
Repetitive Low-Level Blast Exposure via Akt/NF-κB Signaling Pathway Mediates the M1 Polarization of Mouse Alveolar Macrophage MH-S Cells
 and
and
Abstract
:1. Introduction
2. Results
2.1. rLLB Exposure Caused Morphological Changes and Polarization toward M1 Phenotype of MH-S Cells
2.2. rLLB Exposure Induced Oxidative Stress State Changes and Early Apoptosis of MH-S Cells
2.3. rLLB Exposure Activated the Akt/NF-κB Signaling Pathway in MH-S Cells
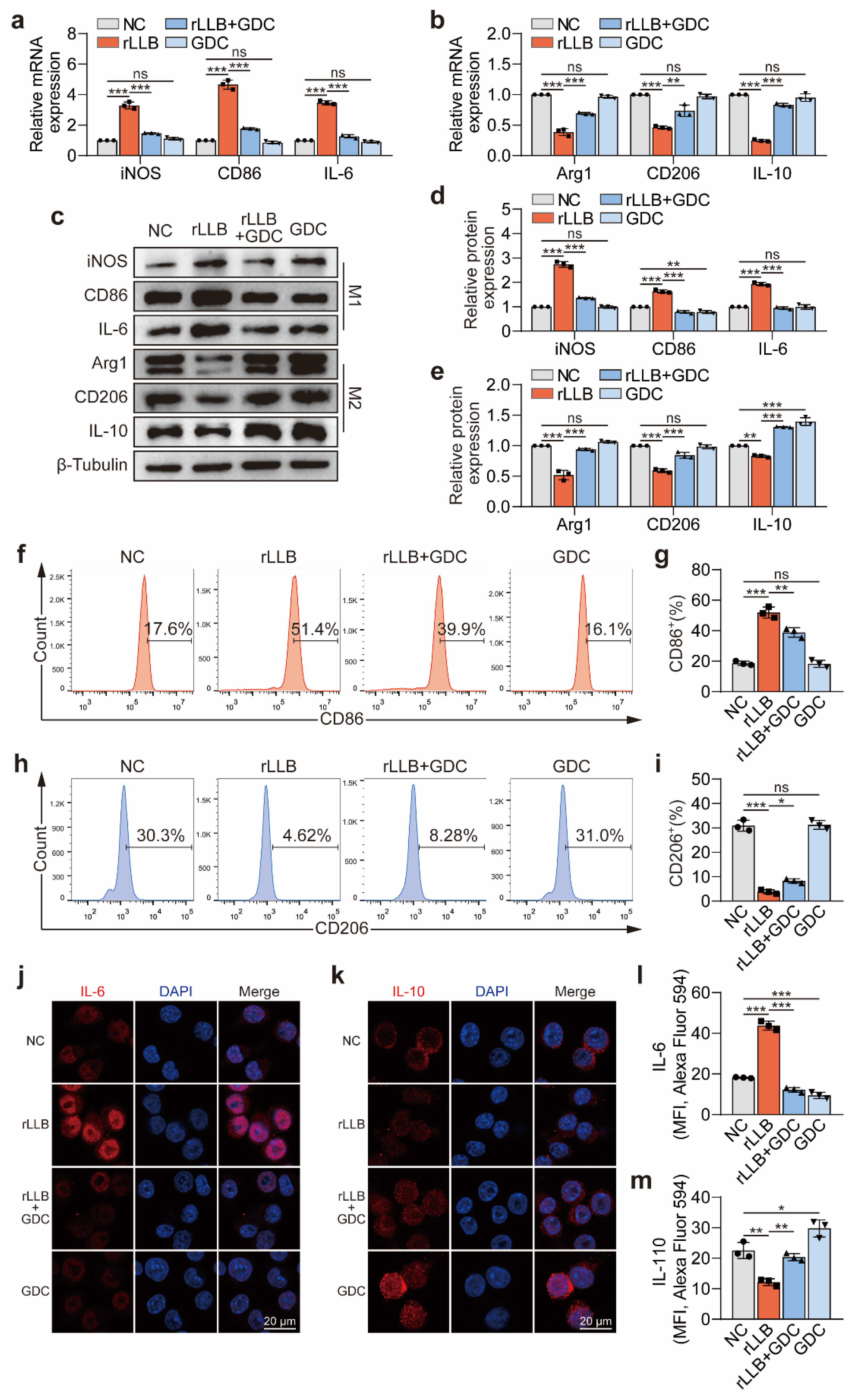
2.4. Inhibition of the Akt/NF-κB Signaling Pathway Downregulated MH-S Cell Phenotypic Changes Induced by rLLB Exposure
2.5. Inhibition of the Akt/NF-κB Signaling Pathway Had No Significant Effect on Early Apoptosis of MH-S Cells Induced by rLLB Exposure
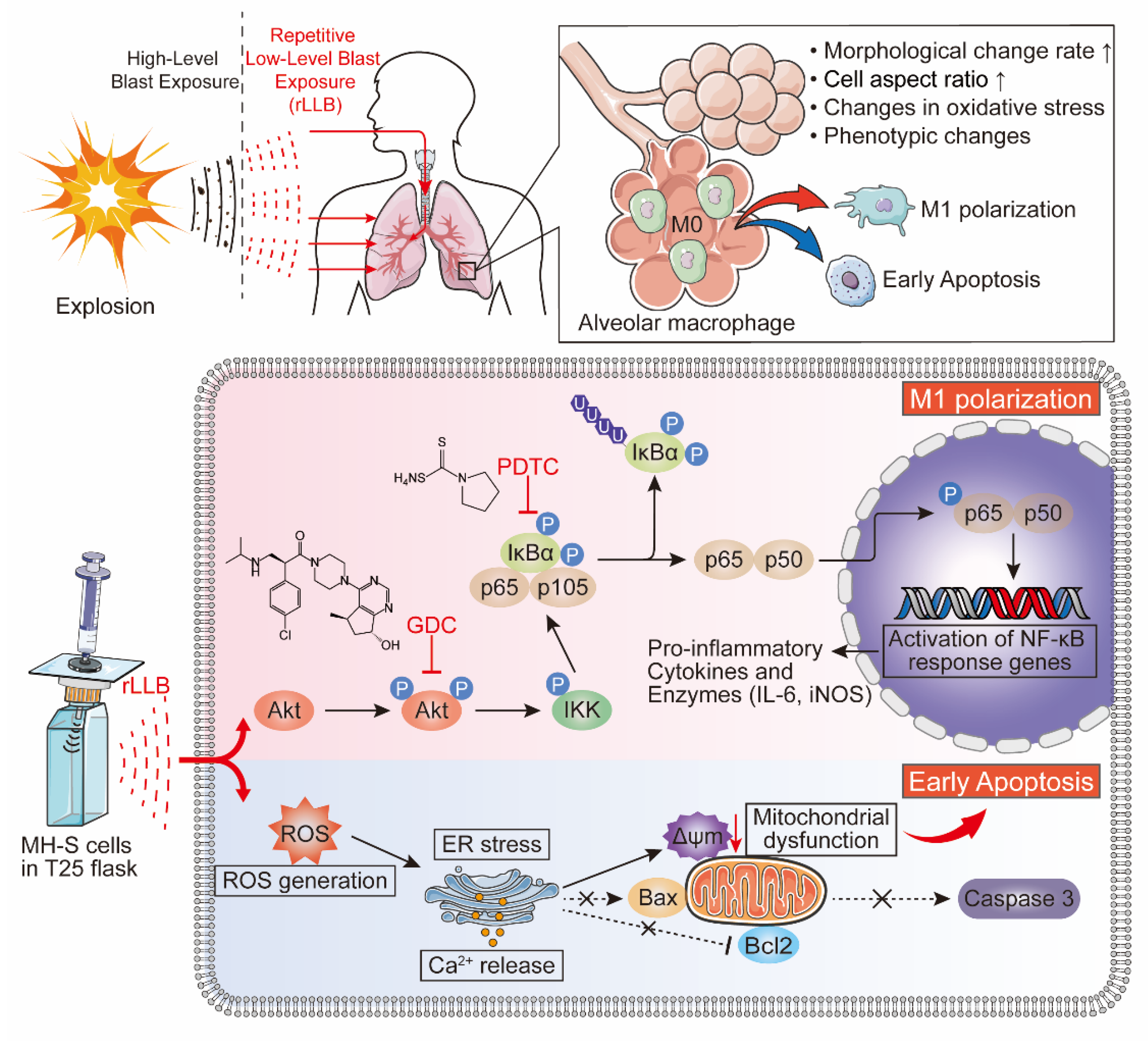
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Grouping
4.2. Miniature Manual Reddy-Tube Device for rLLB Generation
4.3. Observation of Cell Morphology and Calculation of Morphological Change Rate and Aspect Ratio
4.4. Cytoskeleton Staining
4.5. Real-Time Quantitative PCR (qPCR)
4.6. Western Blotting (WB) Analysis
4.7. Immunofluorescence
4.8. ROS Staining and Mitochondrial Membrane Potential Detection
4.9. Flow Cytometry
4.10. Cell Viability Assay
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMs | Alveolar macrophages |
| Arg1 | Arginase 1 |
| ARDS | Acute respiratory distress syndrome |
| BOP | Blast overpressure |
| CD | Cluster of differentiation |
| COPD | Chronic obstructive pulmonary disease |
| CCK-8 | Cell counting kit-8 |
| DSS | Dextran sulfate sodium |
| FBS | Fetal bovine serum |
| HLB | High-level blast |
| HBSS | Hank’s balanced salt solution |
| HRP | Horseradish peroxidase |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| IC50 | Half maximal inhibitory concentration |
| IFN | Interferon |
| LPS | Lipopolysaccharide |
| NC | Normal control |
| NDAA | National Defense Authorization Act |
| NF-κB | Nuclear factor kappa-B |
| OD | Optical density |
| PVDF | Polyvinylidene fluoride |
| PTSD | Post-traumatic stress disorder |
| qPCR | Real-Time Quantitative PCR |
| rLLB | Repetitive low-level blast |
| ROS | Reactive oxygen species |
| SD | Standard deviation |
| TNF | Tumor necrosis factor |
| TGF | Transforming growth factor |
| TBI | Traumatic brain injuries |
| TPL | Triptolide |
| WB | Western blotting |
References
- Wang, X.; Du, J.; Zhuang, Z.; Wang, Z.G.; Jiang, J.X.; Yang, C. Incidence, casualties and risk characteristics of civilian explosion blast injury in China: 2000–2017 data from the state Administration of Work Safety. Mil. Med. Res. 2020, 7, 29. [Google Scholar] [CrossRef]
- Wild, H.; Stewart, B.T.; LeBoa, C.; Stave, C.D.; Wren, S.M. Epidemiology of Injuries Sustained by Civilians and Local Combatants in Contemporary Armed Conflict: An Appeal for a Shared Trauma Registry Among Humanitarian Actors. World J. Surg. 2020, 44, 1863–1873. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.J.; Bebarta, V.S.; Bonnett, C.J.; Pons, P.T.; Cantrill, S.V. Blast injuries. Lancet 2009, 374, 405–415. [Google Scholar] [CrossRef]
- Mathews, Z.R.; Koyfman, A. Blast Injuries. J. Emerg. Med. 2015, 49, 573–587. [Google Scholar] [CrossRef]
- Li, N.; Geng, C.; Hou, S.; Fan, H.; Gong, Y. Damage-Associated Molecular Patterns and Their Signaling Pathways in Primary Blast Lung Injury: New Research Progress and Future Directions. Int. J. Mol. Sci. 2020, 21, 6303. [Google Scholar] [CrossRef]
- Vartanian, O.; Tenn, C.; Rhind, S.G.; Nakashima, A.; Di Battista, A.P.; Sergio, L.E.; Gorbet, D.J.; Fraser, D.D.; Colantonio, A.; King, K.; et al. Blast in Context: The Neuropsychological and Neurocognitive Effects of Long-Term Occupational Exposure to Repeated Low-Level Explosives on Canadian Armed Forces’ Breaching Instructors and Range Staff. Front. Neurol. 2020, 11, 588531. [Google Scholar] [CrossRef]
- Perez Garcia, G.; Perez, G.M.; De Gasperi, R.; Gama Sosa, M.A.; Otero-Pagan, A.; Pryor, D.; Abutarboush, R.; Kawoos, U.; Hof, P.R.; Cook, D.G.; et al. Progressive Cognitive and Post-Traumatic Stress Disorder-Related Behavioral Traits in Rats Exposed to Repetitive Low-Level Blast. J. Neurotrauma 2021, 38, 2030–2045. [Google Scholar] [CrossRef] [PubMed]
- Vartanian, O.; Coady, L.; Blackler, K.; Fraser, B.; Cheung, B. Neuropsychological, Neurocognitive, Vestibular, and Neuroimaging Correlates of Exposure to Repetitive Low-Level Blast Waves: Evidence from Four Nonoverlapping Samples of Canadian Breachers. Mil. Med. 2021, 186, e393–e400. [Google Scholar] [CrossRef]
- Nakashima, A.; Vartanian, O.; Rhind, S.G.; King, K.; Tenn, C.; Jetly, C.R. Repeated Occupational Exposure to Low-level Blast in the Canadian Armed Forces: Effects on Hearing, Balance, and Ataxia. Mil. Med. 2021, 187, e201–e208. [Google Scholar] [CrossRef]
- Solis, A.G.; Bielecki, P.; Steach, H.R.; Sharma, L.; Harman, C.C.D.; Yun, S.; de Zoete, M.R.; Warnock, J.N.; To, S.D.F.; York, A.G.; et al. Mechanosensation of cyclical force by PIEZO1 is essential for innate immunity. Nature 2019, 573, 69–74. [Google Scholar] [CrossRef]
- Chen, K.; Henn, D.; Sivaraj, D.; Bonham, C.A.; Griffin, M.; Kussie, H.C.; Padmanabhan, J.; Trotsyuk, A.A.; Wan, D.C.; Januszyk, M.; et al. Mechanical Strain Drives Myeloid Cell Differentiation Toward Proinflammatory Subpopulations. Adv. Wound Care 2022, 11, 466–478. [Google Scholar] [CrossRef]
- Looney, M.R.; Headley, M.B. Live imaging of the pulmonary immune environment. Cell. Immunol. 2020, 350, 103862. [Google Scholar] [CrossRef]
- Feng, Z.; Zhou, J.; Liu, Y.; Xia, R.; Li, Q.; Yan, L.; Chen, Q.; Chen, X.; Jiang, Y.; Chao, G.; et al. Epithelium- and endothelium-derived exosomes regulate the alveolar macrophages by targeting RGS1 mediated calcium signaling-dependent immune response. Cell Death Differ. 2021, 28, 2238–2256. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Wang, Z.; Qu, Y.; Zhu, H.; Zhu, Q.; Tong, W.; Bao, L.; Lv, Q.; Cong, J.; Li, D.; et al. Distinct uptake, amplification, and release of SARS-CoV-2 by M1 and M2 alveolar macrophages. Cell Discov. 2021, 7, 24. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Dagvadorj, J.; Shimada, K.; Chen, S.; Jones, H.D.; Tumurkhuu, G.; Zhang, W.; Wawrowsky, K.A.; Crother, T.R.; Arditi, M. Lipopolysaccharide Induces Alveolar Macrophage Necrosis via CD14 and the P2X7 Receptor Leading to Interleukin-1α Release. Immunity 2015, 42, 640–653. [Google Scholar] [CrossRef] [Green Version]
- Nahrendorf, M.; Swirski, F.K. Abandoning M1/M2 for a Network Model of Macrophage Function. Circ. Res. 2016, 119, 414–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oishi, Y.; Manabe, I. Macrophages in inflammation, repair and regeneration. Int. Immunol. 2018, 30, 511–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Stijn, C.M.; Kim, J.; Lusis, A.J.; Barish, G.D.; Tangirala, R.K. Macrophage polarization phenotype regulates adiponectin receptor expression and adiponectin anti-inflammatory response. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 636–649. [Google Scholar] [CrossRef] [Green Version]
- Hamidzadeh, K.; Christensen, S.M.; Dalby, E.; Chandrasekaran, P.; Mosser, D.M. Macrophages and the Recovery from Acute and Chronic Inflammation. Annu. Rev. Physiol. 2017, 79, 567–592. [Google Scholar] [CrossRef] [Green Version]
- Funes, S.C.; Rios, M.; Escobar-Vera, J.; Kalergis, A.M. Implications of macrophage polarization in autoimmunity. Immunology 2018, 154, 186–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosser, D.M.; Hamidzadeh, K.; Goncalves, R. Macrophages and the maintenance of homeostasis. Cell. Mol. Immunol. 2021, 18, 579–587. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhang, Y.; Xie, F.; Dou, X.; Han, M.; Zhang, H. Role of PI3K/Akt/NF-κB and GSK-3β pathways in the rat model of cardiopulmonary bypass-related lung injury. Biomed. Pharmacother. 2018, 106, 747–754. [Google Scholar] [CrossRef]
- Vergadi, E.; Ieronymaki, E.; Lyroni, K.; Vaporidi, K.; Tsatsanis, C. Akt Signaling Pathway in Macrophage Activation and M1/M2 Polarization. J. Immunol. 2017, 198, 1006–1014. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Sun, L.; Wang, G.; Chen, B.; Luo, F. RUNX1: A Regulator of NF-kB Signaling in Pulmonary Diseases. Curr. Protein Pept. Sci. 2018, 19, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Dorrington, M.G.; Fraser, I.D.C. NF-κB Signaling in Macrophages: Dynamics, Crosstalk, and Signal Integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef]
- Zheng, H.; Liang, W.; He, W.; Huang, C.; Chen, Q.; Yi, H.; Long, L.; Deng, Y.; Zeng, M. Ghrelin attenuates sepsis-induced acute lung injury by inhibiting the NF-κB, iNOS, and Akt signaling in alveolar macrophages. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 317, L381–L391. [Google Scholar] [CrossRef]
- Jarrar, D.; Kuebler, J.F.; Rue, L.W., 3rd; Matalon, S.; Wang, P.; Bland, K.I.; Chaudry, I.H. Alveolar macrophage activation after trauma-hemorrhage and sepsis is dependent on NF-kappaB and MAPK/ERK mechanisms. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 283, L799–L805. [Google Scholar] [CrossRef] [Green Version]
- Le, Y.; Wang, Y.; Zhou, L.; Xiong, J.; Tian, J.; Yang, X.; Gai, X.; Sun, Y. Cigarette smoke-induced HMGB1 translocation and release contribute to migration and NF-κB activation through inducing autophagy in lung macrophages. J. Cell. Mol. Med. 2020, 24, 1319–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, D.K.; Gardner, I.D.; Lawton, J.W. Macrophage function in Mycobacterium lepraemurium infection: Morphological and functional changes of peritoneal and splenic macrophages in vitro. Infect. Immun. 1983, 39, 353–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.Q.; Zhang, L.; Zhou, Y.L.; Tang, J.B. Morphological changes of macrophages and their potential contribution to tendon healing. Colloids Surf. B Biointerfaces 2022, 209, 112145. [Google Scholar] [CrossRef] [PubMed]
- Fresta, C.G.; Fidilio, A.; Lazzarino, G.; Musso, N.; Grasso, M.; Merlo, S.; Amorini, A.M.; Bucolo, C.; Tavazzi, B.; Lazzarino, G.; et al. Modulation of Pro-Oxidant and Pro-Inflammatory Activities of M1 Macrophages by the Natural Dipeptide Carnosine. Int. J. Mol. Sci. 2020, 21, 776. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Shi, F.; Zhou, Z.; Sun, F.; Sun, M.H.; Sun, Q.; Chen, L.; Li, D.; Jiang, C.Y.; Zhao, R.Z.; et al. M1 macrophage mediated increased reactive oxygen species (ROS) influence wound healing via the MAPK signaling in vitro and in vivo. Toxicol. Appl. Pharmacol. 2019, 366, 83–95. [Google Scholar] [CrossRef]
- Kannan, K.; Jain, S.K. Oxidative stress and apoptosis. Pathophysiol. Off. J. Int. Soc. Pathophysiol. 2000, 7, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Yin, L.; Li, R.; Xu, L.; Xu, Y.; Han, X.; Qi, Y. Dioscin alleviates lung ischemia/reperfusion injury by regulating FXR-mediated oxidative stress, apoptosis, and inflammation. Eur. J. Pharmacol. 2021, 908, 174321. [Google Scholar] [CrossRef]
- Westrol, M.S.; Donovan, C.M.; Kapitanyan, R. Blast Physics and Pathophysiology of Explosive Injuries. Ann. Emerg. Med. 2017, 69, S4–S9. [Google Scholar] [CrossRef]
- Scott, T.E.; Kirkman, E.; Haque, M.; Gibb, I.E.; Mahoney, P.; Hardman, J.G. Primary blast lung injury—A review. Br. J. Anaesth. 2017, 118, 311–316. [Google Scholar] [CrossRef] [Green Version]
- McWhorter, F.Y.; Wang, T.; Nguyen, P.; Chung, T.; Liu, W.F. Modulation of macrophage phenotype by cell shape. Proc. Natl. Acad. Sci. USA 2013, 110, 17253–17258. [Google Scholar] [CrossRef] [Green Version]
- Jain, N.; Moeller, J.; Vogel, V. Mechanobiology of Macrophages: How Physical Factors Coregulate Macrophage Plasticity and Phagocytosis. Annu. Rev. Biomed. Eng. 2019, 21, 267–297. [Google Scholar] [CrossRef]
- Smith, T.D.; Tse, M.J.; Read, E.L.; Liu, W.F. Regulation of macrophage polarization and plasticity by complex activation signals. Integr. Biol. Quant. Biosci. Nano Macro 2016, 8, 946–955. [Google Scholar] [CrossRef] [Green Version]
- Juhas, U.; Ryba-Stanisławowska, M.; Szargiej, P.; Myśliwska, J. Different pathways of macrophage activation and polarization. Postep. Hig. I Med. Dosw. 2015, 69, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Qu, K.; Shen, N.Y.; Xu, X.S.; Su, H.B.; Wei, J.C.; Tai, M.H.; Meng, F.D.; Zhou, L.; Zhang, Y.L.; Liu, C. Emodin induces human T cell apoptosis in vitro by ROS-mediated endoplasmic reticulum stress and mitochondrial dysfunction. Acta Pharmacol. Sin. 2013, 34, 1217–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rendra, E.; Riabov, V.; Mossel, D.M.; Sevastyanova, T.; Harmsen, M.C.; Kzhyshkowska, J. Reactive oxygen species (ROS) in macrophage activation and function in diabetes. Immunobiology 2019, 224, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.; Summer, R.; Wilson, A.A.; Kotton, D.N.; Fine, A. The prolonged life-span of alveolar macrophages. Am. J. Respir. Cell Mol. Biol. 2008, 38, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Misistia, A.; Skotak, M.; Cardenas, A.; Alay, E.; Chandra, N.; Kamimori, G.H. Sensor orientation and other factors which increase the blast overpressure reporting errors. PLoS ONE 2020, 15, e0240262. [Google Scholar] [CrossRef]
- Cheng, H.; Sun, L.; Shen, D.; Ren, A.; Ma, F.; Tai, G.; Fan, L.; Zhou, Y. Beta-1,6 glucan converts tumor-associated macrophages into an M1-like phenotype. Carbohydr. Polym. 2020, 247, 116715. [Google Scholar] [CrossRef]
- Embgenbroich, M.; van der Zande, H.J.P.; Hussaarts, L.; Schulte-Schrepping, J.; Pelgrom, L.R.; García-Tardón, N.; Schlautmann, L.; Stoetzel, I.; Händler, K.; Lambooij, J.M.; et al. Soluble mannose receptor induces proinflammatory macrophage activation and metaflammation. Proc. Natl. Acad. Sci. USA 2021, 118, e2103304118. [Google Scholar] [CrossRef]
- Tang, B.; Zhu, J.; Zhang, B.; Wu, F.; Wang, Y.; Weng, Q.; Fang, S.; Zheng, L.; Yang, Y.; Qiu, R.; et al. Therapeutic Potential of Triptolide as an Anti-Inflammatory Agent in Dextran Sulfate Sodium-Induced Murine Experimental Colitis. Front. Immunol. 2020, 11, 592084. [Google Scholar] [CrossRef]
- Mallis, P.; Chatzistamatiou, T.; Dimou, Z.; Sarri, E.F.; Georgiou, E.; Salagianni, M.; Triantafyllia, V.; Andreakos, E.; Stavropoulos-Giokas, C.; Michalopoulos, E. Mesenchymal stromal cell delivery as a potential therapeutic strategy against COVID-19: Promising evidence from in vitro results. World J. Biol. Chem. 2022, 13, 47–65. [Google Scholar] [CrossRef]
- Sundaram, S.; Sellamuthu, K.; Nagavelu, K.; Suma, H.R.; Das, A.; Narayan, R.; Chakravortty, D.; Gopalan, J.; Eswarappa, S.M. Stimulation of angiogenesis using single-pulse low-pressure shock wave treatment. J. Mol. Med. 2018, 96, 1177–1187. [Google Scholar] [CrossRef]
- Mbawuike, I.N.; Herscowitz, H.B. MH-S, a murine alveolar macrophage cell line: Morphological, cytochemical, and functional characteristics. J. Leukoc. Biol. 1989, 46, 119–127. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| iNOS | GTTCTCAGCCCAACAATACAAGA | GTGGACGGGTCGATGTCAC |
| CD86 | TGTTTCCGTGGAGACGCAAG | TTGAGCCTTTGTAAATGGGCA |
| IL-6 | CCAAGAGGTGAGTGCTTCCC | CTGTTGTTCAGACTCTCTCCCT |
| Arg1 | CTCCAAGCCAAAGTCCTTAGAG | AGGAGCTGTCATTAGGGACATC |
| CD206 | CTCTGTTCAGCTATTGGACGC | CGGAATTTCTGGGATTCAGCTTC |
| IL-10 | GCTCTTACTGACTGGCATGAG | CGCAGCTCTAGGAGCATGTG |
| Bax | AGGCGAATTGGCGATGAACTGG | CTAGCAAAGTAGAAAAGGGCAACC |
| Bcl2 | ATGCCTTTGTGGAACTATATGGC | GGTATGCACCCAGAGTGATGC |
| Caspase3 | TGGTGATGAAGGGGTCATTTATG | TTCGGCTTTCCAGTCAGACTC |
| Akt | ATGAACGACGTAGCCATTGTG | TTGTAGCCAATAAAGGTGCCAT |
| p105 | ATGGCAGACGATGATCCCTAC | TGTTGACAGTGGTATTTCTGGTG |
| p65 | AGGCTTCTGGGCCTTATGTG | TGCTTCTCTCGCCAGGAATAC |
| GAPDH | AGGTCGGTGTGAACGGATTTG | TGTAGACCATGTAGTTGAGGTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, C.; Wang, X.; Chen, J.; Sun, N.; Wang, Y.; Li, Z.; Han, L.; Hou, S.; Fan, H.; Li, N.; et al. Repetitive Low-Level Blast Exposure via Akt/NF-κB Signaling Pathway Mediates the M1 Polarization of Mouse Alveolar Macrophage MH-S Cells. Int. J. Mol. Sci. 2023, 24, 10596. https://doi.org/10.3390/ijms241310596
Geng C, Wang X, Chen J, Sun N, Wang Y, Li Z, Han L, Hou S, Fan H, Li N, et al. Repetitive Low-Level Blast Exposure via Akt/NF-κB Signaling Pathway Mediates the M1 Polarization of Mouse Alveolar Macrophage MH-S Cells. International Journal of Molecular Sciences. 2023; 24(13):10596. https://doi.org/10.3390/ijms241310596
Chicago/Turabian StyleGeng, Chenhao, Xinyue Wang, Jiale Chen, Na Sun, Yuru Wang, Zizheng Li, Lu Han, Shike Hou, Haojun Fan, Ning Li, and et al. 2023. "Repetitive Low-Level Blast Exposure via Akt/NF-κB Signaling Pathway Mediates the M1 Polarization of Mouse Alveolar Macrophage MH-S Cells" International Journal of Molecular Sciences 24, no. 13: 10596. https://doi.org/10.3390/ijms241310596