
Site-Specific RNA Editing of Stop Mutations in the CFTR mRNA of Human Bronchial Cultured Cells

 , , , and
, , , and 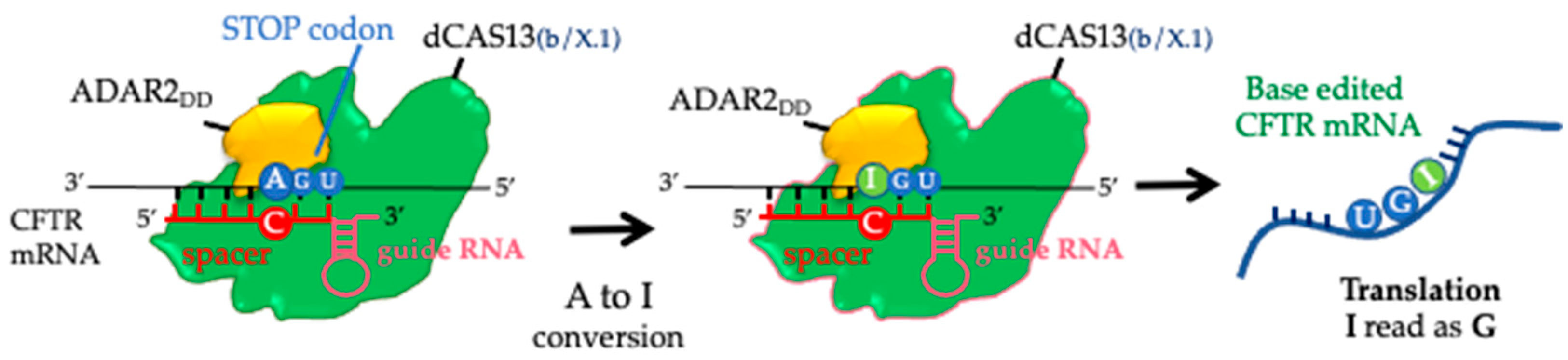{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
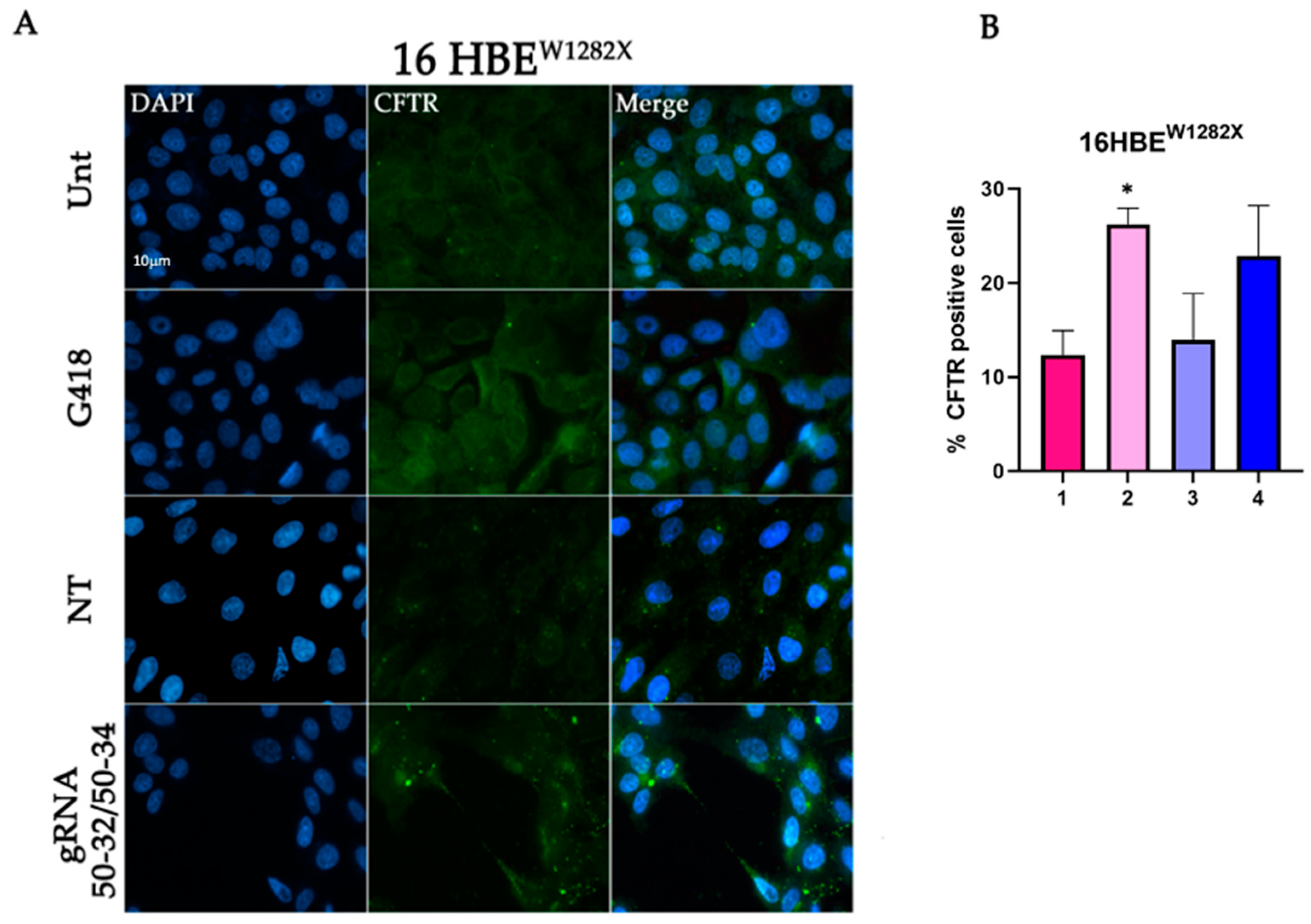
2.1. CFTR Protein Rescue in the 16HBE W1282X Cell Using the REPAIRv2 System
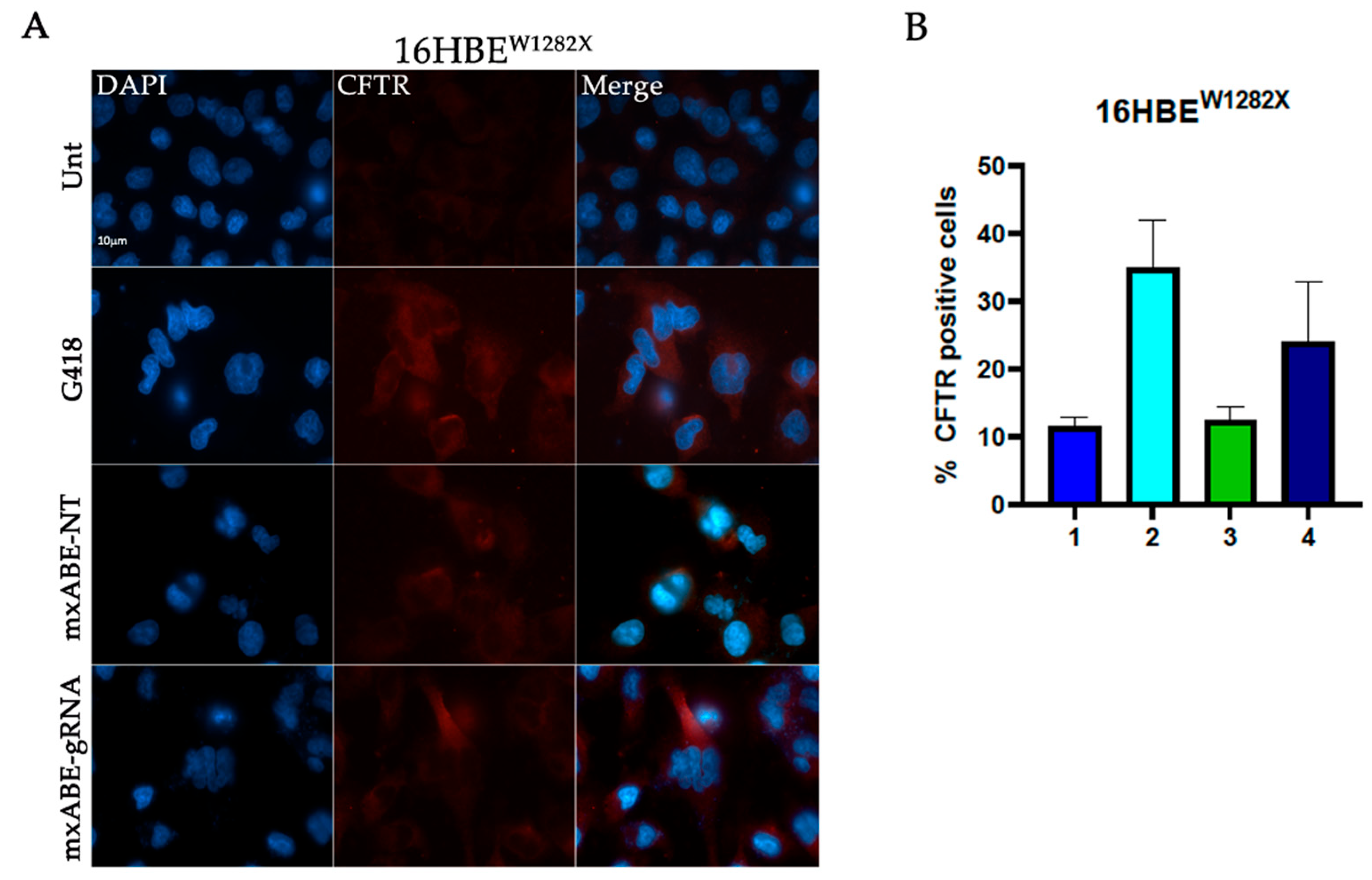
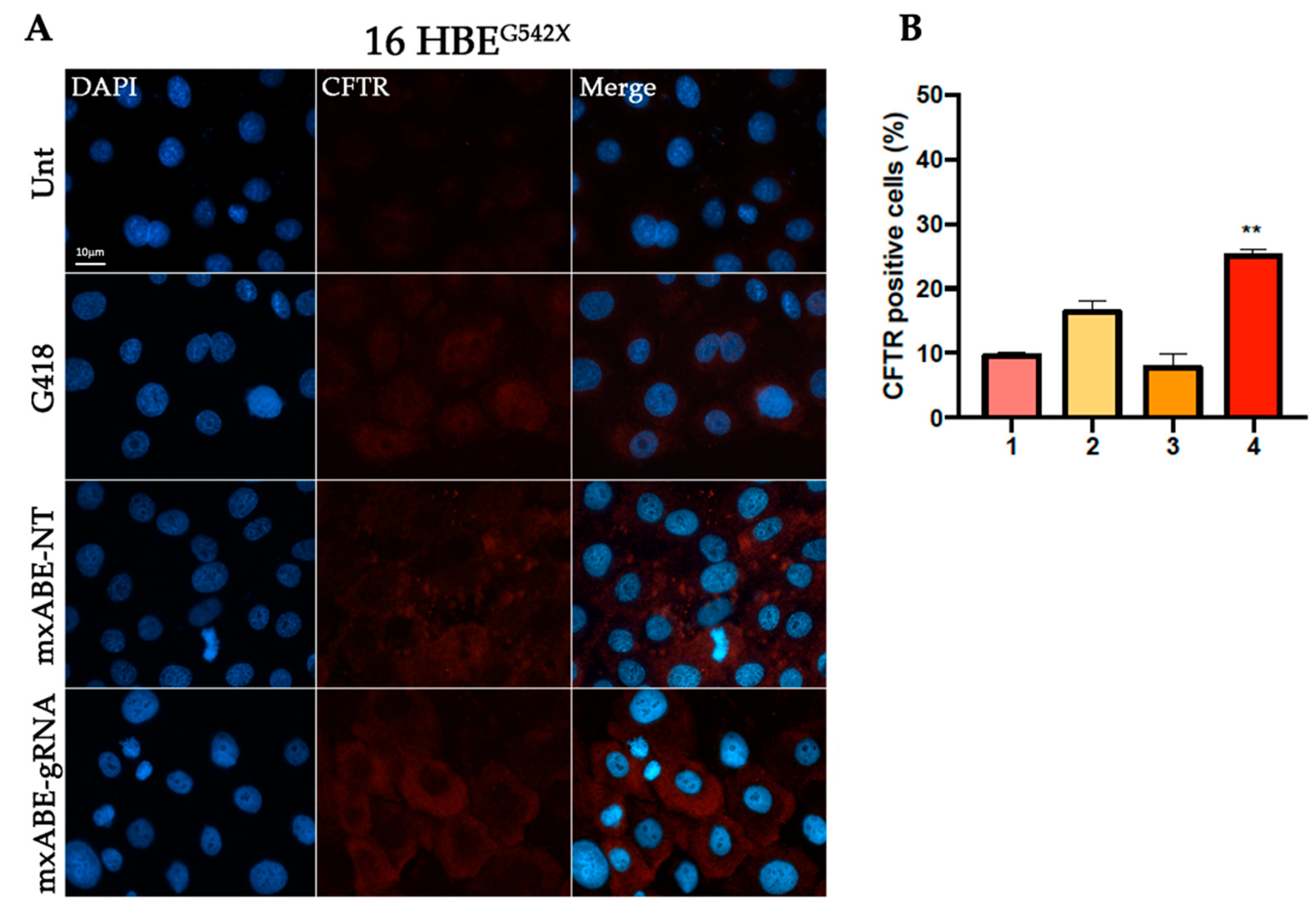
2.2. W1282X- and G542X-Specific gRNA Design and Cloning in the mxABE Platform
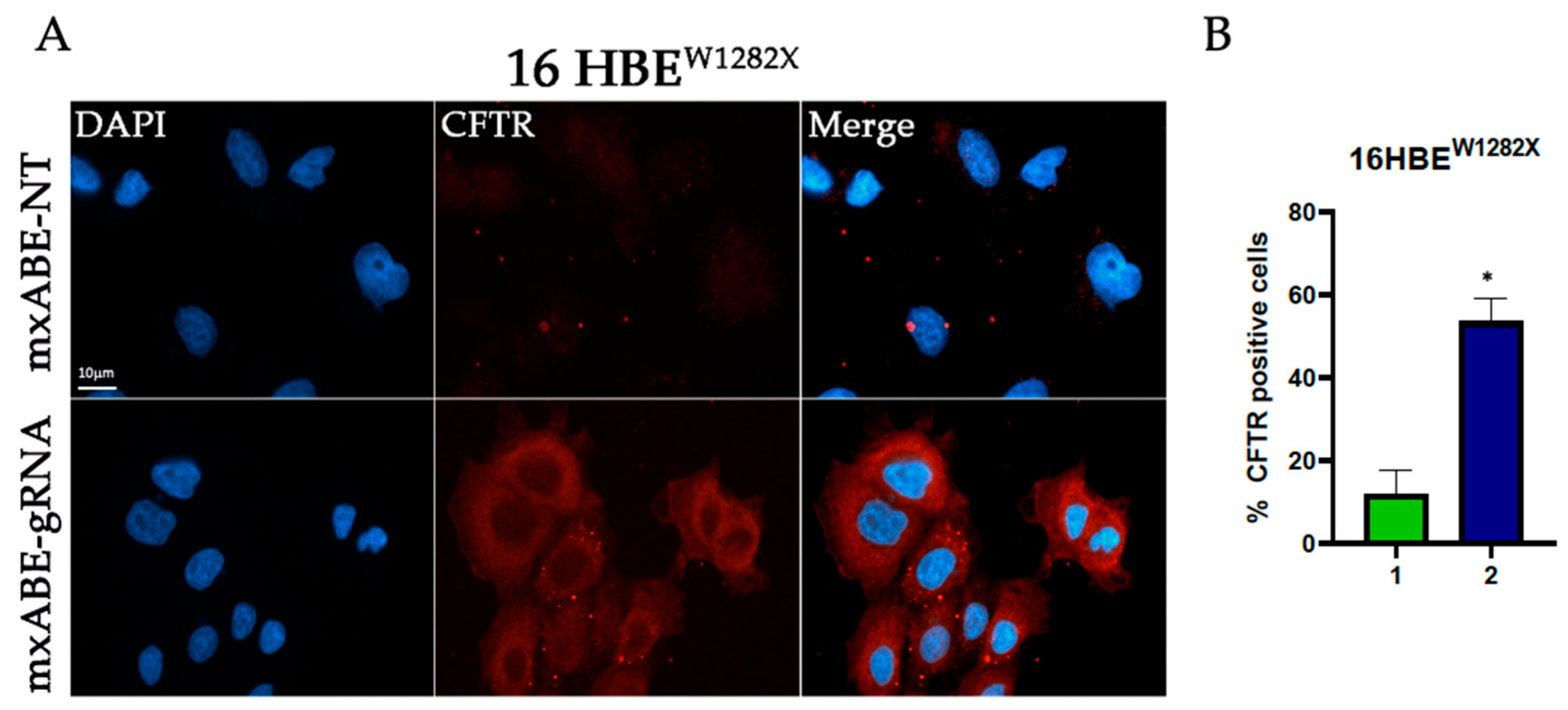
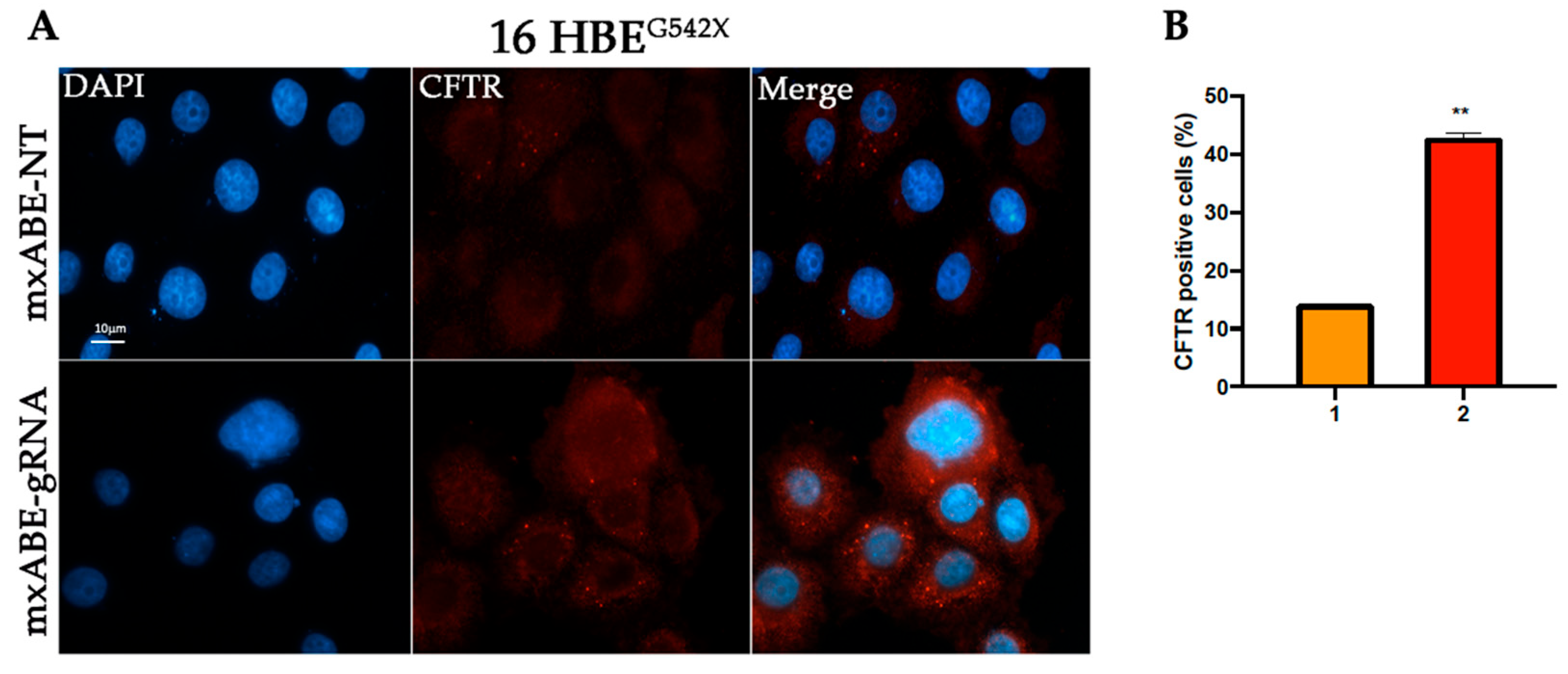
2.3. Evaluation Using Fluorescence Microscopy of CFTR Protein Recovery before and after Cell Sorting
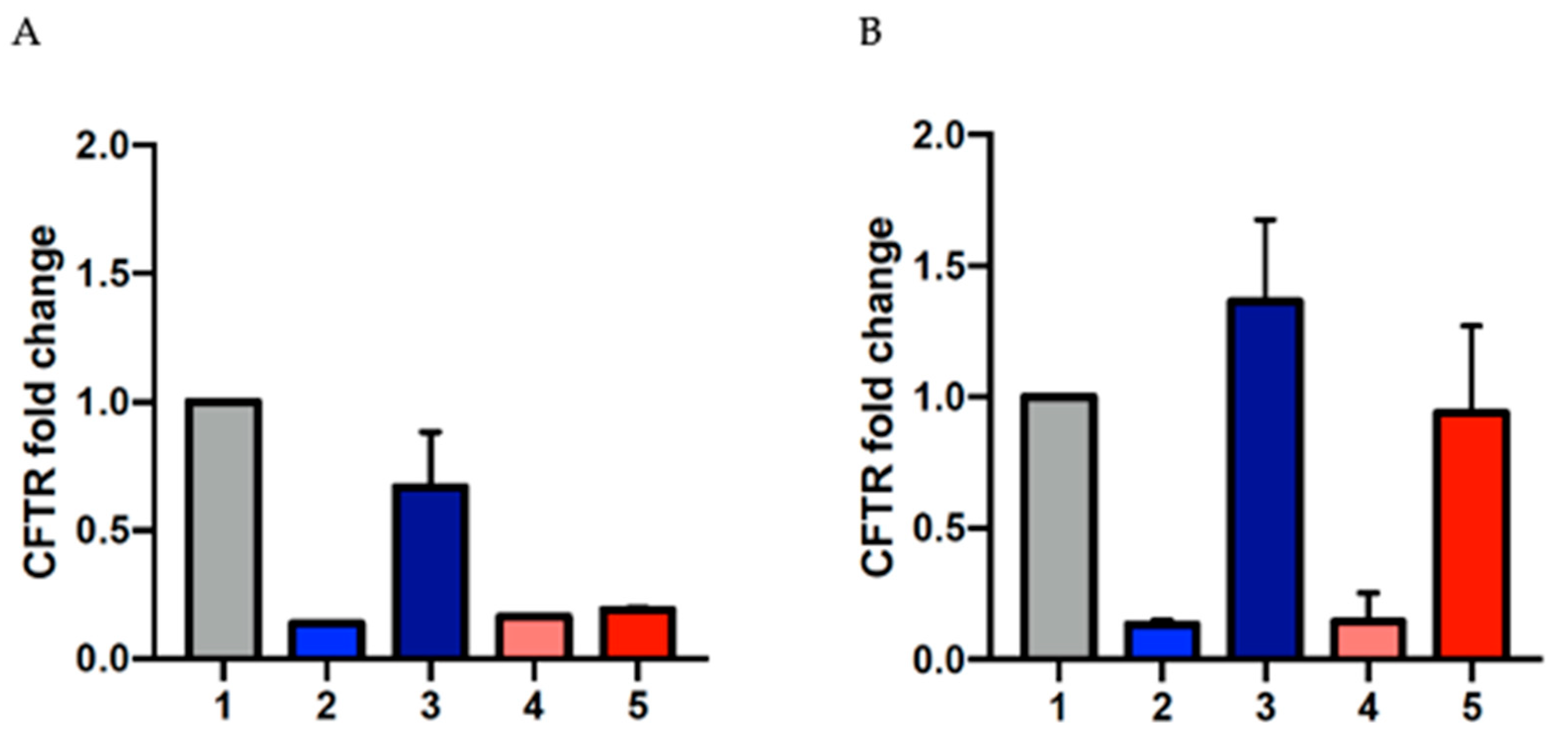
2.4. Evaluation of CFTR mRNA Levels Using RT-qPCR
3. Discussion
4. Materials and Methods
4.1. MinixABE Platform: Spacers Design and Cloning
4.2. Vectors and Clones
- -
- pC0055-CMV-dPspCas13b-GS-ADAR2DD(E488Q/T375G)-delta-984-1090, from Addgene (RRID: Addgene_103871);
- -
- pC0043-PspCas13b crRNA backbone, from Addgene (RRID: Addgene_103854);
- -
- CFTR-gRNA W1282X 50–32 and CFTR-gRNA W1282X 50-34, designed by us [22].
- -
- U6-BbsI-DR_CMV-minidCas13X.1-REPAIRv2-BGHpA_CMV-EGFP-BGHpA, from Addgene (RRID: Addgene_171383);
- -
- mxABE-NT, coding for a control gRNA (non-targeting) (Figure S2);
- -
- mxABE-CFW1282X 50–25, coding for a gRNA specific for the W1282X mutant region, 50 nts long, with a mismatch in position 25 with respect to 5′end (Figure S2);
- -
- mxABE-CFG542X 50–25, coding for a gRNA specific for the G542X mutant region, 50 nts long, with a mismatch in position 25 with respect to 5′end (Figure S2).
4.3. Cell Culture Conditions
4.4. Cell Transfection
4.5. RNA Extraction, cDNA Synthesis, and RT-qPCR
4.6. Immunofluorescence
4.7. Cell Sorting
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rowe, S.M.; Miller, S.; Sorscher, E.J. Cystic Fibrosis. N. Engl. J. Med. 2005, 352, 1992–2001. [Google Scholar] [CrossRef] [PubMed]
- Zabner, J.; Smith, J.J.; Karp, P.H.; Widdicombe, J.H.; Welsh, M.J. Loss of CFTR Chloride Channels Alters Salt Absorption by Cystic Fibrosis Airway Epithelia in Vitro. Mol. Cell 1998, 2, 397–403. [Google Scholar] [CrossRef]
- De Boeck, K.; Zolin, A.; Cuppens, H.; Olesen, H.V.; Viviani, L. The Relative Frequency of CFTR Mutation Classes in European Patients with Cystic Fibrosis. J. Cyst. Fibros. 2014, 13, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Aksit, M.A.; Bowling, A.D.; Evans, T.A.; Joynt, A.T.; Osorio, D.; Patel, S.; West, N.; Merlo, C.; Sosnay, P.R.; Cutting, G.R.; et al. Decreased MRNA and Protein Stability of W1282X Limits Response to Modulator Therapy. J. Cyst. Fibros. 2019, 18, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Kervestin, S.; Jacobson, A. NMD: A Multifaceted Response to Premature Translational Termination. Nat. Rev. Mol. Cell Biol. 2012, 13, 700–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popp, M.W.-L.; Maquat, L.E. Organizing Principles of Mammalian Nonsense-Mediated MRNA Decay. Annu. Rev. Genet. 2013, 47, 139–165. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Katrekar, D.; Mali, P. RNA-Guided Adenosine Deaminases: Advances and Challenges for Therapeutic RNA Editing. Biochemistry 2019, 58, 1947–1957. [Google Scholar] [CrossRef]
- Katrekar, D.; Chen, G.; Meluzzi, D.; Ganesh, A.; Worlikar, A.; Shih, Y.R.; Varghese, S.; Mali, P. In Vivo RNA Editing of Point Mutations via RNA-Guided Adenosine Deaminases. Nat. Methods 2019, 16, 239–242. [Google Scholar] [CrossRef]
- Katrekar, D.; Yen, J.; Xiang, Y.; Saha, A.; Meluzzi, D.; Savva, Y.; Mali, P. Efficient in Vitro and in Vivo RNA Editing via Recruitment of Endogenous ADARs Using Circular Guide RNAs. Nat. Biotechnol. 2022, 40, 938–945. [Google Scholar] [CrossRef]
- Booth, B.J.; Nourreddine, S.; Katrekar, D.; Savva, Y.; Bose, D.; Long, T.J.; Huss, D.J.; Mali, P. RNA Editing: Expanding the Potential of RNA Therapeutics. Mol. Ther. 2023, 31, 1533–1549. [Google Scholar] [CrossRef]
- Montiel-Gonzalez, M.F.; Vallecillo-Viejo, I.; Yudowski, G.A.; Rosenthal, J.J.C. Correction of Mutations within the Cystic Fibrosis Transmembrane Conductance Regulator by Site-Directed RNA Editing. Proc. Natl. Acad. Sci. USA 2013, 110, 18285–18290. [Google Scholar] [CrossRef] [Green Version]
- Savva, Y.A.; Rieder, L.E.; Reenan, R.A. The ADAR Protein Family. Genome Biol. 2012, 13, 252. [Google Scholar] [CrossRef] [Green Version]
- Li, J.B.; Levanon, E.Y.; Yoon, J.-K.; Aach, J.; Xie, B.; Leproust, E.; Zhang, K.; Gao, Y.; Church, G.M. Genome-Wide Identification of Human RNA Editing Sites by Parallel DNA Capturing and Sequencing. Science 2009, 324, 1210–1213. [Google Scholar] [CrossRef] [Green Version]
- Samuel, C.E. Adenosine Deaminase Acting on RNA (ADAR1), a Suppressor of Double-Stranded RNA–Triggered Innate Immune Responses. J. Biol. Chem. 2019, 294, 1710–1720. [Google Scholar] [CrossRef] [Green Version]
- Aquino-Jarquin, G. Novel Engineered Programmable Systems for ADAR-Mediated RNA Editing. Mol. Ther.—Nucleic Acids 2020, 19, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Basilio, C.; Wahba, A.J.; Lengyel, P.; Speyer, J.F.; Ochoa, S. Synthetic polynucleotides and the amino acid code, V. Proc. Natl. Acad. Sci. USA 1962, 48, 613–616. [Google Scholar] [CrossRef] [Green Version]
- Birgaoanu, M.; Sachse, M.; Gatsiou, A. RNA Editing Therapeutics: Advances, Challenges and Perspectives on Combating Heart Disease. Cardiovasc. Drugs Ther. 2023, 37, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, H.M.; Jantsch, M.F. Site-Directed RNA Editing: Recent Advances and Open Challenges. RNA Biol. 2021, 18, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.B.T.; Gootenberg, J.S.; Abudayyeh, O.O.; Franklin, B.; Kellner, M.J.; Joung, J.; Zhang, F. RNA Editing with CRISPR-Cas13. Science 2017, 358, 1019–1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Zhou, Y.; Xiao, Q.; He, B.; Geng, G.; Wang, Z.; Cao, B.; Dong, X.; Bai, W.; Wang, Y.; et al. Programmable RNA Editing with Compact CRISPR–Cas13 Systems from Uncultivated Microbes. Nat. Methods 2021, 18, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Valley, H.C.; Bukis, K.M.; Bell, A.; Cheng, Y.; Wong, E.; Jordan, N.J.; Allaire, N.E.; Sivachenko, A.; Liang, F.; Bihler, H.; et al. Isogenic Cell Models of Cystic Fibrosis-Causing Variants in Natively Expressing Pulmonary Epithelial Cells. J. Cyst. Fibros. 2019, 18, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Melfi, R.; Cancemi, P.; Chiavetta, R.; Barra, V.; Lentini, L.; Di, A. Investigating REPAIRv2 as a Tool to Edit CFTR MRNA 2 with Premature Stop Codons 3. Int. J. Mol. Sci. 2020, 21, 4781. [Google Scholar] [CrossRef] [PubMed]
- Burke, J.F.; Mogg, A.E. Suppression of a Nonsense Mutation in Mammalian Cells in Vivo by the Aminoglycoside Anthiotics G–418 and Paromomycin. Nucleic Acids Res. 1985, 13, 6265–6272. [Google Scholar] [CrossRef] [PubMed]
- Maquat, L.E. Nonsense-Mediated MRNA Decay in Mammals. J. Cell Sci. 2005, 118, 1773–1776. [Google Scholar] [CrossRef] [Green Version]
- Lentini, L.; Melfi, R.; Di Leonardo, A.; Spinello, A.; Barone, G.; Pace, A.; Palumbo Piccionello, A.; Pibiri, I. Toward a Rationale for the PTC124 (Ataluren) Promoted Readthrough of Premature Stop Codons: A Computational Approach and GFP-Reporter Cell-Based Assay. Mol. Pharm. 2014, 11, 653–664. [Google Scholar] [CrossRef]
- Pibiri, I.; Lentini, L.; Melfi, R.; Gallucci, G.; Pace, A.; Spinello, A.; Barone, G.; Di Leonardo, A. Enhancement of Premature Stop Codon Readthrough in the CFTR Gene by Ataluren (PTC124) Derivatives. Eur. J. Med. Chem. 2015, 101, 236–244. [Google Scholar] [CrossRef]
- Pibiri, I.; Lentini, L.; Melfi, R.; Tutone, M.; Baldassano, S.; Ricco Galluzzo, P.; Di Leonardo, A.; Pace, A. Rescuing the CFTR Protein Function: Introducing 1,3,4-Oxadiazoles as Translational Readthrough Inducing Drugs. Eur. J. Med. Chem. 2018, 159, 126–142. [Google Scholar] [CrossRef]
- Campofelice, A.; Lentini, L.; Di Leonardo, A.; Melfi, R.; Tutone, M.; Pace, A.; Pibiri, I. Strategies against Nonsense: Oxadiazoles as Translational Readthrough-Inducing Drugs (TRIDs). Int. J. Mol. Sci. 2019, 20, 3329. [Google Scholar] [CrossRef] [Green Version]
- Tutone, M.; Pibiri, I.; Perriera, R.; Campofelice, A.; Culletta, G.; Melfi, R.; Pace, A.; Almerico, A.M.; Lentini, L. Pharmacophore-Based Design of New Chemical Scaffolds as Translational Readthrough-Inducing Drugs (TRIDs). ACS Med. Chem. Lett. 2020, 11, 747–753. [Google Scholar] [CrossRef]
- Pibiri, I.; Melfi, R.; Tutone, M.; Di Leonardo, A.; Pace, A.; Lentini, L. Targeting Nonsense: Optimization of 1,2,4-Oxadiazole Trids to Rescue Cftr Expression and Functionality in Cystic Fibrosis Cell Model Systems. Int. J. Mol. Sci. 2020, 21, 6420. [Google Scholar] [CrossRef]
- Kerem, E. ELX-02: An Investigational Read-through Agent for the Treatment of Nonsense Mutation-Related Genetic Disease. Expert Opin. Investig. Drugs 2020, 29, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Vallecillo-Viejo, I.C.; Liscovitch-Brauer, N.; Montiel-Gonzalez, M.F.; Eisenberg, E.; Rosenthal, J.J.C. Abundant Off-Target Edits from Site-Directed RNA Editing Can Be Reduced by Nuclear Localization of the Editing Enzyme. RNA Biol. 2018, 15, 104–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montiel-Gonzalez, M.F.; Diaz Quiroz, J.F.; Rosenthal, J.J.C. Current Strategies for Site-Directed RNA Editing Using ADARs. Methods 2019, 156, 16–24. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiavetta, R.F.; Titoli, S.; Barra, V.; Cancemi, P.; Melfi, R.; Di Leonardo, A. Site-Specific RNA Editing of Stop Mutations in the CFTR mRNA of Human Bronchial Cultured Cells. Int. J. Mol. Sci. 2023, 24, 10940. https://doi.org/10.3390/ijms241310940
Chiavetta RF, Titoli S, Barra V, Cancemi P, Melfi R, Di Leonardo A. Site-Specific RNA Editing of Stop Mutations in the CFTR mRNA of Human Bronchial Cultured Cells. International Journal of Molecular Sciences. 2023; 24(13):10940. https://doi.org/10.3390/ijms241310940
Chicago/Turabian StyleChiavetta, Roberta F., Simona Titoli, Viviana Barra, Patrizia Cancemi, Raffaella Melfi, and Aldo Di Leonardo. 2023. "Site-Specific RNA Editing of Stop Mutations in the CFTR mRNA of Human Bronchial Cultured Cells" International Journal of Molecular Sciences 24, no. 13: 10940. https://doi.org/10.3390/ijms241310940