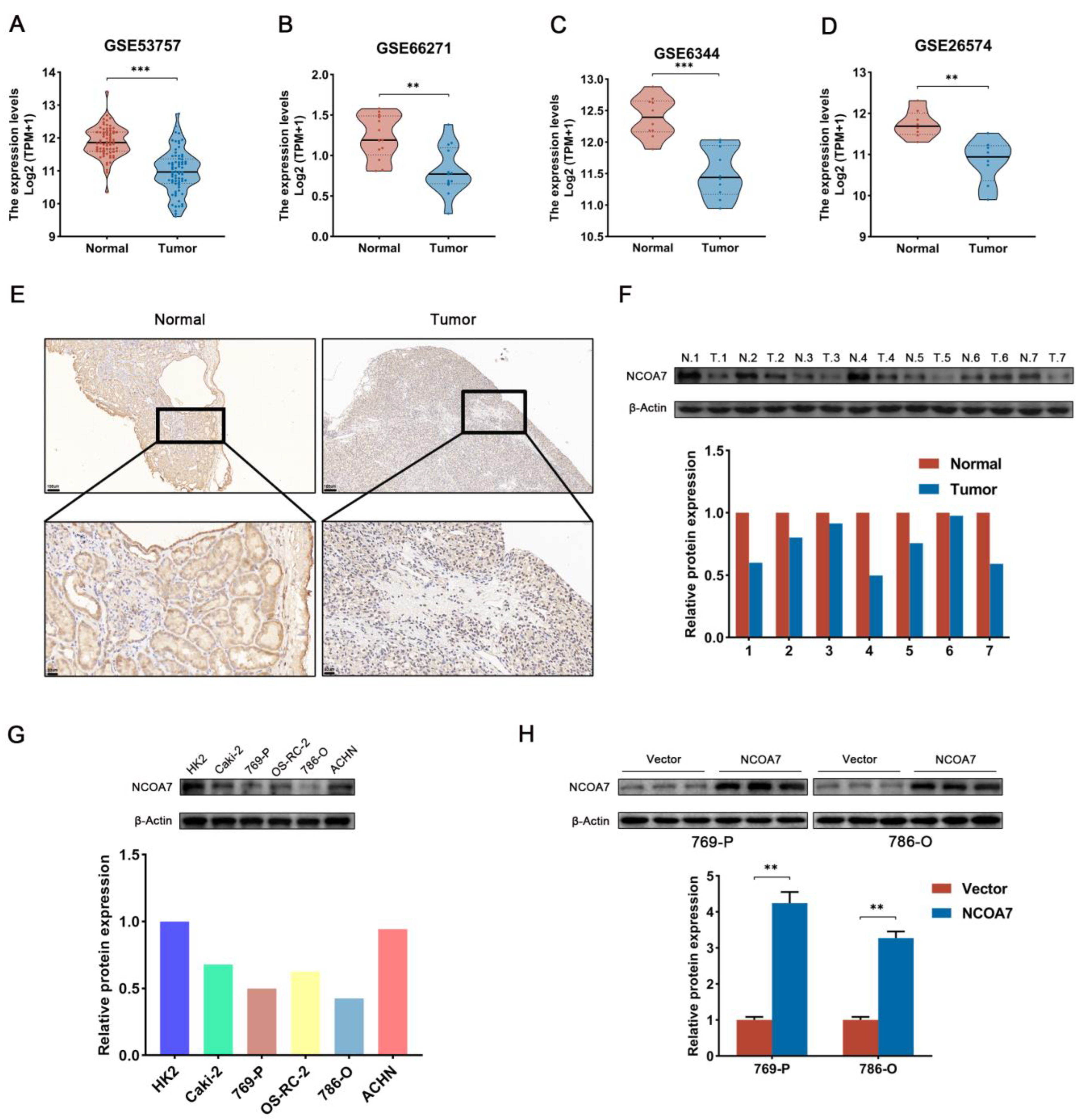
2.3. The Expression of NCOA7 Is Downregulated in ccRCC and Cell Lines
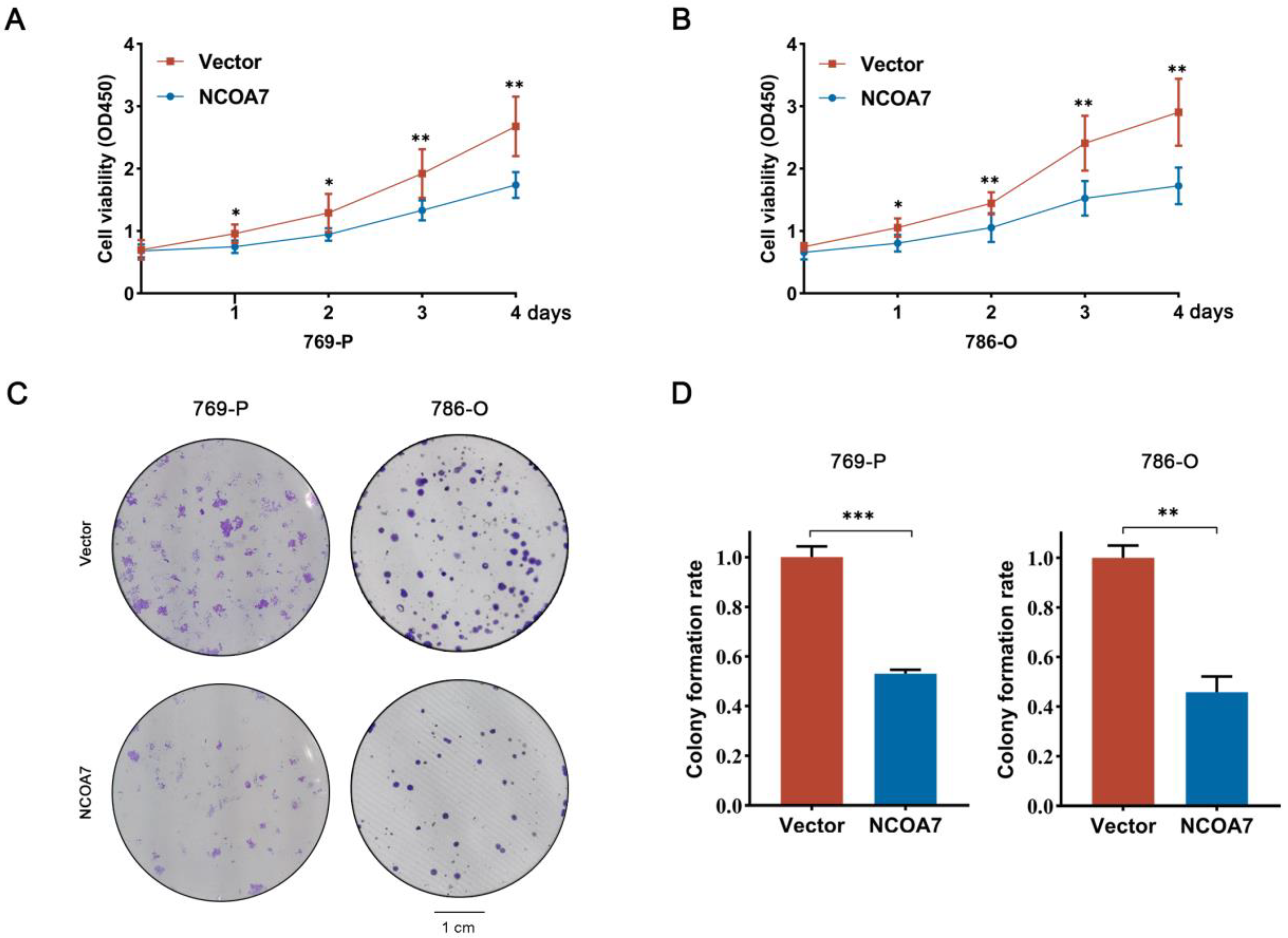
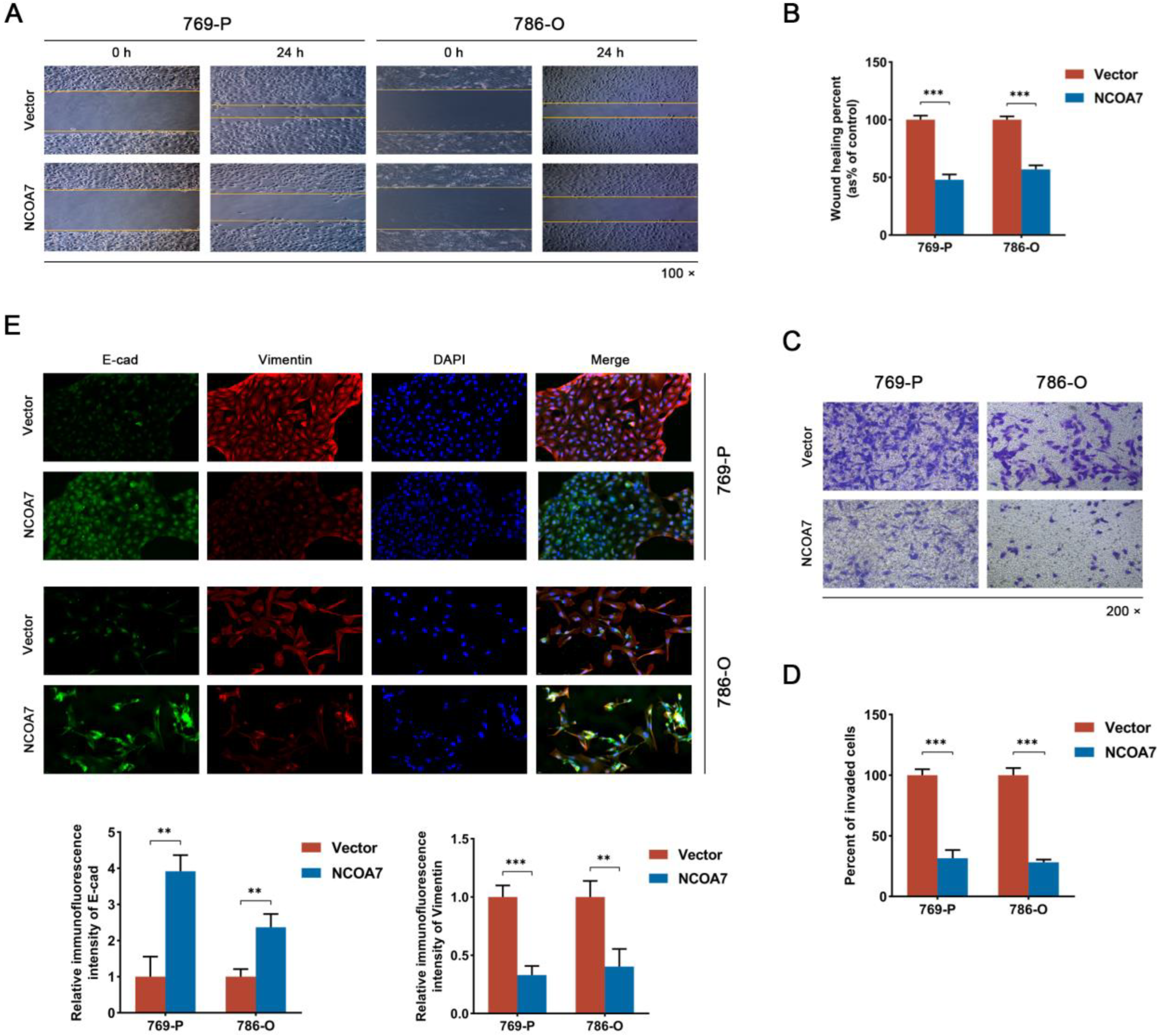
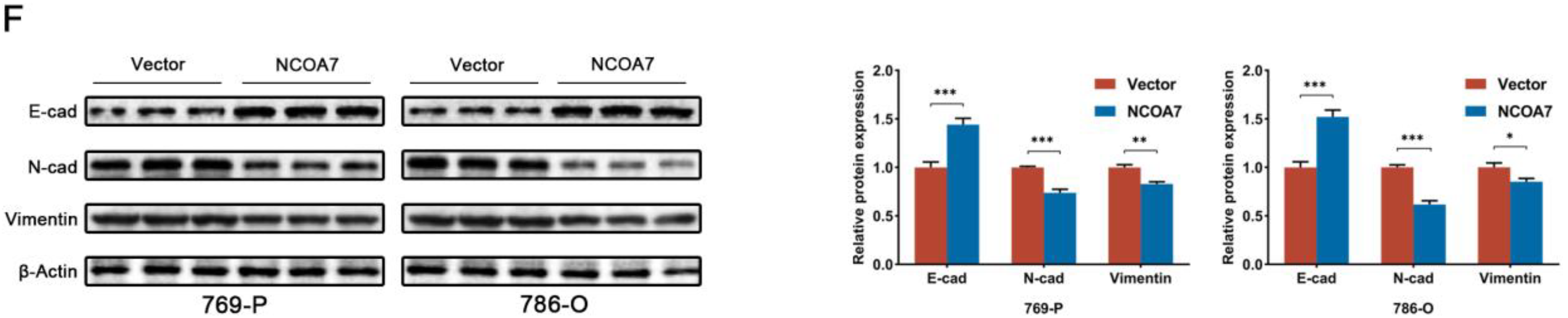
2.4. The Overexpression of NCOA7 Inhibits the Proliferation, Migration, and Invasion of 769-P and 786-O Cells In Vitro
2.5. The Effect of NCOA7 Overexpression on Cycle and Apoptosis in 769-P and 786-O Cells
2.6. MAPK/ERK Signaling Pathway Is Involved in NCOA7-Regulated ccRCC In Vitro
2.7. Activation of the MAPK/ERK Pathway Attenuates the Role of NCOA7
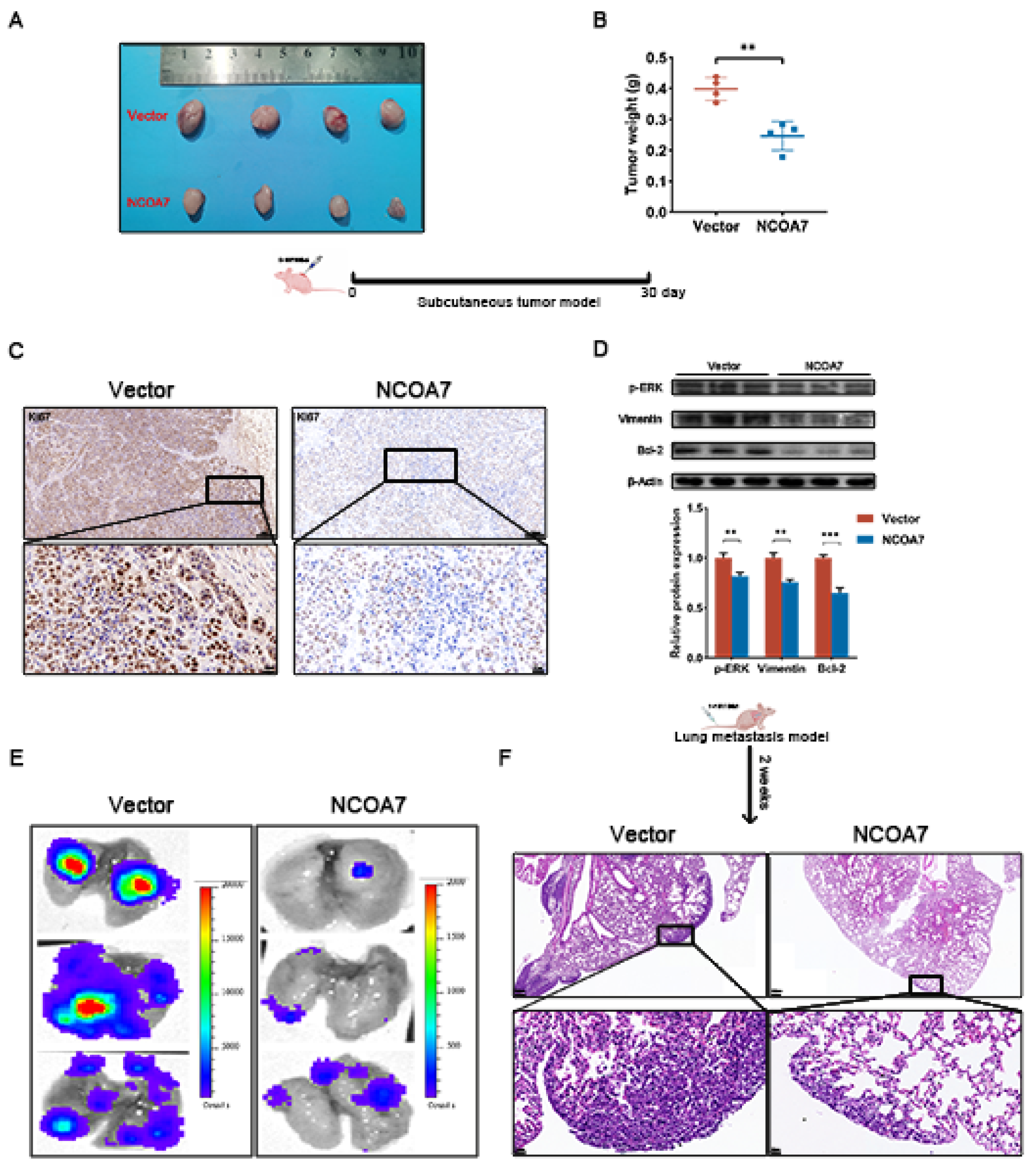
2.8. NCOA7 Inhibits Tumorigenesis and Lung Metastasis of ccRCC In Vivo
3. Discussion
4. Materials and Methods
4.1. Data Sources
4.2. Differential Expression Analysis of NCOA7
4.3. Analysis of Prognostic Value, Diagnosis, and Clinicopathological Parameters Correlation
4.4. Clinical Tissue Specimens
4.5. Cell Culture
4.6. Transfection of Lentiviral Plasmids
4.7. Cell Viability Assay
4.8. Colony Formation Assay
4.9. Wound Healing Assay
4.10. Transwell Invasion Assay
4.11. Western Blot Assay
4.12. Immunofluorescence Staining
4.13. Histology and Immunohistochemistry Assay
4.14. Flow Cytometry
4.15. In Vivo Assays
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, M. Recent developments in small molecule therapies for renal cell carcinoma. Eur. J. Med. Chem. 2017, 142, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Turajlic, S.; Swanton, C.; Boshsoff, C. Kidney cancer: The next decade. J. Exp. Med. 2018, 215, 2477–2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, D.M.; Agarwal, N.; Vaishampayan, U. Evolving Treatment Paradigm in Metastatic Renal Cell Carcinoma. Am. Soc. Clin. Oncol. Educ. Book 2017, 37, 319–329. [Google Scholar] [CrossRef]
- Ingels, A.; Campi, R.; Capitanio, U.; Amparore, D.; Bertolo, R.; Carbonara, U.; Erdem, S.; Kara, O.; Klatte, T.; Kriegmair, M.C.; et al. Complementary roles of surgery and systemic treatment in clear cell renal cell carcinoma. Nat. Rev. Urol. 2022, 19, 391–418. [Google Scholar] [CrossRef]
- Makhov, P.; Joshi, S.; Ghatalia, P.; Kutikov, A.; Uzzo, R.G.; Kolenko, V.M. Resistance to Systemic Therapies in Clear Cell Renal Cell Carcinoma: Mechanisms and Management Strategies. Mol. Cancer Ther. 2018, 17, 1355–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Dong, X.; Yap, J.; Hu, J. The MAPK and AMPK signalings: Interplay and implication in targeted cancer therapy. J. Hematol. Oncol. 2020, 13, 113. [Google Scholar] [CrossRef]
- Ullah, R.; Yin, Q.; Snell, A.H.; Wan, L. RAF-MEK-ERK pathway in cancer evolution and treatment. Semin. Cancer Biol. 2022, 85, 123–154. [Google Scholar] [CrossRef]
- Peluso, I.; Yarla, N.S.; Ambra, R.; Pastore, G.; Perry, G. MAPK signalling pathway in cancers: Olive products as cancer preventive and therapeutic agents. Semin. Cancer Biol. 2019, 56, 185–195. [Google Scholar] [CrossRef]
- Simanshu, D.K.; Nissley, D.V.; McCormick, F. RAS Proteins and Their Regulators in Human Disease. Cell 2017, 170, 17–33. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Ding, Y.; Luo, W.M.; Bender, S.; Qian, C.N.; Kort, E.; Zhang, Z.F.; VandenBeldt, K.; Duesbery, N.S.; Resau, J.H.; et al. Inhibition of MAPK kinase signaling pathways suppressed renal cell carcinoma growth and angiogenesis in vivo. Cancer Res. 2008, 68, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.Y.; Wang, J.; Zhang, J.Q.; Cai, H.M. Human circular RNA hsa_circRNA_101705 (circTXNDC11) regulates renal cancer progression by regulating MAPK/ERK pathway. Bioengineered 2021, 12, 4432–4441. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, Y.; Jiang, J.; Yin, C.; Shi, B. ADAM12 promotes clear cell renal cell carcinoma progression and triggers EMT via EGFR/ERK signaling pathway. J. Transl. Med. 2023, 21, 56. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, S.; Liu, R.; Chen, H.; Xu, B.; Xu, W.; Chen, M. Circular RNA circPOLR2A promotes clear cell renal cell carcinoma progression by facilitating the UBE3C-induced ubiquitination of PEBP1 and, thereby, activating the ERK signaling pathway. Mol. Cancer 2022, 21, 146. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Halachmi, S.; Brown, M. ERAP140, a conserved tissue-specific nuclear receptor coactivator. Mol. Cell Biol. 2002, 22, 3358–3372. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.; Gustafsson, J.A. The different roles of ER subtypes in cancer biology and therapy. Nat. Rev. Cancer 2011, 11, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Arai, H.; Ozaki, T.; Niizuma, H.; Nakamura, Y.; Ohira, M.; Takano, K.; Matsumoto, M.; Nakagawara, A. ERAP140/Nbla10993 is a novel favorable prognostic indicator for neuroblastoma induced in response to retinoic acid. Oncol. Rep. 2008, 19, 1381–1388. [Google Scholar]
- Lu, J.; Annunziata, F.; Sirvinskas, D.; Omrani, O.; Li, H.; Rasa, S.M.M.; Krepelova, A.; Adam, L.; Neri, F. Establishment and evaluation of module-based immune-associated gene signature to predict overall survival in patients of colon adenocarcinoma. J. Biomed. Sci. 2022, 29, 81. [Google Scholar] [CrossRef]
- Higginbotham, K.S.; Breyer, J.P.; Bradley, K.M.; Schuyler, P.A.; Plummer, W.D., Jr.; Freudenthal, M.E.; Trentham-Dietz, A.; Newcomb, P.A.; Sanders, M.E.; Page, D.L.; et al. A multistage association study identifies a breast cancer genetic locus at NCOA7. Cancer Res. 2011, 71, 3881–3888. [Google Scholar] [CrossRef] [Green Version]
- Suski, J.M.; Braun, M.; Strmiska, V.; Sicinski, P. Targeting cell-cycle machinery in cancer. Cancer Cell 2021, 39, 759–778. [Google Scholar] [CrossRef]
- O’Malley, B.W.; Kumar, R. Nuclear receptor coregulators in cancer biology. Cancer Res. 2009, 69, 8217–8222. [Google Scholar] [CrossRef] [Green Version]
- Han, S.J.; Lonard, D.M.; O’Malley, B.W. Multi-modulation of nuclear receptor coactivators through posttranslational modifications. Trends Endocrinol. Metab. 2009, 20, 8–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullner, J.; Lattrich, C.; Haring, J.; Gorse, R.; Ortmann, O.; Treeck, O. A polymorphism in the nuclear receptor coactivator 7 gene and breast cancer susceptibility. Oncol. Lett. 2012, 3, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Jiang, Y.; Yuan, Y.; Wang, P.; Li, X.; Chen, F.; Sun, C.; Zhao, H.; Zeng, X.; Jiang, L.; et al. MALDI imaging reveals NCOA7 as a potential biomarker in oral squamous cell carcinoma arising from oral submucous fibrosis. Oncotarget 2016, 7, 59987–60004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merkulova, M.; Paunescu, T.G.; Nair, A.V.; Wang, C.Y.; Capen, D.E.; Oliver, P.L.; Breton, S.; Brown, D. Targeted deletion of the Ncoa7 gene results in incomplete distal renal tubular acidosis in mice. Am. J. Physiol. Ren. Physiol. 2018, 315, F173–F185. [Google Scholar] [CrossRef] [Green Version]
- Blaise, M.; Alsarraf, H.M.; Wong, J.E.; Midtgaard, S.R.; Laroche, F.; Schack, L.; Spaink, H.; Stougaard, J.; Thirup, S. Crystal structure of the TLDc domain of oxidation resistance protein 2 from zebrafish. Proteins 2012, 80, 1694–1698. [Google Scholar] [CrossRef]
- Durand, M.; Kolpak, A.; Farrell, T.; Elliott, N.A.; Shao, W.; Brown, M.; Volkert, M.R. The OXR domain defines a conserved family of eukaryotic oxidation resistance proteins. BMC Cell Biol. 2007, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef]
- Alao, J.P. The regulation of cyclin D1 degradation: Roles in cancer development and the potential for therapeutic invention. Mol. Cancer 2007, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.T.; Liu, H.; Mao, M.J.; Tan, Y.; Mo, X.Q.; Meng, X.J.; Cao, M.T.; Zhong, C.Y.; Liu, Y.; Shan, H.; et al. Crosstalk between autophagy and epithelial-mesenchymal transition and its application in cancer therapy. Mol. Cancer 2019, 18, 101. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, V.; Brabletz, T.; Ceppi, P. Targeting EMT in Cancer with Repurposed Metabolic Inhibitors. Trends Cancer 2020, 6, 942–950. [Google Scholar] [CrossRef]
- Kim, E.K.; Choi, E.J. Compromised MAPK signaling in human diseases: An update. Arch. Toxicol. 2015, 89, 867–882. [Google Scholar] [CrossRef]
- Braicu, C.; Buse, M.; Busuioc, C.; Drula, R.; Gulei, D.; Raduly, L.; Rusu, A.; Irimie, A.; Atanasov, A.G.; Slaby, O.; et al. A Comprehensive Review on MAPK: A Promising Therapeutic Target in Cancer. Cancers 2019, 11, 1618. [Google Scholar] [CrossRef] [Green Version]
- Samatar, A.A.; Poulikakos, P.I. Targeting RAS-ERK signalling in cancer: Promises and challenges. Nat. Rev. Drug Discov. 2014, 13, 928–942. [Google Scholar] [CrossRef]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- von Roemeling, C.A.; Radisky, D.C.; Marlow, L.A.; Cooper, S.J.; Grebe, S.K.; Anastasiadis, P.Z.; Tun, H.W.; Copland, J.A. Neuronal pentraxin 2 supports clear cell renal cell carcinoma by activating the AMPA-selective glutamate receptor-4. Cancer Res. 2014, 74, 4796–4810. [Google Scholar] [CrossRef] [Green Version]
- Wotschofsky, Z.; Gummlich, L.; Liep, J.; Stephan, C.; Kilic, E.; Jung, K.; Billaud, J.N.; Meyer, H.A. Integrated microRNA and mRNA Signature Associated with the Transition from the Locally Confined to the Metastasized Clear Cell Renal Cell Carcinoma Exemplified by miR-146-5p. PLoS ONE 2016, 11, e0148746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumz, M.L.; Zou, H.; Kreinest, P.A.; Childs, A.C.; Belmonte, L.S.; LeGrand, S.N.; Wu, K.J.; Luxon, B.A.; Sinha, M.; Parker, A.S.; et al. Secreted frizzled-related protein 1 loss contributes to tumor phenotype of clear cell renal cell carcinoma. Clin. Cancer Res. 2007, 13, 4740–4749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ooi, A.; Wong, J.C.; Petillo, D.; Roossien, D.; Perrier-Trudova, V.; Whitten, D.; Min, B.W.; Tan, M.H.; Zhang, Z.; Yang, X.J.; et al. An antioxidant response phenotype shared between hereditary and sporadic type 2 papillary renal cell carcinoma. Cancer Cell 2011, 20, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Ke, S.; Chen, Q.; Zhou, J.; Guo, J.; Qiu, T. NCOA7 Regulates Growth and Metastasis of Clear Cell Renal Cell Carcinoma via MAPK/ERK Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 11584. https://doi.org/10.3390/ijms241411584
Guo J, Ke S, Chen Q, Zhou J, Guo J, Qiu T. NCOA7 Regulates Growth and Metastasis of Clear Cell Renal Cell Carcinoma via MAPK/ERK Signaling Pathway. International Journal of Molecular Sciences. 2023; 24(14):11584. https://doi.org/10.3390/ijms241411584
Chicago/Turabian StyleGuo, Jiayu, Shuai Ke, Qi Chen, Jiangqiao Zhou, Jia Guo, and Tao Qiu. 2023. "NCOA7 Regulates Growth and Metastasis of Clear Cell Renal Cell Carcinoma via MAPK/ERK Signaling Pathway" International Journal of Molecular Sciences 24, no. 14: 11584. https://doi.org/10.3390/ijms241411584