
Innovative Pre-Clinical Data Using Peptides to Intervene in the Evolution of Pulmonary Fibrosis
 , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
2. Results
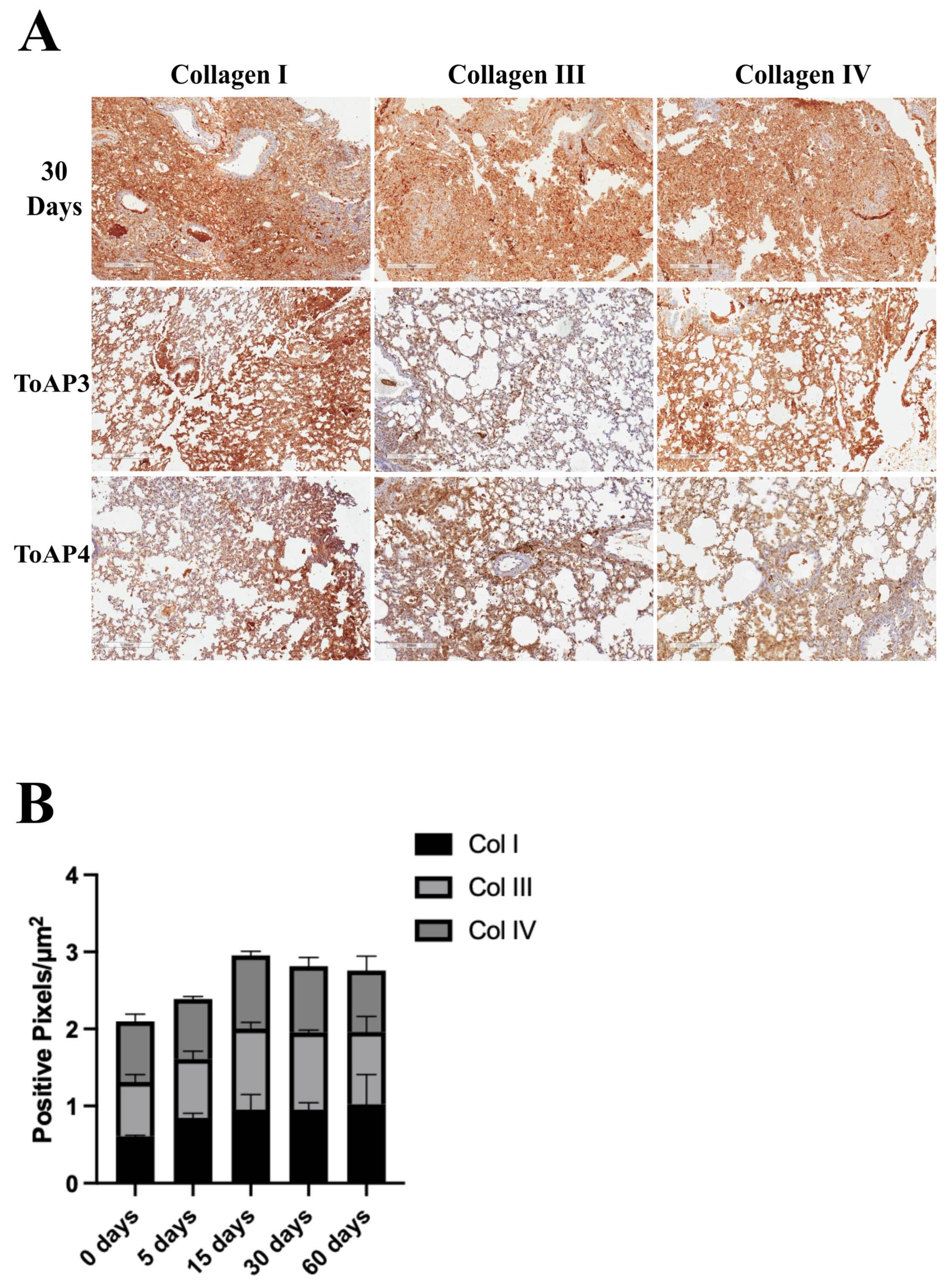
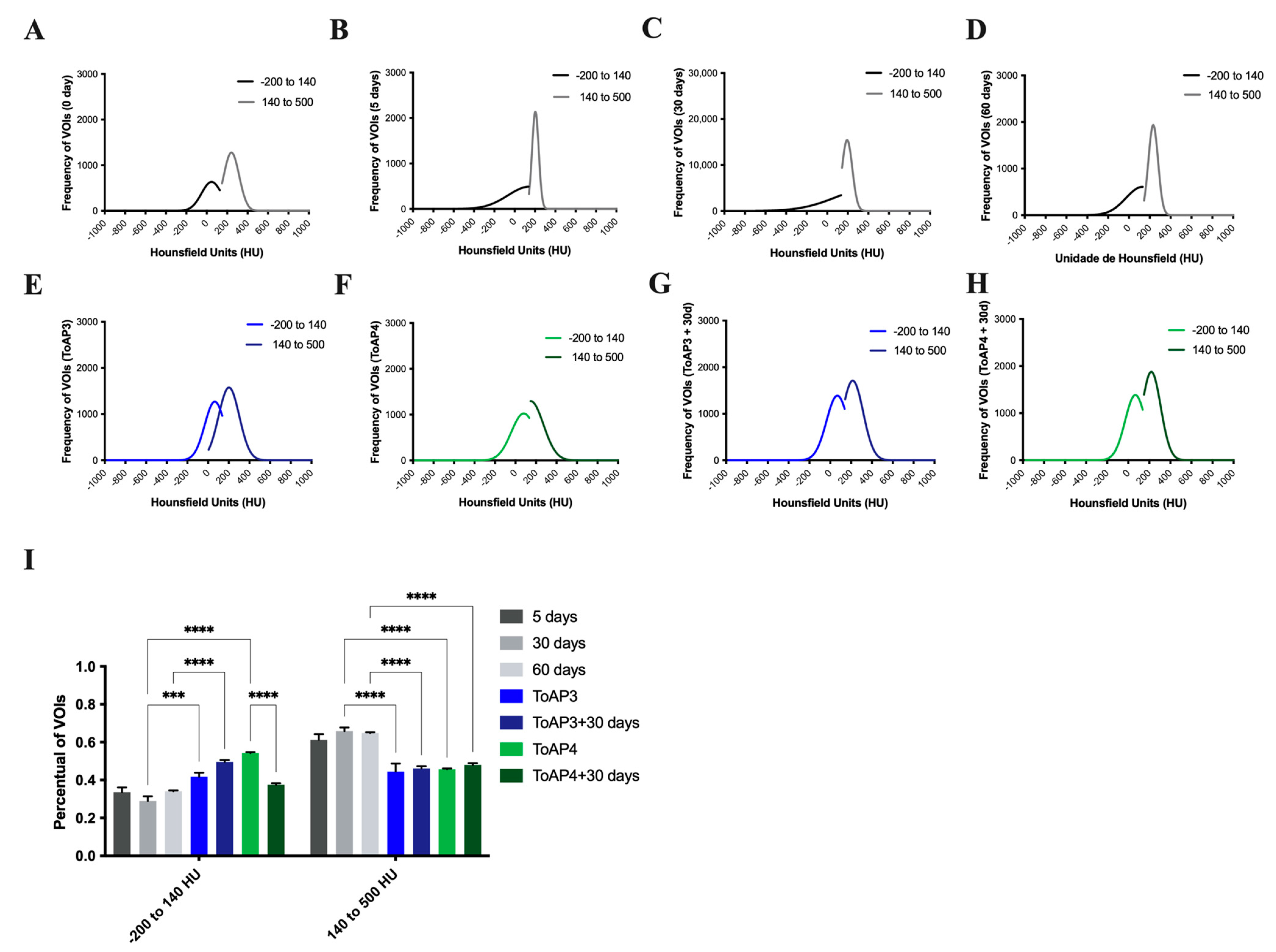
2.1. The Animal Model Strategy Showed Long-Term Inflammation with Lung Fibrosis Formation
2.2. The Treatment of Important Morphological Parameters That Contribute to Fibrosis Formation in Lung Tissue
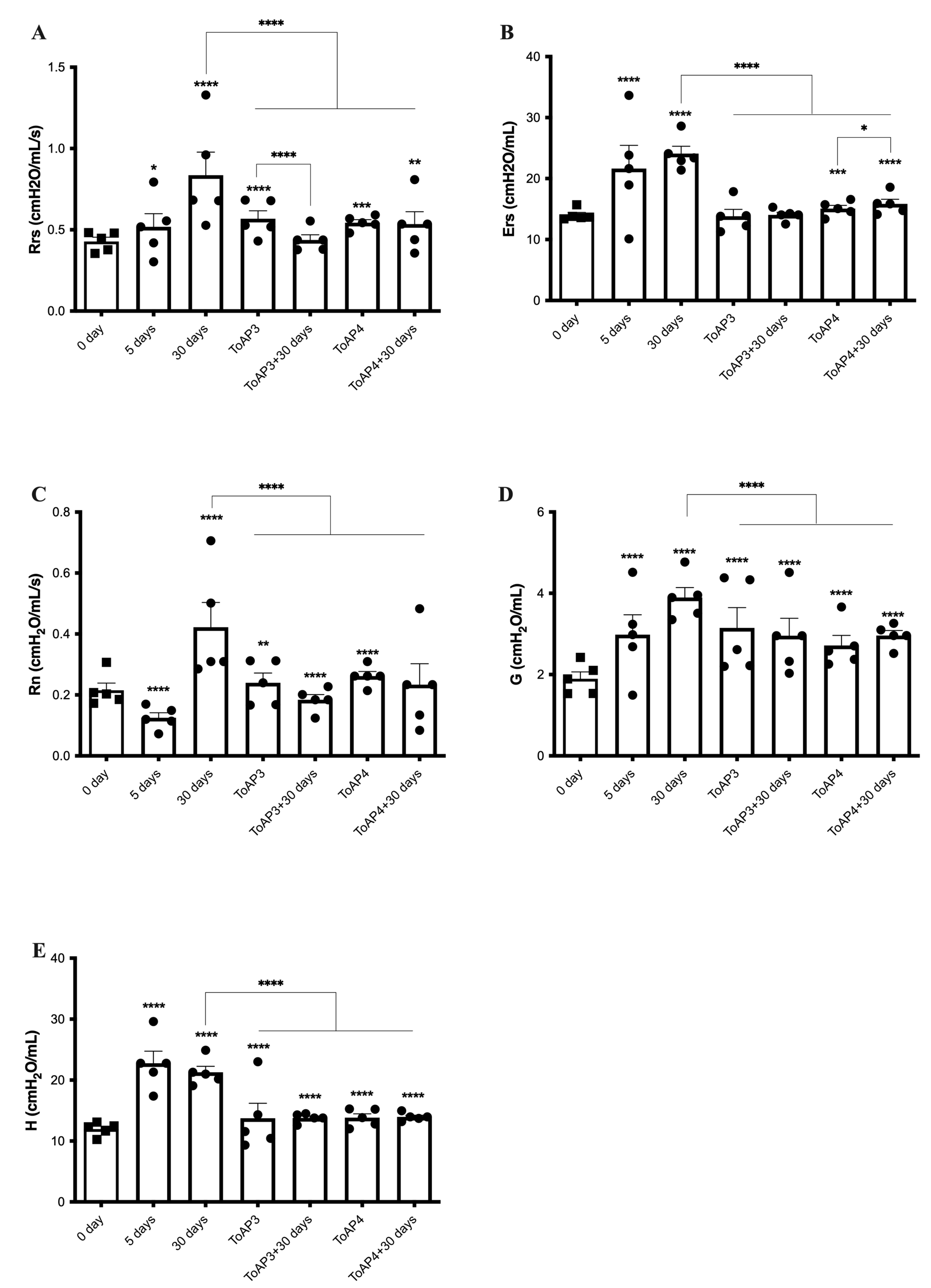
2.3. Peptide Treatments Prevent the Worsening of Pulmonary Mechanical Parameters
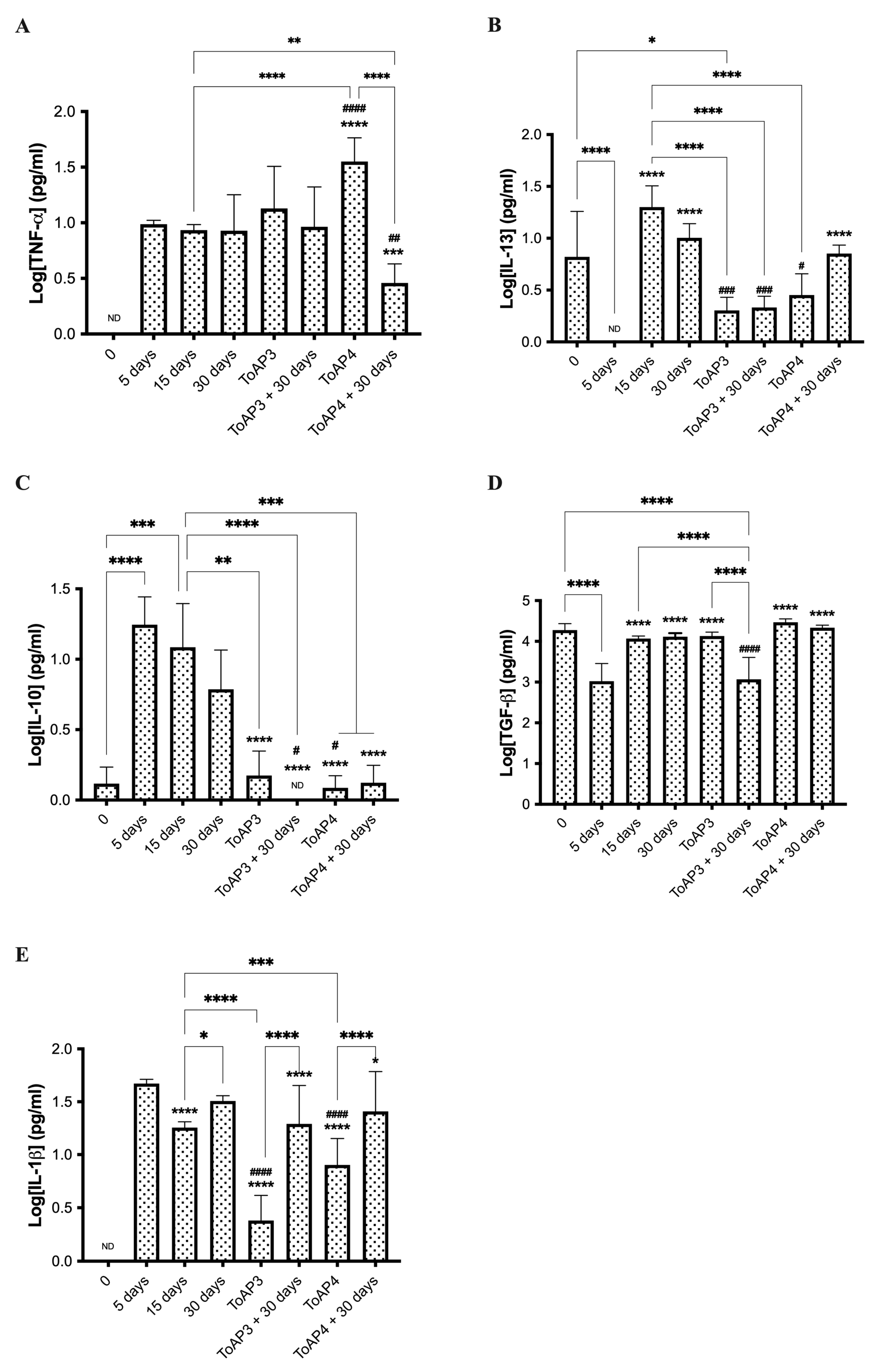
2.4. ToAp3- and ToAP4-Treatments Modulate Fibrosis Formation through Different Pathways
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Anti-Inflammatory Peptide Treatment
4.3. Morphological Evaluation
4.4. Pulmonary Ventilatory Mechanics
4.5. PCR Array
4.6. Cytokine Quantification
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, K.; Phan, S.H. Cytokines and Pulmonary Fibrosis. Biol. Signals 1996, 5, 232–239. [Google Scholar] [CrossRef]
- Wynn, T.A. Integrating mechanisms of pulmonary fibrosis. J. Exp. Med. 2011, 208, 1339–1350. [Google Scholar] [CrossRef] [Green Version]
- Raghu, G.; Collard, H.R.; Egan, J.J.; Martinez, F.J.; Behr, J.; Brown, K.K.; Colby, T.V.; Cordier, J.-F.; Flaherty, K.R.; Lasky, J.A.; et al. An Official ATS/ERS/JRS/ALAT Statement: Idiopathic pulmonary fibrosis: Evidence-based guidelines for diagnosis and management. Am. J. Respir. Crit. Care Med. 2011, 183, 788–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collard, H.R. Improving Survival in Idiopathic Pulmonary Fibrosis: The Race Has Just Begun. Chest 2017, 151, 527–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottin, V.; Schmidt, A.; Catella, L.; Porte, F.; Fernandez-Montoya, C.; Le Lay, K.; Bernard, S. Burden of idiopathic pulmonary fibrosis progression: A 5-year longitudinal follow-up study. PLoS ONE 2017, 12, e0166462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mooney, J.J.; Raimundo, K.; Chang, E.; Broder, M.S. Hospital cost and length of stay in idiopathic pulmonary fibrosis. J. Med. Econ. 2017, 20, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.T.; Spiteri, M.A. Growth factors in idiopathic pulmonary fibrosis: Relative roles. Respir. Res. 2002, 3, 13. [Google Scholar] [CrossRef] [Green Version]
- Aiello, M.; Bertorelli, G.; Bocchino, M.; Chetta, A.; Fiore-Donati, A.; Fois, A.; Marinari, S.; Oggionni, T.; Polla, B.; Rosi, E.; et al. The earlier, the better: Impact of early diagnosis on clinical outcome in idiopathic pulmonary fibrosis. Pulm. Pharmacol. Ther. 2017, 44, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Inayama, M.; Nishioka, Y.; Azuma, M.; Muto, S.; Aono, Y.; Makino, H.; Tani, K.; Uehara, H.; Izumi, K.; Itai, A.; et al. A novel IκB kinase-β inhibitor ameliorates bleomycin-induced pulmonary fibrosis in mice. Am. J. Respir. Crit. Care Med. 2006, 173, 1016–1022. [Google Scholar] [CrossRef]
- Wilson, M.S.; Wynn, T.A. Pulmonary fibrosis: Pathogenesis, etiology and regulation. Mucosal Immunol. 2009, 2, 103–121. [Google Scholar] [CrossRef] [Green Version]
- Vilcek, J.; Palombella, V.; Henriksen-DeStefano, D.; Swenson, C.; Feinman, R.; Hirai, M.; Tsujimoto, M. Fibroblast Growth Enhancing Activity of to Necrosis Factor and Its Relationship other Growth Factors. J. Exp. Med. 1986, 163, 632–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tashiro, J.; Rubio, G.A.; Limper, A.H.; Williams, K.; Elliot, S.J.; Ninou, I.; Aidinis, V.; Tzouvelekis, A.; Glassberg, K. Exploring Animal Models That Resemble Idiopathic Pulmonary Fibrosis. Front. Med. 2017, 4, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, O.; Winkler, J.; Minasyan, M.; Herzog, E.L. The Role of Immune and Inflammatory Cells in Idiopathic Pulmonary Fibrosis. Front. Med. 2018, 5, 43. [Google Scholar] [CrossRef]
- Willis, B.C.; Liebler, J.M.; Luby-Phelps, K.; Nicholson, A.G.; Crandall, E.D.; du Bois, R.M.; Borok, Z. Induction of Epithelial-Mesenchymal Transition in Alveolar Epithelial Cells by Transforming Growth Factor-β1. Am. J. Pathol. 2005, 166, 1321–1332. [Google Scholar] [CrossRef]
- Belperio, J.A.; Dy, M.; Burdick, M.D.; Xue, Y.Y.; Li, K.; Elias, J.A.; Keane, M.P. Interaction of IL-13 and C10 in the pathogenesis of bleomycin-induced pulmonary fibrosis. Am. J. Respir. Cell. Mol. Biol. 2002, 27, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Borie, R.; Justet, A.; Beltramo, G.; Manali, E.D.; Pradre, P.; Spagnolo, P.; Crestani, B. Pharmacological management of IPF. Respirology 2016, 21, 615–625. [Google Scholar]
- Sköld, C.M.; Bendstrup, E.; Myllärniemi, M.; Gudmundsson, G.; Sjåheim, T.; Hilberg, O.; Altraja, A.; Kaarteenaho, R.; Ferrara, G. Treatment of idiopathic pulmonary fibrosis: A position paper from a Nordic expert group. J. Intern. Med. 2017, 281, 149–166. [Google Scholar] [CrossRef]
- Roth, G.J.; Binder, R.; Colbatzky, F.; Dallinger, C.; Schlenker-Herceg, R.; Hilberg, F.; Wollin, S.; Kaiser, R. Nintedanib: From discovery to the clinic. J. Med. Chem. 2015, 58, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Betensley, A.; Sharif, R.; Karamichos, D. A Systematic Review of the Role of Dysfunctional Wound Healing in the Pathogenesis and Treatment of Idiopathic Pulmonary Fibrosis. J. Clin. Med. 2016, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Maher, T.M.; Strek, M.E. Antifibrotic therapy for idiopathic pulmonary fibrosis: Time to treat. Respir. Res. 2019, 20, 205. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Heiden JAVander Uttarwar, S.; Xi, Y.; Diaye, E.N.; Lacanna, R.; Caplazi, P.; Gierke, S.; Moffat, J.; Wolters, P.; Ding, N. Inhibition of MRTF activation as a clinically achievable antifibrotic mechanism for pirfenidone. Eur. Respir. J. 2023, 61, 2200604. [Google Scholar] [CrossRef] [PubMed]
- Jatene, F.B.; Fernandes, P.M.P.; De Medeiros, I.L. Transplante pulmonar—Lung transplantation. Rev. Med. 2009, 88, 111–122. [Google Scholar]
- Veloso Júnior, P.H.H.; Simon, K.S.; de Castro, R.J.A.; Coelho, L.C.; Erazo, F.A.H.; de Souza, A.C.B.; das Neves, R.C.; Lozano, V.F.; Schwartz, E.F.; Tavares, A.H.; et al. Peptides ToAP3 and ToAP4 decrease release of inflammatory cytokines through TLR-4 blocking. Biomed. Pharmacother. 2019, 118, 109152. [Google Scholar] [CrossRef] [PubMed]
- Guilhelmelli, F.; Vilela, N.; Smidt, K.S.; De Oliveira, M.A.; da Cunha Morales Álvares, A.; Rigonatto, M.C.; Costa, P.H.S.; Tavares, A.H.; Freitas, S.M.; Nicola, A.M.; et al. Activity of scorpion venom-derived antifungal peptides against planktonic cells of Candida spp. and Cryptococcus neoformans and Candida albicans biofilms. Front. Microbiol. 2016, 7, 1844. [Google Scholar] [CrossRef] [Green Version]
- Akahori, D.; Inoue, N.U.Y.; Yasui, H.; Hozumi, H.; Suzuki, Y.; Karayama, M.; Furuhashi, K.; Enomoto, N.; Tomoyuki Fujisawa, T.; Suda, T. Effect of Hypoxia on Pulmonary Endothelial Cells from Bleomycin-Induced Pulmonary Fibrosis Model Mice. Int. J. Mol. Sci. 2022, 23, 8996. [Google Scholar] [CrossRef]
- Principi, L.; Ferrini, E.; Ciccimarra, R.; Pagani, L.; Chinello, C.; Previtali, P.; Smith, A.; Villetti, G.; Zoboli, M.; Ravanetti, F.; et al. Proteomic Fingerprint of Lung Fibrosis Progression and Response to Therapy in Bleomycin-Induced Mouse Model. Int. J. Mol. Sci. 2023, 24, 4410. [Google Scholar] [CrossRef]
- Phan, S.; Kunkel, S. Lung cytokine production in bleomycin-induced pulmonary fibrosis. Exp. Lung Res. 1992, 43, 29–43. [Google Scholar] [CrossRef]
- Cook, D.N.; Brass, D.M.; Schwartz, D.A. Perspective A Matrix for New Ideas in Pulmonary Fibrosis. Am. J. Respir. Cell. Mol. Biol. 2002, 27, 122–124. [Google Scholar] [CrossRef]
- Marcos, C.M.; de Oliveira, H.C.; Assato, P.A.; de Andrade, C.R.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Paracoccidioides brasiliensis 14-3-3 protein is important for virulence in a murine model. Med. Mycol. 2019, 57, 900–904. [Google Scholar] [CrossRef]
- Rodriguez-Echeverri, C.; Puerta-Arias, J.D.; González, Á. Paracoccidioides brasiliensis activates mesenchymal stem cells through TLR2, TLR4, and Dectin-1. Med. Mycol. 2021, 59, 149–157. [Google Scholar] [CrossRef]
- Bueter, C.L.; Deepe, G.S., Jr. Aeroallergens Exacerbate Histoplasma capsulatum Infection. J. Immunol. 2018, 201, 3352–3361. [Google Scholar] [CrossRef] [PubMed]
- Defaverp, J.; Graybill, J.R. Immunohistopathology of murine pulmonary histoplasmosis during normal and hypersensitive conditions. Am. Rev. Respir. Dis. 1991, 144, 1366–1372. [Google Scholar] [CrossRef]
- Suzuki, A.; Kondoh, Y. The clinical impact of major comorbidities on idiopathic pulmonary fibrosis. Respir. Investig. 2017, 55, 94–103. [Google Scholar] [CrossRef]
- Koo, J.-W.; Myong, J.-P.; Yoon, H.-K.; Rhee, C.K.; Kim, Y.; Kim, J.S.; Jo, B.S.; Cho, Y.; Byun, J.; Choi, M. Occupational exposure and idiopathic pulmonary fibrosis: A multicentre case-control study in Korea. Int. J. Tuberc. Lung Dis. 2017, 21, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Harari, S.; Madotto, F.; Conti, S.; Cesana, G. prevalence of idiopathic pulmonary fibrosis in the USA and Canada. Eur. Respir. J. 2017, 49, 1601504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galdino de Souza, D.; Santos, D.S.; Simon, K.S.; Morais, J.A.V.; Coelho, L.C.; Pacheco, T.J.A.; Azevedo, R.B.; Bocca, A.L.; Melo-Silva, C.A.; Longo, J.P.F. Fish Oil Nanoemulsion Supplementation Attenuates Bleomycin-Induced Pulmonary Fibrosis BALB/c Mice. Nanomaterials 2022, 12, 1683. [Google Scholar] [CrossRef]
- Shen, A.S.; Haslett, C.; Feldsien, D.C.; Henson, P.M.; Cherniack, R.M. The Intensity of Chronic Lung Inflammation and Fibrosis after Bleomycin Is Directly Related to the Severity of Acute Injury. Am. Rev. Respir Dis. 1988, 137, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Piguet, P.F.; Vesin, C. Pulmonary platelet trapping induced by bleomycin: Correlation with fibrosis and involvement of the beta 2 integrins. Int. J. Exp. Pathol. 1994, 75, 321–328. [Google Scholar]
- Bringardner, B.D.; Baran, C.P.; Eubank, T.D.; Marsh, B. The Role of Inflammation in the Pathogenesis of Idiopathic Pulmonary Fibrosis. Antioxid. Redox Signal. 2008, 10, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Dolhnikoff, M.; Mauad, T.; Ludwig, M.S. Extracellular matrix and oscillatory mechanics of rat lung parenchyma in bleomycin-induced fibrosis. Am. J. Respir. Crit. Care Med. 1999, 160, 1750–1757. [Google Scholar] [CrossRef] [Green Version]
- Lutz, D.; Gazdhar, A.; Lopez-Rodriguez, E.; Ruppert, C.; Mahavadi, P.; Günther, A.; Klepetko, W.; Bates, J.; Smith, B.; Geiser, T.; et al. Alveolar derecruitment and collapse induration a crucial mechanisms in lung injury and fibrosis. Am. J. Respir. Cell. Mol. Biol. 2015, 52, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Polosukhin, V.V.; Degryse, A.L.; Newcomb, D.C.; Jones, B.R.; Ware, L.B.; Lee, J.W.; Loyd, J.E.; Blackwell, T.S.; Lawson, W.E. Intratracheal bleomycin causes airway remodeling and airflow obstruction in mice. Exp. Lung Res. 2012, 38, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Piguet, P.F.; Ribaux, C.; Karpuz, V.; Grau, G.E. Short Communication Expression and Localization of Tumor Necrosis. Am. J. Pathol. 1993, 143, 651–655. [Google Scholar] [PubMed]
- Agostini, C.; Gurrieri, C. Chemokine/cytokine cocktail in idiopathic pulmonary fibrosis. Proc. Am. Thorac. Soc. 2006, 3, 357–363. [Google Scholar] [CrossRef]
- Duncan, M.R.; Berman, B. Differential Regulation of Collagen, Glycosaminoglycan, Fibronectin, and Collagenase Activity Production in Cultured Human Adult Dermal Fibroblasts by Interleukin l-Alpha and Beta and Tumor Necrosis Factor-Alpha and Beta. J. Investig. Dermatol. 1989, 92, 699–706. [Google Scholar] [CrossRef] [Green Version]
- Piguet, P.F.; Grau, G.E.; Vassalli, P. Subcutaneous perfusion of tumor necrosis factor induces local proliferation of fibroblasts, capillaries, and epidermal cells, or massive tissue necrosis. Am. J. Pathol. 1990, 136, 103–110. [Google Scholar]
- Sullivan, D.E.; Ferris, M.B.; Pociask, D.; Brody, A.R. Tumor necrosis factor-α induces transforming growth factor-β1 expression in lung fibroblasts through the extracellular signal-regulated kinase pathway. Am. J. Respir. Cell. Mol. Biol. 2005, 32, 342–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamitani, S.; Yamauchi, Y.; Kawasaki, S.; Takami, K.; Takizawa, H.; Nagase, T.; Kohyama, T. Simultaneous stimulation with TGF-β1 and TNF-α induces epithelial mesenchymal transition in bronchial epithelial cells. Int. Arch. Allergy Immunol. 2011, 155, 119–128. [Google Scholar] [CrossRef]
- Hou, J.; Ma, T.; Cao, H.; Chen, Y.; Wang, C.; Chen, X.; Xiang, Z.; Han, X. NF-κB signaling exacerbates bleomycin-induced pulmonary fibrosis. J. Cell. Physiol. 2018, 233, 2409–2419. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, K.; Sugishita, M.; Usui, M.; Kawabe, T.; Hashimoto, N.; Hasegawa, Y. Pulmonary infectious complications associated with anti-TNFalpha therapy (infliximab) for rheumatoid arthritis. Internal Med. 2006, 45, 685–688. [Google Scholar] [CrossRef] [Green Version]
- Raghu, G.; Brown, K.K.; Costabel, U.; Cottin, V.; du Bois, R.M.; Lasky, J.A.; Thomeer, M.; Utz, J.P.; Khandker, R.K.; McDermott, L.; et al. Treatment of idiopathic pulmonary fibrosis with etanercept: An exploratory, placebo-controlled trial. Am. J. Respir. Crit. Care Med. 2008, 178, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Thavarajah, K.; Wu, P.; Rhew, E.J.; Yeldandi, A.K.; Kamp, D.W. Pulmonary Complications of Tumor Necrosis Factor-Targeted Therapy. Respir. Med. 2009, 103, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Prasse, A.; Pechkovsky, D.V.; Toews, G.B.; Jungraithmayr, W.; Kollert, F.; Goldmann, T.; Vollmer, E.; Müller-Quernheim, J.; Zissel, G. A vicious circle of alveolar macrophages and fibroblasts perpetuates pulmonary fibrosis via CCL18. Am. J. Respir. Crit. Care Med. 2006, 173, 781–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhu, X.; Hua, Y.; Zhao, Q.; Wang, K.; Zhen, L.; Wang, G.; Lü, J.; Luo, A.; Cho, W.; et al. YY1 mediates TGF-β1-induced EMT and pro-fibrogenesis in alveolar epithelial cells. Respir. Res. 2019, 20, 249. [Google Scholar] [CrossRef] [PubMed]
- Sakai, N.; Tager, A.M. Fibrosis of two: Epithelial cell-fibroblast interactions in pulmonary fibrosis. Biochim. Biophys. Acta-Mol. Basis Dis. 2013, 1832, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Morishita, Y.; Yoshizawa, H.; Watanabe, M.; Ishibashi, K.; Muto, S.; Kusano, E.; Nagata, D. SiRNAs targeted to Smad4 prevent renal fibrosis in vivo. Sci. Rep. 2014, 4, 6464. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Wang, W.; Yuan, H.; Sun, J.; Li, L.; Wu, X.; Lou, J.; Gu, Y. Sunitinib, a Small-Molecule Kinase Inhibitor, Attenuates Bleomycin-Induced Pulmonary Fibrosis in Mice. Tohoku J. Exp. Med. 2016, 239, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, K.; Gemma, A.; Yoshimura, A.; Hosoya, Y.; Nara, M.; Hosomi, Y.; Okano, T.; Kunungi, S.; Koizumi, K.; Fukuda, Y.; et al. Reduced transcription of the Smad4 gene during pulmonary carcinogenesis in idiopathic pulmonary fibrosis. Mol. Med. Rep. 2009, 2, 73–80. [Google Scholar]
- Zhu, J.; Guo, L.; Watson, C.J.; Hu-Li, J.; Paul, W.E. Stat6 Is Necessary and Sufficient for IL-4’s Role in Th2 Differentiation and Cell Expansion. J. Immunol. 2001, 166, 7276–7281. [Google Scholar] [CrossRef]
- Walford, H.H.; Doherty, T.A. STAT6 and lung inflammation. Jak-Stat. 2013, 2, e25301. [Google Scholar] [CrossRef] [Green Version]
- Richards, C.D. Innate Immune Cytokines, Fibroblast Phenotypes, and Regulation of Extracellular Matrix in Lung. J. Interf. Cytokine Res. 2017, 37, 52–61. [Google Scholar] [CrossRef]
- DosReis, G.A.; Borges, V.M.; Zin, W.A. The central role of Fas-ligand cell signaling in inflammatory lung diseases. J. Cell. Mol. Med. 2004, 8, 285–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Paepe, M.E.; Patel, C.; Tsai, A.; Gundavarapu, S.; Mao, Q. Endoglin (CD105) up-regulation in pulmonary microvasculature of ventilated preterm infants. Am. J. Respir. Crit. Care Med. 2008, 178, 180–187. [Google Scholar] [CrossRef] [Green Version]
- Dancer, R.C.A.; Wood, A.M.; Thickett, D.R. Metalloproteinases in idiopathic pulmonary fibrosis. Eur. Respir. J. 2011, 38, 1461–1467. [Google Scholar] [CrossRef] [Green Version]
- Pechkovsky, D.V.; Prasse, A.; Kollert, F.; Engel, K.M.Y.; Dentler, J.; Luttmann, W.; Friedrich, K.; Müller-Quernheim, J.; Zissel, G. Alternatively activated alveolar macrophages in pulmonary fibrosis-mediator production and intracellular signal transduction. Clin. Immunol. 2010, 137, 89–101. [Google Scholar] [CrossRef]
- Belperio, J.A.; Dy, M.; Murray, L.; Burdick, M.D.; Xue, Y.Y.; Strieter, R.M.; Keane, M.P. The role of the Th2 CC chemokine ligand CCL17 in pulmonary fibrosis. J. Immunol. 2004, 173, 4692–4698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, L.A.; Argentieri, R.L.; Farrell, F.X.; Bracht, M.; Sheng, H.; Whitaker, B.; Beck, H.; Tsui, P.; Cochilin, K.; Evanoff, H.L.; et al. Hyper-responsiveness of IPF/UIP fibroblasts: Interplay between TGFβ1, IL-13 and CCL2. Int. J. Biochem. Cell Biol. 2008, 40, 2174–2182. [Google Scholar] [CrossRef]
- Larson-Casey, J.L.; Murthy, S.; Ryan, A.J.; Brent Carter, A. Modulation of the mevalonate pathway by AKT regulates macrophage survival and development of pulmonary fibrosis. J. Biol. Chem. 2014, 289, 36204–36219. [Google Scholar] [CrossRef] [Green Version]
- Burdick, M.D.; Murray, L.A.; Keane, M.P.; Xue, Y.Y.; Zisman, D.A.; Belperio, J.A.; Strieter, R.M. CXCL11 attenuates bleomycin-induced pulmonary fibrosis via inhibition of vascular remodeling. Am. J. Respir. Crit. Care Med. 2005, 171, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolb, M.; Margetts, P.J.; Anthony, D.C.; Pitossi, F.; Gauldie, J. Transient expression of IL-1beta induces acute lung injury and chronic repair leading to pulmonary fibrosis. J. Clin. Investig. 2001, 107, 1529–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raines, E.W.; Dower, S.K.; Ross, R. Interleukin-1 mitogenic activity for fibroblasts and smooth muscle cells is due to PDGF-AA. Science 1989, 243, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Suwabe, A.; Takahashi, K.; Yasui, S.; Arai, S.; Sendo, F. Bleomycin-stimulated hamster alveolar macrophages release interleukin-1. Am. J. Pathol. 1988, 132, 512–520. [Google Scholar]
- Sánchez, F.; Orero, A.; Soriano, A.; Correcher, C.; Conde, P.; González, A.; Hernández, L.; Moliner, L.; Rodríguez-Alvarez, M.J.; Vidal, L.F.; et al. ALBIRA: A small animal PETSPECTCT imaging system. Med. Phys. 2013, 40, 051906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttmann, J.; Eberhard, L.; Wolff, G.; Bertschmann, W.; Zeravik, J.; Adolph, M. Maneuver-free determination of compliance and resistance in ventilated ARDS patients. Chest 1992, 102, 1235–1242. [Google Scholar] [CrossRef] [Green Version]
- Hantos, Z.; Daroczy, B.; Suki, B.; Nagy, S.; Fredberg, J.J. Input impedance and peripheral inhomogeneity of dog lungs. J. Appl. Physiol. 1992, 72, 168–178. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Description | 15th Day | 30th Day | ||
|---|---|---|---|---|---|
| Fold Change | p Value | Fold Change | p Value | ||
| Eng | Endoglin | 0.47 | 0.349698 | 0.37 | 0.038711 |
| Fasl | Fas ligand (TNF superfamily, member 6) | 0.65 | 0.352122 | 6.68 | 0.039064 |
| Itgb6 | Integrin beta 6 | 3.06 | 0.014495 | 2.44 | 0.019880 |
| Smad4 | Homolog of MAD 4 (Drosophila) | 0.48 | 0.005721 | 0.62 | 0.027655 |
| Stat6 | Transducer and activator of transcription 6 | 0.30 | 0.033396 | 0.54 | 0.078273 |
| Mmp13 | Matrix metalloperoxidase 13 | 6.21 | 0.149880 | 3.09 | 0.029585 |
| Symbol | Description | ToAP3 Treatment | ToAP4 Treatment | ||
|---|---|---|---|---|---|
| Fold Change | p Value | Fold Change | p Value | ||
| Akt1 | Proto-oncogene Viral de Timoma 1 | 0.34 | 0.043212 | 0.83 | 0.570278 |
| Ccl11 | Chemokine (motivo c-c) ligante 11 | 3.18 | 0.021522 | 4.41 | 0.323722 |
| Ccr2 | Chemokine RECEPTOR (motive c-c) 2 | 1.18 | 0.593361 | 0.14 | 0.043364 |
| Fasl | Fas ligand (TNF superfamily, member 6) | 4.30 | 0.006065 | 0.20 | 0.060712 |
| Il1a | Interleukin 1 alpha | 6.53 | 0.000911 | 3.19 | 0.239425 |
| Il1b | Interleukin 1 beta | 2.27 | 0.046645 | 0.62 | 0.750740 |
| Itgav | Integrin alpha V | 0.46 | 0.045766 | 1.79 | 0.375390 |
| Itgb6 | Integrin beta 6 | 0.21 | 0.005560 | 0.83 | 0.678812 |
| Mmp14 | Matrix metallopeptidase 14 | 0.18 | 0.015702 | 0.46 | 0.278868 |
| Mmp2 | Matrix metallopeptidase 2 | 0.39 | 0.043679 | 1.86 | 0.365113 |
| Pdgfa | Platelet-derived growth factor, alpha | 0.25 | 0.020553 | 1.18 | 0.536549 |
| Serpine1 | Serine (or cysteine) peptidase inhibitor, clade E, member 1 | 0.40 | 0.026483 | 0.24 | 0.000152 |
| Serpinh1 | Serine (or cysteine) peptidase inhibitor, clade H, member 1 | 0.21 | 0.025890 | 1.59 | 0.358584 |
| Smad4 | SMAD family member 4 | 0.92 | 0.778855 | 0.50 | 0.035041 |
| Tgfb1 | Transforming growth factor, beta 1 | 0.28 | 0.029165 | 2.23 | 0.338805 |
| Tgfb2 | Transforming growth factor, beta 2 | 0.47 | 0.019035 | 1.74 | 0.355887 |
| Tgfb3 | Transforming growth factor, beta 3 | 0.37 | 0.011840 | 0.71 | 0.155044 |
| Tgfbr1 | Transforming growth factor, beta receptor I | 0.44 | 0.035557 | 0.71 | 0.142083 |
| Tgfbr2 | Transforming growth factor, beta receptor 2 | 0.45 | 0.000265 | 1.34 | 0.038078 |
| Smad6 | SMAD family member 6 | 0.22 | 0.025221 | 1.20 | 0.562480 |
| Timp2 | Tissue inhibitor of metalloproteinase 2 | 0.45 | 0.003830 | 2.19 | 0.310287 |
| Genes’ Group | Name of Genes |
|---|---|
| Profibrotic | Acta2 (α-SMA), Agt, Ccl11 (Eotaxin), Ccl12 (MCP-5, Scya12), Ccl3 (Mip-1a), Ccn2, Grem1, Il13, Il13ra2, Il4, Il5, Snai1 (Snail). |
| Antifibrotic | Bmp7, Hgf, Ifng, Il10, Il13ra2 |
| Extracellular Matrix (ECM) & Cell Adhesion Molecules | Col1a2, Col3a1. Mmp13, Mmp14, Mmp1a, Mmp2, Mmp3, Mmp8, Mmp9, Plat (tPA), Plau(uPA), PlgSerpina1a, Serpine1 (Pai-1), Serpinh1 (Hsp47), Timp1, Timp2, Timp3, Timp4 Itga1, Itga2, Itga3, Itgav, Itgb1, Itgb3, Itgb5, Itgb6, Itgb8 |
| Inflammatory Cytokines & Chemokines | Ccl11 (Eotaxin), Ccl12 (MCP-5, Scya12), Ccl3 (Mip1a), Ccr2, Cxcr4, Ifng, Il10, Il13, Il13ra2, Il1a, Il1b, Il4, Il5, Ilk, Tnf |
| Growth Factors | Agt, Ccn2, Edn1, Egf, Hgf, Pdgfa, Pdgfb, Vegfa |
| Signal Transduction | TGFβ Superfamily Members-Bmp7, Cav1, Dcn, Eng (Evi1), Grem1, Inhbe, Ltbp1, Smad2, Smad3, Smad4, Smad6, Smad7, Tgfb1, Tgfb2, Tgfb3, Tgfbr1 (ALK5), Tgfbr2, Tgif1, Thbs1 (TSP-1), Thbs2. Transcription Factor-Cebpb, Jun, Myc, Nfkb1, Sp1, Stat1, Stat6 |
| Epithelial-to-Mesenchymal Transition (EMT) | Akt1, Bmp7, Col1a2, Col3a1, Itgav, Itgb1, Mmp2, Mmp3, Mmp9, Serpine1 (Pai-1), Smad2, Snai1 (Snail), Tgfb1, Tgfb2, Tgfb3, Timp1 |
| Other Fibrosis Genes | Bcl2, Fasl |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simon, K.S.; Coelho, L.C.; Veloso, P.H.d.H., Jr.; Melo-Silva, C.A.; Morais, J.A.V.; Longo, J.P.F.; Figueiredo, F.; Viana, L.; Silva Pereira, I.; Amado, V.M.; et al. Innovative Pre-Clinical Data Using Peptides to Intervene in the Evolution of Pulmonary Fibrosis. Int. J. Mol. Sci. 2023, 24, 11049. https://doi.org/10.3390/ijms241311049
Simon KS, Coelho LC, Veloso PHdH Jr., Melo-Silva CA, Morais JAV, Longo JPF, Figueiredo F, Viana L, Silva Pereira I, Amado VM, et al. Innovative Pre-Clinical Data Using Peptides to Intervene in the Evolution of Pulmonary Fibrosis. International Journal of Molecular Sciences. 2023; 24(13):11049. https://doi.org/10.3390/ijms241311049
Chicago/Turabian StyleSimon, Karina Smidt, Luísa Coutinho Coelho, Paulo Henrique de Holanda Veloso, Jr., Cesar Augusto Melo-Silva, José Athayde Vasconcelos Morais, João Paulo Figueiró Longo, Florencio Figueiredo, Leonora Viana, Ildinete Silva Pereira, Veronica Moreira Amado, and et al. 2023. "Innovative Pre-Clinical Data Using Peptides to Intervene in the Evolution of Pulmonary Fibrosis" International Journal of Molecular Sciences 24, no. 13: 11049. https://doi.org/10.3390/ijms241311049