
Age-Dependent Changes in the Relationships between Traits Associated with the Pathogenesis of Stress-Sensitive Hypertension in ISIAH Rats

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Age-Dependent Differences in Phenotypic Traits between Two Groups of F2(ISIAHxWAG) Hybrid Rats of Different Ages
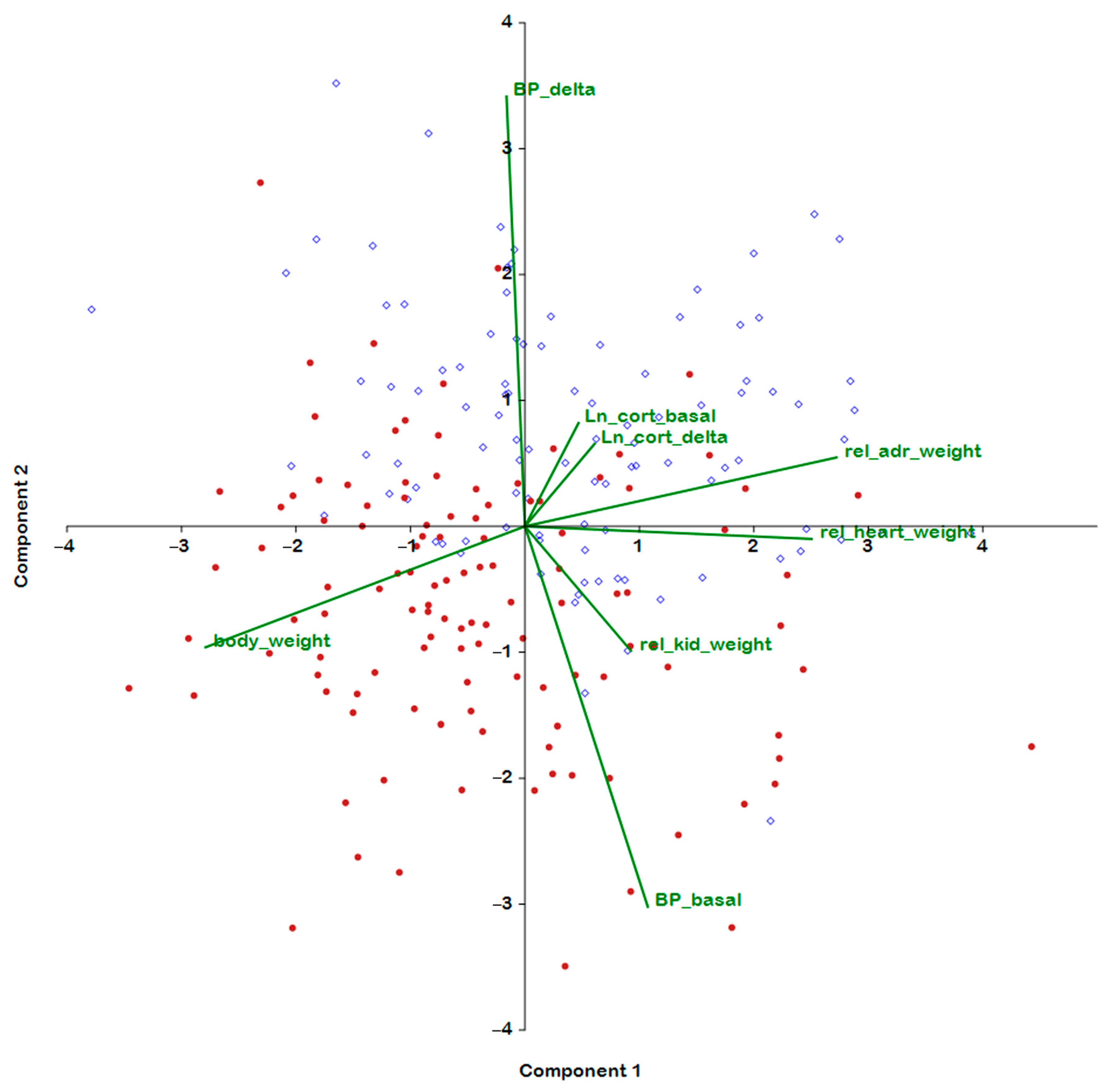
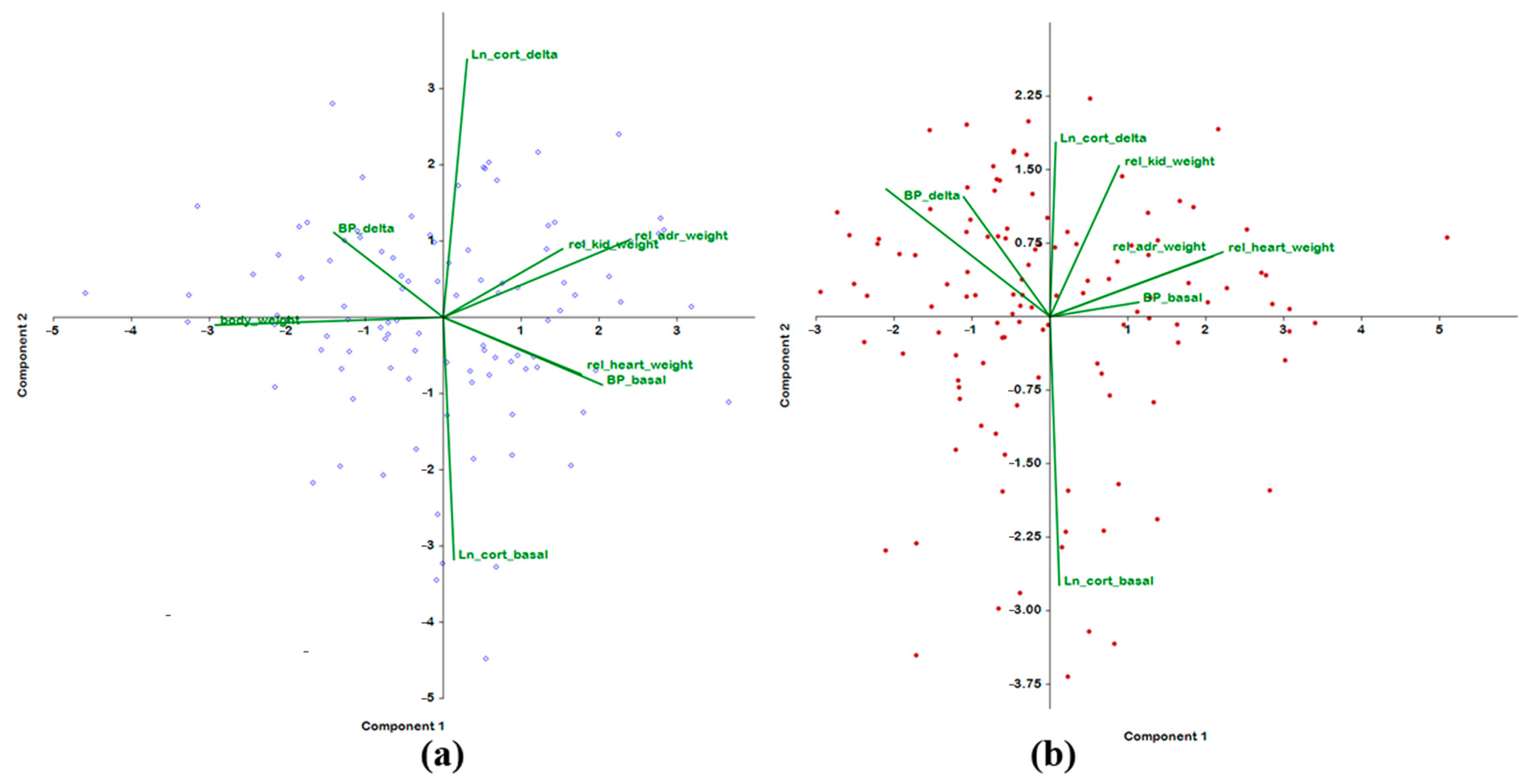
2.2. Analysis of Associations of BP Traits and of Weight Parameters with Plasma Corticosterone Concentration
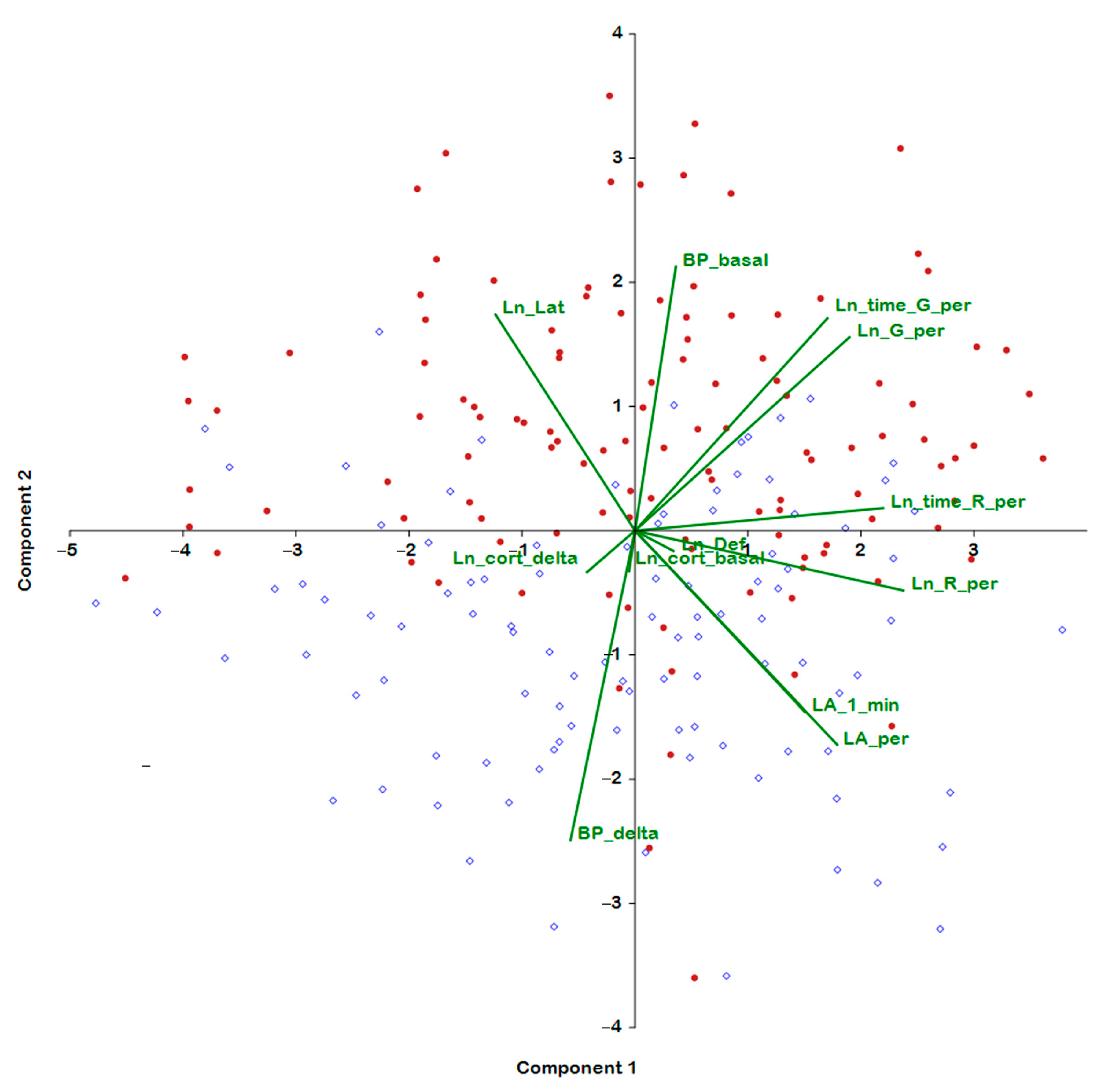
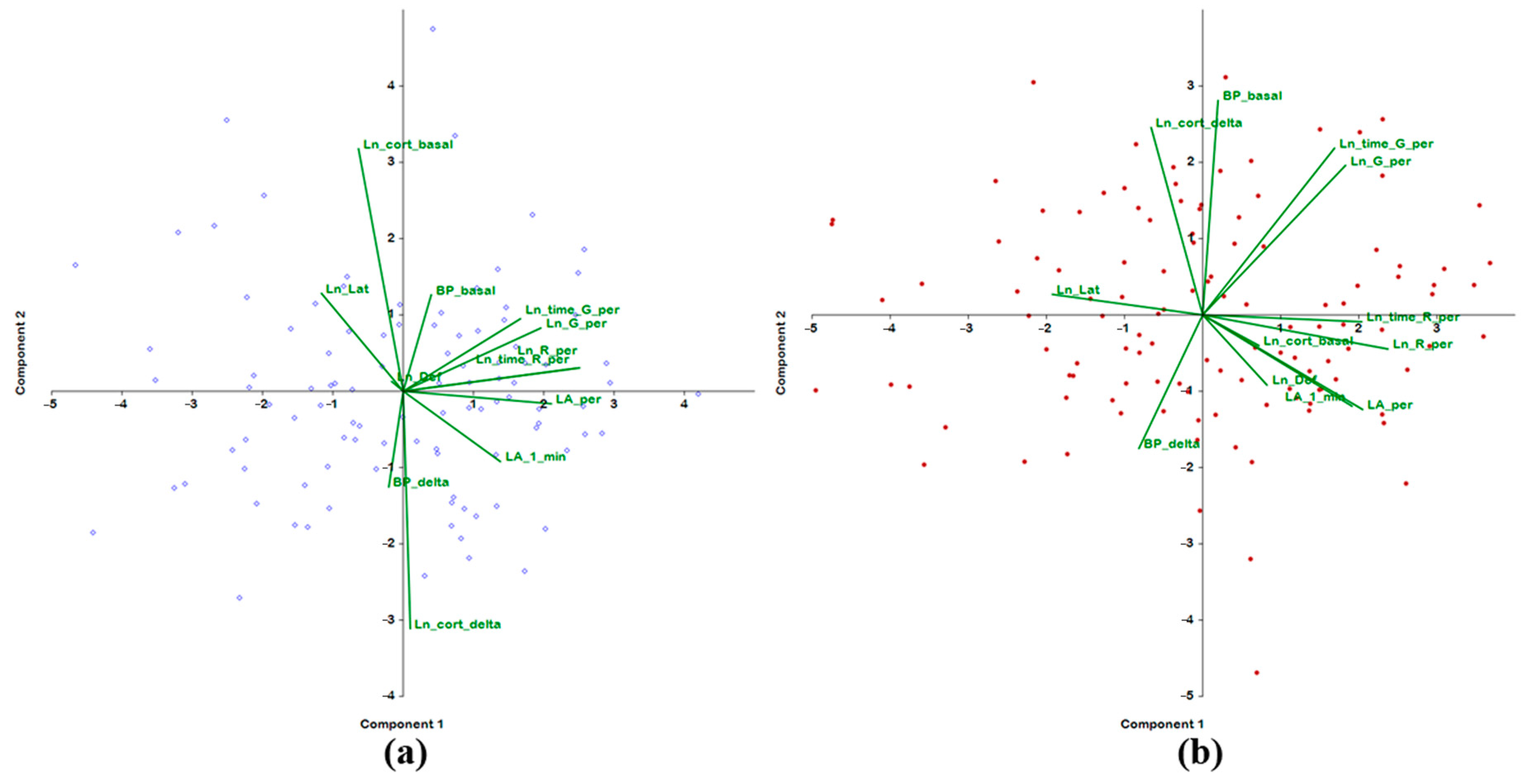
2.3. Analysis of the Associations of Behavior in the Open Field Test with BP and Hormonal Parameters
3. Discussion
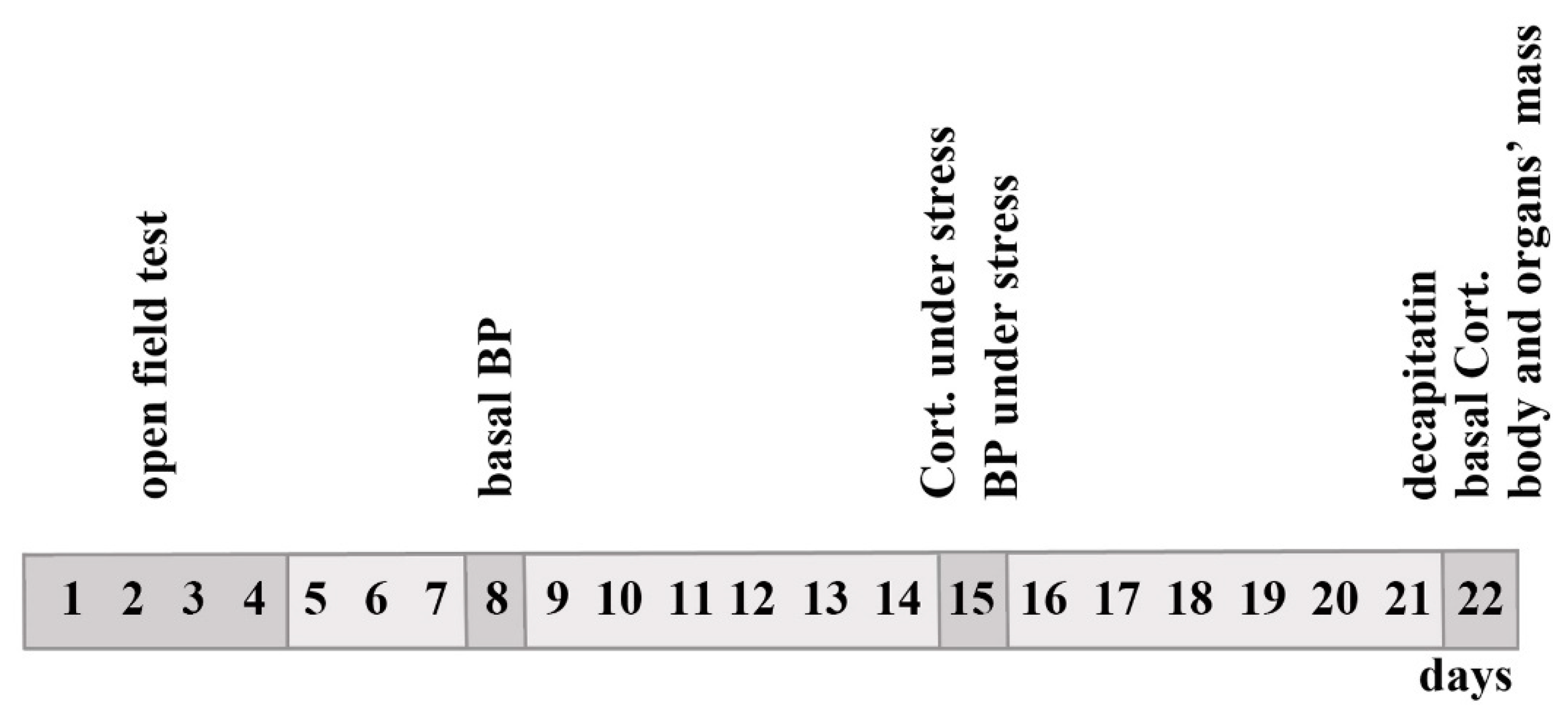
4. Materials and Methods
4.1. Animals
4.2. Measurements of BP and Other Traits
4.3. Parameters of Rat Behavior
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bozkurt, B.; Aguilar, D.; Deswal, A.; Dunbar, S.B.; Francis, G.S.; Horwich, T.; Jessup, M.; Kosiborod, M.; Pritchett, A.M.; Ramasubbu, K.; et al. Contributory Risk and Management of Comorbidities of Hypertension, Obesity, Diabetes Mellitus, Hyperlipidemia, and Metabolic Syndrome in Chronic Heart Failure: A Scientific Statement from the American Heart Association. Circulation 2016, 134, e535–e578. [Google Scholar] [CrossRef] [PubMed]
- Hamrahian, S.M.; Falkner, B. Hypertension in Chronic Kidney Disease. Adv. Exp. Med. Biol. 2017, 956, 307–325. [Google Scholar] [CrossRef] [PubMed]
- Teck, J. Diabetes-Associated Comorbidities. Prim. Care 2022, 49, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Di Palo, K.E.; Barone, N.J. Hypertension and Heart Failure: Prevention, Targets, and Treatment. Heart Fail Clin. 2020, 16, 99–106. [Google Scholar] [CrossRef]
- Omote, K.; Verbrugge, F.H.; Borlaug, B.A. Heart Failure with Preserved Ejection Fraction: Mechanisms and Treatment Strategies. Annu. Rev. Med. 2022, 73, 321–337. [Google Scholar] [CrossRef]
- Flack, J.M.; Adekola, B. Blood pressure and the new ACC/AHA hypertension guidelines. Trends Cardiovasc. Med. 2020, 30, 160–164. [Google Scholar] [CrossRef]
- Ding, P.; Pan, Y.; Wang, Q.; Xu, R. Prediction and evaluation of combination pharmacotherapy using natural language processing, machine learning and patient electronic health records. J. Biomed. Inform. 2022, 133, 104164. [Google Scholar] [CrossRef]
- Johnson, R.J.; Rodriguez-Iturbe, B.; Kang, D.H.; Feig, D.I.; Herrera-Acosta, J. A unifying pathway for essential hypertension. Am. J. Hypertens. 2005, 18, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Feldstein, C.; Julius, S. Establishing targets for hypertension control in patients with comorbidities. Curr. Hypertens. Rep. 2010, 12, 465–473. [Google Scholar] [CrossRef]
- Magvanjav, O.; Cooper-DeHoff, R.M.; McDonough, C.W.; Gong, Y.; Segal, M.S.; Hogan, W.R.; Johnson, J.A. Antihypertensive therapy prescribing patterns and correlates of blood pressure control among hypertensive patients with chronic kidney disease. J. Clin. Hypertens. 2019, 21, 91–101. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, K.I.; Cho, M.C. Current status and therapeutic considerations of hypertension in the elderly. Korean J. Intern. Med. 2019, 34, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Oliveros, E.; Patel, H.; Kyung, S.; Fugar, S.; Goldberg, A.; Madan, N.; Williams, K.A. Hypertension in older adults: Assessment, management, and challenges. Clin. Cardiol. 2020, 43, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Stawski, R.S.; Scott, S.B.; Zawadzki, M.J.; Sliwinski, M.J.; Marcusson-Clavertz, D.; Kim, J.; Lanza, S.T.; Green, P.A.; Almeida, D.M.; Smyth, J.M. Age differences in everyday stressor-related negative affect: A coordinated analysis. Psychol. Aging. 2019, 34, 91–105. [Google Scholar] [CrossRef]
- Whittaker, A.C.; Ginty, A.; Hughes, B.M.; Steptoe, A.; Lovallo, W.R. Cardiovascular Stress Reactivity and Health: Recent Questions and Future Directions. Psychosom. Med. 2021, 83, 756–766. [Google Scholar] [CrossRef]
- Markel, A.L. Development of a new strain of rats with inherited stress-induced arterial hypertension. In Genetic Hypertension; Sassard, J., Libbey, J., Eurotext, Eds.; Colloque INSERM: Paris, France, 1992; Volume 218, pp. 405–407. [Google Scholar]
- Markel, A.L.; Maslova, L.N.; Shishkina, G.T.; Bulygina, V.V.; Machanova, N.A.; Jacobson, G.S. Developmental influences on blood pressure regulation in ISIAH rats. In Development of the Hypertensive Phenotype: Basic and Clinical Studies; McCarty, R., Blizard, D.A., Chevalier, R.L., Birkenhager, W.H., Reid, J.L., Eds.; Handbook of hypertension; Elsevier: Amsterdam, The Netherlands, 1999; Volume 19, pp. 493–526. [Google Scholar]
- Markel, A.L.; Redina, O.E.; Gilinsky, M.A.; Dymshits, G.M.; Kalashnikova, E.V.; Khvorostova, Y.V.; Fedoseeva, L.A.; Jacobson, G.S. Neuroendocrine profiling in inherited stress-induced arterial hypertension rat strain with stress-sensitive arterial hypertension. J. Endocrinol. 2007, 195, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Redina, O.E.; Markel, A.L. Stress, Genes, and Hypertension. Contribution of the ISIAH Rat Strain Study. Curr. Hypertens. Rep. 2018, 20, 66. [Google Scholar] [CrossRef]
- Markel, A.L.; Galaktionov Yu, K.; Efimov, V.M. Factor analysis of rat behavior in an open field test. Neurosci. Behav. Physiol. 1989, 19, 279–286. [Google Scholar] [CrossRef]
- Efimov, V.M.; Kovaleva, V.Y.; Markel, A.L. A new approach to the study of genetic variability of complex characters. Heredity 2005, 94, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Sturman, O.; Germain, P.L.; Bohacek, J. Exploratory rearing: A context- and stress-sensitive behavior recorded in the open-field test. Stress 2018, 21, 443–452. [Google Scholar] [CrossRef]
- Johnson, E.O.; Kamilaris, T.C.; Chrousos, G.P.; Gold, P.W. Mechanisms of stress: A dynamic overview of hormonal and behavioral homeostasis. Neurosci. Biobehav. Rev. 1992, 16, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Marin, J. Age-related changes in vascular responses: A review. Mech. Ageing. Dev. 1995, 79, 71–114. [Google Scholar] [CrossRef] [PubMed]
- Hausler, A.; Girard, J.; Baumann, J.B.; Ruch, W.; Otten, U.H. Stress-induced secretion of ACTH and corticosterone during development of spontaneous hypertension in rats. Clin. Exp. Hypertens A 1983, 5, 11–19. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, K.; Docherty, J.R.; Kelly, J.G. Adrenoceptor status and cardiovascular function in ageing. J. Hypertens Suppl. 1988, 6, S59–S62. [Google Scholar]
- Yiallouris, A.; Tsioutis, C.; Agapidaki, E.; Zafeiri, M.; Agouridis, A.P.; Ntourakis, D.; Johnson, E.O. Adrenal Aging and Its Implications on Stress Responsiveness in Humans. Front Endocrinol. 2019, 10, 54. [Google Scholar] [CrossRef]
- Hamer, M.; Steptoe, A. Cortisol responses to mental stress and incident hypertension in healthy men and women. J. Clin. Endocrinol. Metab. 2012, 97, E29–E34. [Google Scholar] [CrossRef] [Green Version]
- Ramsay, D.; Lewis, M. Reactivity and regulation in cortisol and behavioral responses to stress. Child. Dev. 2003, 74, 456–464. [Google Scholar] [CrossRef]
- Carroll, D.; Ginty, A.T.; Whittaker, A.C.; Lovallo, W.R.; de Rooij, S.R. The behavioural, cognitive, and neural corollaries of blunted cardiovascular and cortisol reactions to acute psychological stress. Neurosci. Biobehav. Rev. 2017, 77, 74–86. [Google Scholar] [CrossRef]
- Lovallo, W.R.; Cohoon, A.J.; Sorocco, K.H.; Vincent, A.S.; Acheson, A.; Hodgkinson, C.A.; Goldman, D. Early-Life Adversity and Blunted Stress Reactivity as Predictors of Alcohol and Drug use in Persons with COMT (rs4680) Val158Met Genotypes. Alcohol. Clin. Exp. Res. 2019, 43, 1519–1527. [Google Scholar] [CrossRef]
- Oshchepkov, D.; Chadaeva, I.; Kozhemyakina, R.; Zolotareva, K.; Khandaev, B.; Sharypova, E.; Ponomarenko, P.; Bogomolov, A.; Klimova, N.V.; Shikhevich, S.; et al. Stress Reactivity, Susceptibility to Hypertension, and Differential Expression of Genes in Hypertensive Compared to Normotensive Patients. Int. J. Mol. Sci. 2022, 23, 2835. [Google Scholar] [CrossRef]
- Fedoseeva, L.A.; Klimov, L.O.; Ershov, N.I.; Alexandrovich, Y.V.; Efimov, V.M.; Markel, A.L.; Redina, O.E. Molecular determinants of the adrenal gland functioning related to stress-sensitive hypertension in ISIAH rats. BMC Genomics 2016, 17 (Suppl. 14), 989. [Google Scholar] [CrossRef] [Green Version]
- Fedoseeva, L.A.; Klimov, L.O.; Ershov, N.I.; Efimov, V.M.; Markel, A.L.; Orlov, Y.L.; Redina, O.E. The differences in brain stem transcriptional profiling in hypertensive ISIAH and normotensive WAG rats. BMC Genom. 2019, 20, 297. [Google Scholar] [CrossRef] [PubMed]
- Fedoseeva, L.A.; Ryazanova, M.A.; Ershov, N.I.; Markel, A.L.; Redina, O.E. Comparative transcriptional profiling of renal cortex in rats with inherited stress-induced arterial hypertension and normotensive Wistar Albino Glaxo rats. BMC Genet. 2016, 17 (Suppl. 1), 12. [Google Scholar] [CrossRef] [Green Version]
- Klimov, L.O.; Ershov, N.I.; Efimov, V.M.; Markel, A.L.; Redina, O.E. Genome-wide transcriptome analysis of hypothalamus in rats with inherited stress-induced arterial hypertension. BMC Genet. 2016, 17 (Suppl. 1), 13. [Google Scholar] [CrossRef] [Green Version]
- Ryazanova, M.A.; Fedoseeva, L.A.; Ershov, N.I.; Efimov, V.M.; Markel, A.L.; Redina, O.E. The gene-expression profile of renal medulla in ISIAH rats with inherited stress-induced arterial hypertension. BMC Genet. 2016, 17, 155–171. [Google Scholar] [CrossRef] [Green Version]
- Girotti, M.; Pace, T.W.; Gaylord, R.I.; Rubin, B.A.; Herman, J.P.; Spencer, R.L. Habituation to repeated restraint stress is associated with lack of stress-induced c-fos expression in primary sensory processing areas of the rat brain. Neuroscience 2006, 138, 1067–1081. [Google Scholar] [CrossRef]
- Mansi, J.A.; Rivest, S.; Drolet, G. Effect of immobilization stress on transcriptional activity of inducible immediate-early genes, corticotropin-releasing factor, its type 1 receptor, and enkephalin in the hypothalamus of borderline hypertensive rats. J. Neurochem. 1998, 70, 1556–1566. [Google Scholar] [CrossRef] [Green Version]
- Imaki, T.; Shibasaki, T.; Chikada, N.; Harada, S.; Naruse, M.; Demura, H. Different expression of immediate-early genes in the rat paraventricular nucleus induced by stress: Relation to corticotropin-releasing factor gene transcription. Endocr. J. 1996, 43, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Imaki, T.; Naruse, M.; Harada, S.; Chikada, N.; Nakajima, K.; Yoshimoto, T.; Demura, H. Stress-induced changes of gene expression in the paraventricular nucleus are enhanced in spontaneously hypertensive rats. J. Neuroendocrinol. 1998, 10, 635–643. [Google Scholar] [CrossRef]
- Brosschot, J.F. Ever at the ready for events that never happen. Eur. J. Psychotraumatol. 2017, 8, 1309934. [Google Scholar] [CrossRef] [Green Version]
- van den Beld, A.W.; Kaufman, J.M.; Zillikens, M.C.; Lamberts, S.W.J.; Egan, J.M.; van der Lely, A.J. The physiology of endocrine systems with ageing. Lancet Diabetes Endocrinol. 2018, 6, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Toczek, M.; Baranowska-Kuczko, M.; Grzeda, E.; Pedzinska-Betiuk, A.; Weresa, J.; Malinowska, B. Age-specific influences of chronic administration of the fatty acid amide hydrolase inhibitor URB597 on cardiovascular parameters and organ hypertrophy in DOCA-salt hypertensive rats. Pharmacol. Rep. 2016, 68, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Redina, O.E.; Smolenskaya, S.E.; Maslova, L.N.; Markel, A.L. Genetic Control of the Corticosterone Level at Rest and Under Emotional Stress in ISIAH Rats with Inherited Stress-Induced Arterial Hypertension. Clin. Exp. Hypertens. 2010, 32, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Redina, O.E.; Smolenskaya, S.E.; Maslova, L.N.; Markel, A.L. The genetic control of blood pressure and body composition in rats with stress-sensitive hypertension. Clin. Exp. Hypertens. 2013, 35, 484–495. [Google Scholar] [CrossRef]
- Redina, O.E.; Smolenskaya, S.E.; Markel, A.L. Genetic control of the behavior of ISIAH rats in the open field test. Russ. J. Genet. 2022, 58, 791–803. [Google Scholar] [CrossRef]
- Samani, N.J.; Gauguier, D.; Vincent, M.; Kaiser, M.A.; Bihoreau, M.T.; Lodwick, D.; Wallis, R.; Parent, V.; Kimber, P.; Rattray, F.; et al. Analysis of quantitative trait loci for blood pressure on rat chromosomes 2 and 13. Age-related differences in effect. Hypertension 1996, 28, 1118–1122. [Google Scholar] [CrossRef]
- Kovacs, P.; van den Brandt, J.; Kloting, I. Effects of quantitative trait loci for lipid phenotypes in the rat are influenced by age. Clin. Exp. Pharmacol. Physiol. 1998, 25, 1004–1007. [Google Scholar] [CrossRef]
- Garrett, M.R.; Dene, H.; Rapp, J.P. Time-course genetic analysis of albuminuria in Dahl salt-sensitive rats on low-salt diet. J. Am. Soc. Nephrol. 2003, 14, 1175–1187. [Google Scholar] [CrossRef] [Green Version]
- Beck, S.R.; Brown, W.M.; Williams, A.H.; Pierce, J.; Rich, S.S.; Langefeld, C.D. Age-stratified QTL genome scan analyses for anthropometric measures. BMC Genet. 2003, 4, S31. [Google Scholar] [CrossRef] [Green Version]
- Turana, Y.; Tengkawan, J.; Chia, Y.C.; Shin, J.; Chen, C.H.; Park, S.; Tsoi, K.; Buranakitjaroen, P.; Soenarta, A.A.; Siddique, S.; et al. Mental health problems and hypertension in the elderly: Review from the HOPE Asia Network. J. Clin. Hypertens. 2021, 23, 504–512. [Google Scholar] [CrossRef]
- Liang, X.; Shan, Y.; Ding, D.; Zhao, Q.; Guo, Q.; Zheng, L.; Deng, W.; Luo, J.; Tse, L.A.; Hong, Z. Hypertension and High Blood Pressure Are Associated with Dementia Among Chinese Dwelling Elderly: The Shanghai Aging Study. Front Neurol. 2018, 9, 664. [Google Scholar] [CrossRef] [Green Version]
- Tinnikov, A.A.; Bazhan, N.M. Determination of glucocorticoids in blood and adrenal gland cultures by competitive protein binding without preliminary extraction. Lab Delo 1984, 12, 709–713. [Google Scholar]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9–18. [Google Scholar]
- Schaare, H.L.; Blochl, M.; Kumral, D.; Uhlig, M.; Lemcke, L.; Valk, S.L.; Villringer, A. Associations between mental health, blood pressure and the development of hypertension. Nat. Commun. 2023, 14, 1953. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | F2(ISIAHxWAG) (3–4 Months Old) n = 101 | F2(ISIAHxWAG) (6–7 Months Old) n = 118 |
|---|---|---|
| Basal BP, mmHg | 158.6 ± 1.6 ** xxx | 164.3 ± 1.7 xx |
| BP under stress, mmHg | 189.9 ± 1.8 *** | 172.2 ± 1.8 |
| Increase in BP during stress, mmHg | 31.3 ± 2.0 *** | 7.9 ± 1.9 |
| Plasma corticosterone concentration at rest, µg/100 mL | 3.38 ± 0.35 ** xxx | 1.92 ± 0.17 xxx |
| Plasma corticosterone concentration during stress, µg/100 mL | 31.6 ± 0.53 *** | 25.5 ± 0.93 |
| Increase in plasma corticosterone concentration under stress, µg/100 mL | 28.2 ± 0.64 *** | 23.7 ± 0.95 |
| Body weight, g | 253.9 ± 3.52 *** | 297.4 ± 3.9 |
| Adrenal weight, mg | 36.0 ± 0.47 *** | 39.5 ± 0.54 |
| Adrenal weight/body weight, mg/100 g body weight | 14.4 ± 0.23 ** | 13.4 ± 0.2 |
| Kidney weight, g | 1.52 ± 0.02 *** | 1.87 ± 0.03 |
| Kidney weight/body weight, g/100 g body weight | 0.60 ± 0.006 | 0.63 ± 0.005 |
| Heart weight, g | 0.95 ± 0.016 *** | 1.09 ± 0.01 |
| Heart weight/body weight, g/100 g body weight | 0.38 ± 0.0035 | 0.37 ± 0.003 |
| Locomotor activity in the first minute of the first trial of the test, crossed squares | 51.4 ± 2.3 | 47.4 ± 1.9 |
| Locomotor activity on the periphery of the open-field area, crossed squares | 319 ± 14.9 *** | 241.3 ± 10.1 |
| Grooming at the periphery of the open-field area | 3.2 ± 0.36 *** | 5.4 ± 0.5 |
| Time of grooming at the periphery of the open-field area, s | 43.4 ±0.52 *** | 94.0 ±10.2 |
| Rearings at the periphery of the open-field area | 19.0 ± 1.2 | 19.2 ± 0.9 |
| Time of rearing at the periphery of the open-field area, s | 57.2 ± 3.8 *** | 75.7 ± 4.0 |
| Defecation | 13.0 ± 0.76 | 13.2 ± 0.6 |
| Latency period, s | 40.0 ± 2.1 *** | 84.5 ± 6.2 |
| Trait | PC 1 | PC 2 | PC 3 | PC 4 | PC 5 |
|---|---|---|---|---|---|
| BP_basal | 0.313 | −0.765 | 0.029 | −0.339 | 0.184 |
| BP_delta | −0.047 | 0.865 | 0.178 | 0.114 | 0.213 |
| Ln_cort_basal | 0.136 | 0.208 | −0.808 | −0.193 | 0.269 |
| Ln_cort_delta | 0.177 | 0.166 | 0.723 | −0.482 | 0.274 |
| body_weight | −0.813 | −0.243 | 0.216 | 0.103 | 0.188 |
| rel_adr_weight | 0.793 | 0.139 | 0.163 | −0.058 | −0.428 |
| rel_kid_weight | 0.270 | −0.250 | 0.222 | 0.800 | 0.116 |
| rel_heart_weight | 0.729 | −0.025 | −0.045 | 0.189 | 0.450 |
| age_indicator | −0.287 | −0.559 | −0.003 | 0.311 | −0.094 |
| % of variance | 25.55 | 19.33 | 16.65 | 13.58 | 8.26 |
| 3–4 Months Old | |||||
|---|---|---|---|---|---|
| Trait | PC 1 | PC 2 | PC 3 | PC 4 | PC 5 |
| BP_basal | 0.606 | −0.227 | −0.540 | −0.006 | 0.081 |
| BP_delta | −0.416 | 0.284 | 0.713 | −0.143 | −0.151 |
| Ln_cort_basal | 0.041 | −0.814 | 0.313 | −0.241 | 0.087 |
| Ln_cort_delta | 0.091 | 0.867 | −0.188 | 0.042 | −0.083 |
| body_weight | −0.869 | −0.026 | −0.126 | 0.135 | 0.104 |
| rel_adr_weight | 0.709 | 0.260 | 0.212 | −0.516 | −0.147 |
| rel_kid_weight | 0.454 | 0.230 | 0.426 | 0.256 | 0.701 |
| rel_heart_weight | 0.526 | −0.191 | 0.332 | 0.603 | −0.440 |
| % of variance | 28.64 | 21.31 | 16.07 | 9.91 | 9.51 |
| 6–7 Months Old | |||||
| Trait | PC 1 | PC 2 | PC 3 | PC 4 | PC 5 |
| BP_basal | 0.411 | 0.044 | 0.761 | −0.246 | 0.117 |
| BP_delta | −0.400 | 0.370 | −0.643 | 0.272 | 0.243 |
| Ln_cort_basal | 0.043 | −0.830 | −0.058 | 0.095 | 0.486 |
| Ln_cort_delta | 0.027 | 0.539 | 0.499 | 0.503 | 0.379 |
| body_weight | −0.758 | 0.394 | 0.288 | −0.155 | −0.030 |
| rel_adr_weight | 0.756 | 0.187 | −0.154 | 0.335 | −0.290 |
| rel_kid_weight | 0.320 | 0.466 | −0.351 | −0.618 | 0.300 |
| rel_heart_weight | 0.801 | 0.200 | −0.167 | 0.024 | 0.148 |
| % of variance | 27.77 | 19.58 | 18.78 | 11.43 | 8.12 |
| Trait | PC 1 | PC 2 | PC 3 | PC 4 | PC 5 |
|---|---|---|---|---|---|
| BP_basal | 0.131 | 0.581 | −0.376 | −0.214 | 0.539 |
| BP_delta | −0.207 | −0.680 | 0.150 | 0.428 | −0.240 |
| Ln_cort_basal | −0.019 | −0.089 | 0.594 | −0.303 | 0.316 |
| Ln_cort_delta | −0.155 | −0.091 | −0.553 | 0.629 | 0.259 |
| LA_1_min | 0.545 | −0.398 | 0.110 | −0.031 | 0.308 |
| LA_per | 0.649 | −0.470 | −0.239 | −0.207 | −0.003 |
| Ln_G_per | 0.689 | 0.426 | 0.280 | 0.439 | −0.047 |
| Ln_R_per | 0.862 | −0.132 | −0.197 | −0.179 | −0.207 |
| Ln_time_G_per | 0.618 | 0.467 | 0.320 | 0.468 | −0.040 |
| Ln_time_R_per | 0.797 | 0.050 | −0.229 | −0.192 | −0.344 |
| Ln_Def | 0.124 | −0.045 | 0.435 | −0.042 | 0.127 |
| Ln_Lat | −0.448 | 0.475 | −0.020 | −0.179 | −0.566 |
| age_indicator | 0.086 | 0.577 | 0.039 | −0.181 | −0.280 |
| % of variance | 27.11 | 15.36 | 11.28 | 10.62 | 9.26 |
| 3–4 Months Old | |||||
|---|---|---|---|---|---|
| Trait | PC 1 | PC 2 | PC 3 | PC 4 | PC 5 |
| BP_basal | 0.139 | 0.318 | −0.632 | 0.439 | 0.182 |
| BP_delta | −0.071 | −0.316 | 0.719 | −0.330 | 0.013 |
| Ln_cort_basal | −0.221 | 0.801 | 0.153 | −0.201 | 0.101 |
| Ln_cort_delta | 0.035 | −0.785 | −0.071 | 0.397 | −0.127 |
| LA_1_min | 0.480 | −0.232 | 0.059 | −0.207 | 0.128 |
| LA_per | 0.733 | −0.041 | −0.139 | −0.411 | −0.031 |
| Ln_G_per | 0.681 | 0.209 | 0.365 | 0.515 | −0.168 |
| Ln_R_per | 0.873 | 0.077 | −0.117 | −0.249 | 0.011 |
| Ln_time_G_per | 0.580 | 0.239 | 0.495 | 0.512 | −0.144 |
| Ln_time_R_per | 0.852 | 0.076 | −0.092 | −0.242 | −0.007 |
| Ln_Def | −0.057 | 0.032 | 0.388 | 0.149 | 0.790 |
| Ln_Lat | −0.404 | 0.323 | 0.208 | −0.106 | −0.502 |
| % of variance | 27.48 | 14.45 | 13.02 | 11.61 | 8.35 |
| 6–7 Months Old | |||||
| Trait | PC 1 | PC 2 | PC 3 | PC 4 | PC 5 |
| BP_basal | 0.069 | 0.676 | −0.534 | −0.207 | −0.081 |
| BP_delta | −0.288 | −0.421 | 0.548 | 0.474 | −0.058 |
| Ln_cort_basal | 0.254 | −0.097 | 0.216 | −0.628 | −0.465 |
| Ln_cort_delta | −0.233 | 0.591 | −0.063 | 0.355 | 0.063 |
| LA_1_min | 0.675 | −0.288 | 0.025 | −0.257 | −0.082 |
| LA_per | 0.723 | −0.298 | −0.368 | 0.181 | −0.007 |
| Ln_G_per | 0.645 | 0.471 | 0.516 | 0.020 | 0.079 |
| Ln_R_per | 0.837 | −0.108 | −0.179 | 0.340 | −0.100 |
| Ln_time_G_per | 0.596 | 0.526 | 0.521 | 0.016 | 0.101 |
| Ln_time_R_per | 0.718 | −0.022 | −0.213 | 0.469 | −0.099 |
| Ln_Def | 0.290 | −0.221 | −0.048 | −0.303 | 0.822 |
| Ln_Lat | −0.679 | 0.065 | −0.069 | 0.176 | 0.017 |
| % of variance | 30.97 | 14.40 | 11.62 | 11.21 | 7.90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oshchepkov, D.Y.; Makovka, Y.V.; Ponomarenko, M.P.; Redina, O.E.; Markel, A.L. Age-Dependent Changes in the Relationships between Traits Associated with the Pathogenesis of Stress-Sensitive Hypertension in ISIAH Rats. Int. J. Mol. Sci. 2023, 24, 10984. https://doi.org/10.3390/ijms241310984
Oshchepkov DY, Makovka YV, Ponomarenko MP, Redina OE, Markel AL. Age-Dependent Changes in the Relationships between Traits Associated with the Pathogenesis of Stress-Sensitive Hypertension in ISIAH Rats. International Journal of Molecular Sciences. 2023; 24(13):10984. https://doi.org/10.3390/ijms241310984
Chicago/Turabian StyleOshchepkov, Dmitry Yu., Yulia V. Makovka, Mikhail P. Ponomarenko, Olga E. Redina, and Arcady L. Markel. 2023. "Age-Dependent Changes in the Relationships between Traits Associated with the Pathogenesis of Stress-Sensitive Hypertension in ISIAH Rats" International Journal of Molecular Sciences 24, no. 13: 10984. https://doi.org/10.3390/ijms241310984