
Circular Intronic RNA circTTN Inhibits Host Gene Transcription and Myogenesis by Recruiting PURB Proteins to form Heterotypic Complexes
 ,
,
Abstract
:1. Introduction
2. Results
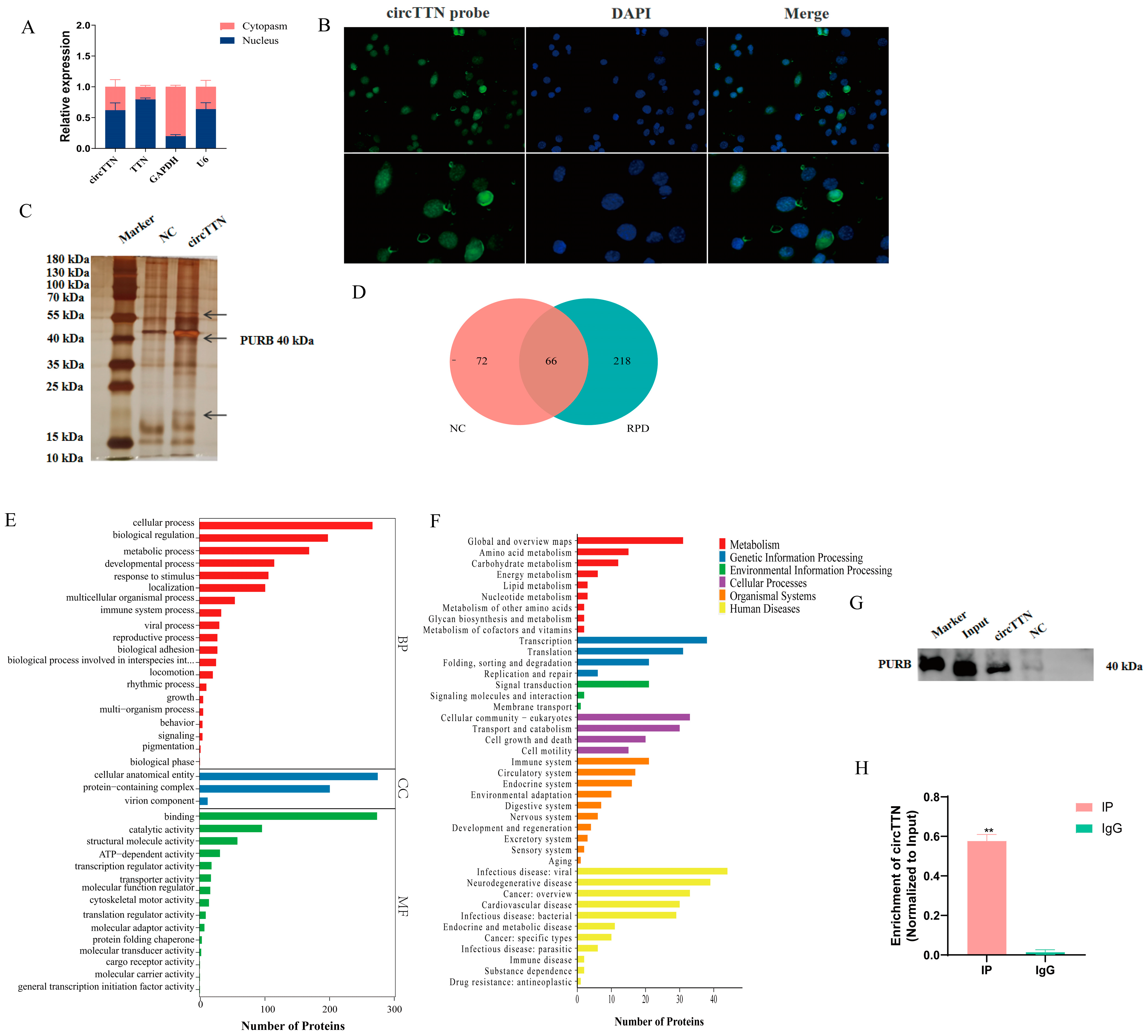
2.1. Characterization Analysis of circTTN
2.2. circTTN Inhibits Proliferation and Differentiation of C2C12 Cells and Host Gene TTN
2.3. circTTN Interacts with PURB Protein
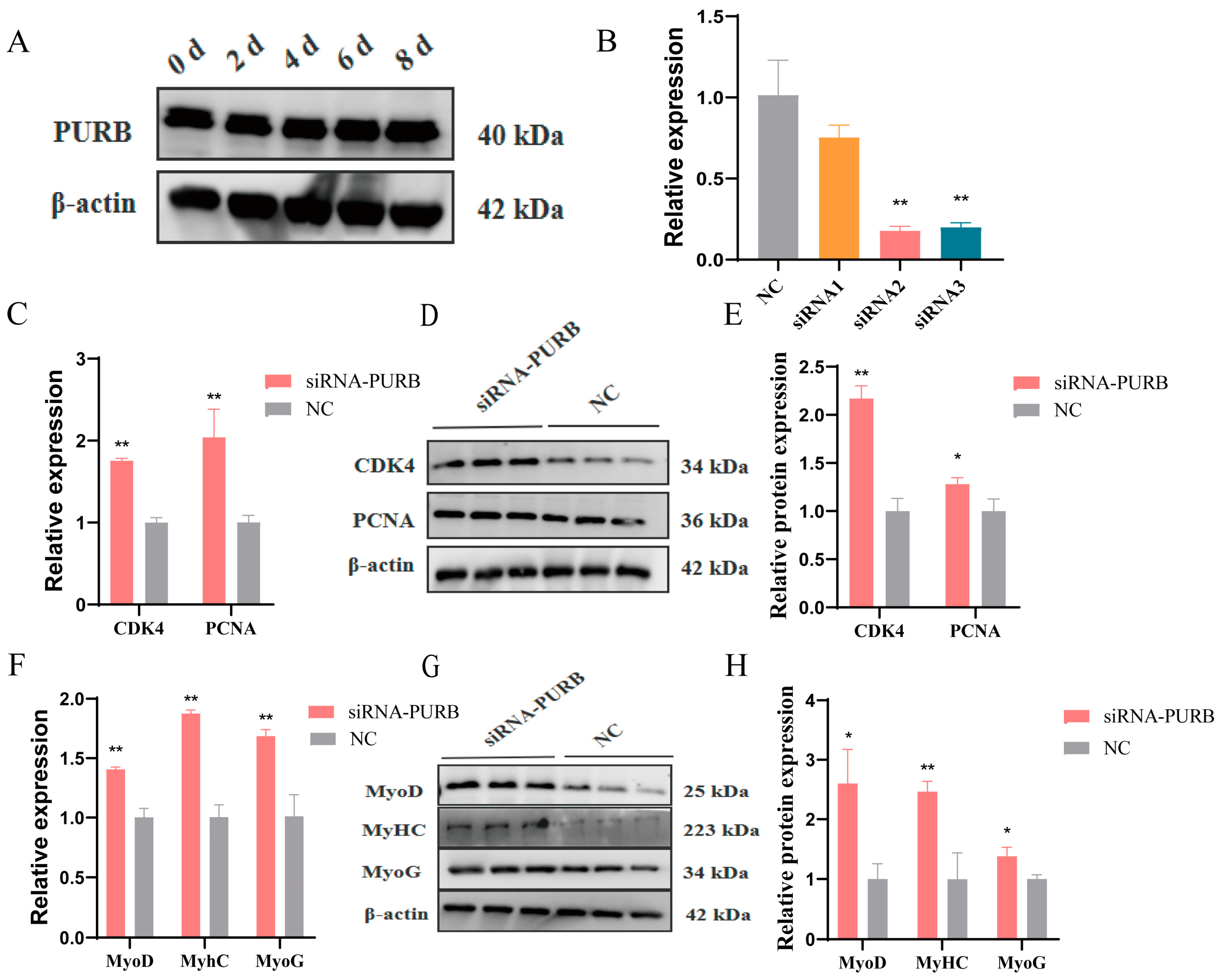
2.4. PURB Knockdown Promotes Proliferation and Differentiation of C2C12 Cells and Increase TTN Expression
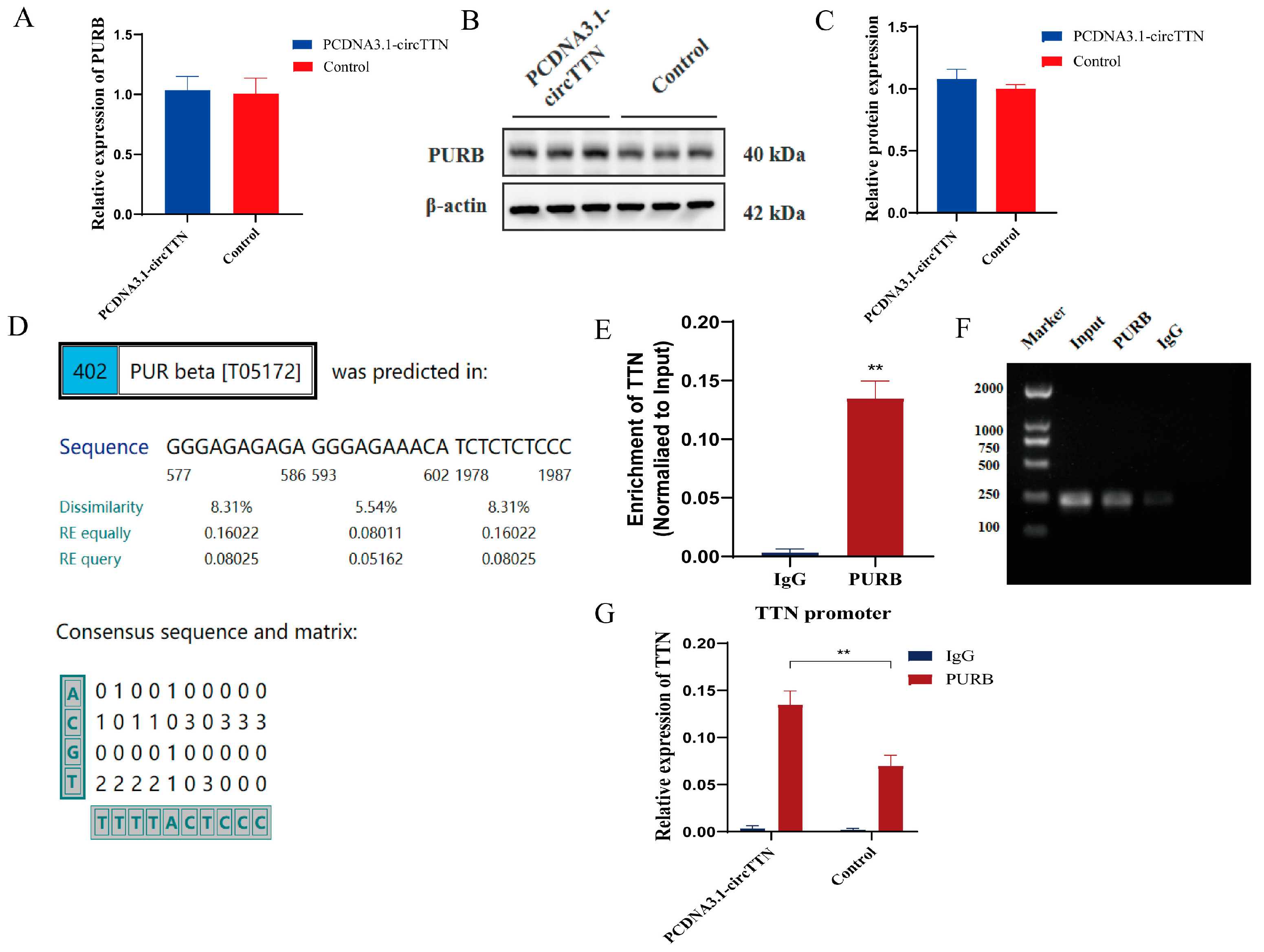
2.5. circTTN Inhibits TTN Transcription by Recruiting PURB
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Samples
4.2. Culture and Differentiation of C2C12 Cells
4.3. RNA Extraction and cDNA Synthesis
4.4. Quantitative Real-Time PCR
4.5. Vector Construction and siRNA
4.6. CCK8 and EdU Assays
4.7. Flow Cytometry
4.8. Immunofluorescence Staining
4.9. Western Blot Analysis
4.10. Nucleoplasmic Separation and RNA Fluorescence In Situ Hybridization
4.11. RNA Pull-Down and Mass Spectrometry
4.12. RNA Immunoprecipitation (RIP) Analysis
4.13. ChIP-qPCR Analysis
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wolfe, R.R. The underappreciated role of muscle in health and disease. Am. J. Clin. Nutr. 2006, 84, 475–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: Molecular regulation of myogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charge, S.B.; Rudnicki, M.A. Cellular and molecular regulation of muscle regeneration. Physiol. Rev. 2004, 84, 209–238. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Hernandez, J.M.; Garcia-Gonzalez, E.G.; Brun, C.E.; Rudnicki, M.A. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. Semin. Cell Dev. Biol. 2017, 72, 10–18. [Google Scholar] [CrossRef]
- Li, H.; Zhu, C.; Tao, Z.; Xu, W.; Song, W.; Hu, Y.; Zhu, W.; Song, C. MyoD and Myf6 gene expression patterns in skeletal muscle during embryonic and posthatch development in the domestic duck (Anas platyrhynchos domestica). J. Anim. Breed. Genet. 2014, 131, 194–201. [Google Scholar] [CrossRef]
- Diener, T.O. Potato spindle tuber “virus”. IV. A replicating, low molecular weight RNA. Virology 1971, 45, 411–428. [Google Scholar] [CrossRef]
- Capel, B.; Swain, A.; Nicolis, S.; Hacker, A.; Walter, M.; Koopman, P.; Goodfellow, P.; Lovell-Badge, R. Circular transcripts of the testis-determining gene Sry in adult mouse testis. Cell 1993, 73, 1019–1030. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Yin, D.; Li, P.; Zhang, Z.; Zhang, X.; Li, H.; Li, R.; Hou, L.; Liu, H.; Wu, W. Profiling and Functional Analysis of Circular RNAs in Porcine Fast and Slow Muscles. Front. Cell Dev. Biol. 2020, 8, 322. [Google Scholar] [CrossRef]
- Wei, X.; Li, H.; Yang, J.; Hao, D.; Dong, D.; Huang, Y.; Lan, X.; Plath, M.; Lei, C.; Lin, F.; et al. Circular RNA profiling reveals an abundant circLMO7 that regulates myoblasts differentiation and survival by sponging miR-378a-3p. Cell Death Dis. 2017, 8, e3153. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.; Zhang, X.; Odame, E.; Xu, X.; Chen, Y.; Ye, J.; Zhou, H.; Dai, D.; Kyei, B.; Zhan, S.; et al. CircRNA-Protein Interactions in Muscle Development and Diseases. Int. J. Mol. Sci. 2021, 22, 3262. [Google Scholar] [CrossRef]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 Is a Circular RNA that Can Be Translated and Functions in Myogenesis. Mol. Cell 2017, 66, 22–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Fang, L. CircRPAP2 regulates the alternative splicing of PTK2 by binding to SRSF1 in breast cancer. Cell Death Discov. 2022, 8, 152. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.R.; Yang, J.H.; Tsitsipatis, D.; Panda, A.C.; Noh, J.H.; Kim, K.M.; Munk, R.; Nicholson, T.; Hanniford, D.; Argibay, D.; et al. circSamd4 represses myogenic transcriptional activity of PUR proteins. Nucleic Acids Res. 2020, 48, 3789–3805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelmohsen, K.; Panda, A.C.; Munk, R.; Grammatikakis, I.; Dudekula, D.B.; De, S.; Kim, J.; Noh, J.H.; Kim, K.M.; Martindale, J.L.; et al. Identification of HuR target circular RNAs uncovers suppression of PABPN1 translation by CircPABPN1. RNA Biol. 2017, 14, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Yang, J.; Yang, J.; He, J.; Gong, Y.; Xiao, Y.; Zeng, Q.; Xu, K.; Duan, Y.; He, J.; et al. Spatiotemporal Regulation and Functional Analysis of Circular RNAs in Skeletal Muscle and Subcutaneous Fat during Pig Growth. Biology 2021, 10, 841. [Google Scholar] [CrossRef]
- Yin, L.; Li, N.; Jia, W.; Wang, N.; Liang, M.; Yang, X.; Du, G. Skeletal muscle atrophy: From mechanisms to treatments. Pharmacol. Res. 2021, 172, 105807. [Google Scholar] [CrossRef]
- Chen, R.; Lei, S.; Jiang, T.; Zeng, J.; Zhou, S.; She, Y. Roles of lncRNAs and circRNAs in regulating skeletal muscle development. Acta Physiol. 2020, 228, e13356. [Google Scholar] [CrossRef]
- Ding, H.; Heng, B.; He, W.; Shi, L.; Lai, C.; Xiao, L.; Ren, H.; Mo, S.; Su, Z. Chronic reactive oxygen species exposure inhibits glucose uptake and causes insulin resistance in C2C12 myotubes. Biochem. Biophys. Res. Commun. 2016, 478, 798–803. [Google Scholar] [CrossRef]
- Wong, C.Y.; Al-Salami, H.; Dass, C.R. C2C12 cell model: Its role in understanding of insulin resistance at the molecular level and pharmaceutical development at the preclinical stage. J. Pharm. Pharmacol. 2020, 72, 1667–1693. [Google Scholar] [CrossRef]
- Sun, D.; An, J.; Cui, Z.; Li, J.; You, Z.; Lu, C.; Yang, Y.; Gao, P.; Guo, X.; Li, B.; et al. CircCSDE1 Regulates Proliferation and Differentiation of C2C12 Myoblasts by Sponging miR-21-3p. Int. J. Mol. Sci. 2022, 23, 12038. [Google Scholar] [CrossRef] [PubMed]
- Patop, I.L.; Wust, S.; Kadener, S. Past, present, and future of circRNAs. EMBO J. 2019, 38, e100836. [Google Scholar] [CrossRef]
- Dhawan, A.; Harris, A.L.; Buffa, F.M.; Scott, J.G. Endogenous miRNA sponges mediate the generation of oscillatory dynamics for a non-coding RNA network. J. Theor. Biol. 2019, 481, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.C. Circular RNAs Act as miRNA Sponges. Adv. Exp. Med. Biol. 2018, 1087, 67–79. [Google Scholar]
- You, X.; Vlatkovic, I.; Babic, A.; Will, T.; Epstein, I.; Tushev, G.; Akbalik, G.; Wang, M.; Glock, C.; Quedenau, C.; et al. Neural circular RNAs are derived from synaptic genes and regulated by development and plasticity. Nat. Neurosci. 2015, 18, 603–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Lei, X. A web server for identifying circRNA-RBP variable-length binding sites based on stacked generalization ensemble deep learning network. Methods 2022, 205, 179–190. [Google Scholar] [CrossRef]
- Yang, Y.; Hou, Z.; Ma, Z.; Li, X.; Wong, K.C. iCircRBP-DHN: Identification of circRNA-RBP interaction sites using deep hierarchical network. Brief. Bioinform. 2021, 22, bbaa27. [Google Scholar] [CrossRef]
- Wang, Z.; Lei, X. Matrix factorization with neural network for predicting circRNA-RBP interactions. BMC Bioinform. 2020, 21, 229. [Google Scholar] [CrossRef]
- Wang, X.; Cao, X.; Dong, D.; Shen, X.; Cheng, J.; Jiang, R.; Yang, Z.; Peng, S.; Huang, Y.; Lan, X. Circular RNA TTN Acts As a miR-432 Sponge to Facilitate Proliferation and Differentiation of Myoblasts via the IGF2/PI3K/AKT Signaling Pathway. Mol. Ther. Nucleic Acids 2019, 18, 966–980. [Google Scholar] [CrossRef] [Green Version]
- Kelm, R.J., Jr.; Wang, S.X.; Polikandriotis, J.A.; Strauch, A.R. Structure/function analysis of mouse Purbeta, a single-stranded DNA-binding repressor of vascular smooth muscle alpha-actin gene transcription. J. Biol. Chem. 2003, 278, 38749–38757. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Tsika, G.L.; Rindt, H.; Schreiber, K.L.; McCarthy, J.J.; Kelm, R.J., Jr.; Tsika, R. Puralpha and Purbeta collaborate with Sp3 to negatively regulate beta-myosin heavy chain gene expression during skeletal muscle inactivity. Mol. Cell. Biol. 2007, 27, 1531–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, E.M. The Pur protein family: Clues to function from recent studies on cancer and AIDS. Anticancer Res. 2003, 23, 2093–2100. [Google Scholar] [PubMed]
- Kelm, R.J., Jr.; Lamba, G.S.; Levis, J.E.; Holmes, C.E. Characterization of purine-rich element binding protein B as a novel biomarker in acute myelogenous leukemia prognostication. J. Cell. Biochem. 2018, 119, 2073–2083. [Google Scholar] [CrossRef] [PubMed]
- Hariharan, S.; Kelm, R.J., Jr.; Strauch, A.R. The Puralpha/Purbeta single-strand DNA-binding proteins attenuate smooth-muscle actin gene transactivation in myofibroblasts. J. Cell. Physiol. 2014, 229, 1256–1271. [Google Scholar] [CrossRef] [PubMed]
- Maamar, H.; Cabili, M.N.; Rinn, J.; Raj, A. linc-HOXA1 is a noncoding RNA that represses Hoxa1 transcription in cis. Genes Dev. 2013, 27, 1260–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Wang, X.; Tan, Y.; Xu, Y.; Guo, X.; Wu, Y.; Wang, W.; Jing, R.; Zhu, F.; Ye, D.; et al. LncCMRR Plays an Important Role in Cardiac Differentiation by Regulating the Purb/Flk1 Axis. Stem Cells 2023, 41, 11–25. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Chen, Y.; Yu, X.; Yu, X.; Wang, S.; Sun, H.; Zheng, X.; Zheng, X.; Zhang, L.; et al. Circ-calm4 regulates hypoxia-induced pulmonary artery smooth muscle autophagy by binding Purb. J. Mol. Cell. Cardiol. 2023, 176, 41–54. [Google Scholar] [CrossRef]
- Li, L.; Chen, Y.; Nie, L.; Ding, X.; Zhang, X.; Zhao, W.; Xu, X.; Kyei, B.; Dai, D.; Zhan, S.; et al. MyoD-induced circular RNA CDR1as promotes myogenic differentiation of skeletal muscle satellite cells. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 807–821. [Google Scholar] [CrossRef]
- Chen, M.; Wei, X.; Song, M.; Jiang, R.; Huang, K.; Deng, Y.; Liu, Q.; Shi, D.; Li, H. Circular RNA circMYBPC1 promotes skeletal muscle differentiation by targeting MyHC. Mol. Ther. Nucleic Acids 2021, 24, 352–368. [Google Scholar] [CrossRef]
- Lv, W.; Zhao, S.; Hou, Y.; Tong, Q.; Peng, Y.; Li, J.; Xu, Z.; Zuo, B. Single Nucleotide Polymorphisms of Porcine lncMGPF Regulate Meat Production Traits by Affecting RNA Stability. Front. Cell Dev. Biol. 2021, 9, 731712. [Google Scholar] [CrossRef]
- Lv, W.; Jin, J.; Xu, Z.; Luo, H.; Guo, Y.; Wang, X.; Wang, S.; Zhang, J.; Zuo, H.; Bai, W.; et al. lncMGPF is a novel positive regulator of muscle growth and regeneration. J. Cachexia Sarcopenia Muscle 2020, 11, 1723–1746. [Google Scholar] [CrossRef] [PubMed]
- Chauveau, C.; Rowell, J.; Ferreiro, A. A rising titan: TTN review and mutation update. Hum. Mutat. 2014, 35, 1046–1059. [Google Scholar] [CrossRef] [PubMed]
- Gautel, M.; Djinovic-Carugo, K. The sarcomeric cytoskeleton: From molecules to motion. J. Exp. Biol. 2016, 219, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accessions | Names | Unused | Description |
|---|---|---|---|
| Q6QAQ1 | ACTB | 191.97 | Actin, cytoplasmic 1 |
| P02543 | VIM | 149.62 | Vimentin |
| A0A287AU59 | ACACA | 133.63 | Acetyl-CoA carboxylase 1 |
| A0A287BNK7 | PLEC | 133 | Uncharacterized protein |
| A0A287BHM1 | TPM1 | 91.59 | Tropomyosin alpha-1 chain |
| A0A5G2RI50 | MYO1C | 83.87 | Unconventional myosin-Ic isoform b |
| Q2XVP4 | TUBA1B | 61.55 | Tubulin alpha-1B chain |
| P02554 | Tubulin beta | 60.54 | Tubulin beta chain |
| F6Q364 | ACTA2 | 59.23 | Uncharacterized protein |
| P02540 | DES | 52.77 | Desmin |
| A0A287BQP1 | RBM25 | 36.79 | Uncharacterized protein |
| A0A288CG57 | EEF1A1 | 36.1 | Elongation factor 1-alpha |
| A0A286ZVS0 | SF3B3 | 35.56 | Splicing factor 3B subunit 3 |
| F1SSA6 | MYH10 | 33.34 | Myosin motor domain-containing protein |
| A0A287BSP1 | MYO5A | 32.65 | Uncharacterized protein |
| A0A287A4H2 | PURB | 32.47 | Transcriptional activator protein Pur-beta |
| P80021 | ATPATP5F1A | 31.56 | ATP synthase subunit alpha, mitochondrial |
| F1RUL1 | RBM17 | 29.87 | Splicing factor 45 |
| A0A287B1F6 | MPRIP | 29.21 | Uncharacterized protein |
| K9J4V0 | SNRNP200 | 29 | Protogenin isoform X1 |
| I3L920 | CORO1C | 27.2 | Coronin |
| I3L650 | CALD1 | 26.54 | Uncharacterized protein |
| A0A287BIL8 | GN = HSPA5 | 26.3 | 78 kDa glucose-regulated protein |
| K7GLT8 | ATP5F1B | 24.2 | ATP synthase subunit beta |
| A0A5G2R8T3 | MYO1E | 23.9 | Uncharacterized protein |
| A0A286ZHW0 | SF3B1 | 21.89 | Splicing factor 3B subunit 1 isoform 1 |
| F1S5A8 | DHX15 | 21.49 | RNA helicase |
| A0A286ZWK2 | HSPA8 | 20.98 | Uncharacterized protein |
| I3LLT2 | CAD | 20.86 | Aspartate carbamoyltransferase |
| A0A286ZXU2 | FLNA | 19.38 | Uncharacterized protein |
| Genes | Primer (5′→3′) | Length |
|---|---|---|
| circTTN (convergent primer) | F: TGTCTCACTGCCTTGTCTGATG | 241 bp |
| R: TTAGCAGCTGGTTCAGTCACC | ||
| circTTN (divergent primer) | F: AGCTGCTAAAGTGCCCATTCC | 139 bp |
| R: AAACAAAGAACATCAGACAAGGCA | ||
| β-actin | F: GGACTGTTACTGAGCTGCGTT | 290 bp |
| R: CGCCTTCACCGTTCCAGTT | ||
| CDK4 | F: CGAGCGTAAGGCTGATGGAT | 177 bp |
| R: CCAGGCCGCTTAGAAACTGA | ||
| PCNA | F: GCCGAGACCTTAGCCACATT | 229 bp |
| R: GTAGGAGACAGTGGAGTGGC | ||
| CCND | F: TCAAGTGTGACCCGGACTG | 235 bp |
| R: GCTCCTTCCTCTTTGCGGG | ||
| MyoD | F: AGTGAATGAGGCCTTCGAGA | 169 bp |
| R: GCATCTGAGTCGCCACTGTA | ||
| MyoG | F: CAATGCACTGGAGTTCGGT | 134 bp |
| R: CTGGGAAGGCAACAGACAT | ||
| MyHC | F: CGGTCGAAGTTGCATCCCT | 141 bp |
| R: GAGCCTCGATTCGCTCCTTT | ||
| TTN | F: GTCCTCCATCTCCTCCTGGT | 173 bp |
| R: GCGTTTTGTTGACCCTCACC | ||
| PURB | F: GGAGCGACAGAGGGATAAGC | 181 bp |
| R: TAGCTCACTGGGGACGAGAA | ||
| TTN-promoter | F: TGCCCACAAGTGTCTTCAACT | 179 bp |
| R: ACCTGTTCCTCTGATTGGCAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ai, N.; Yu, Z.; Xu, X.; Liufu, S.; Wang, K.; Huang, S.; Li, X.; Liu, X.; Chen, B.; Ma, H.; et al. Circular Intronic RNA circTTN Inhibits Host Gene Transcription and Myogenesis by Recruiting PURB Proteins to form Heterotypic Complexes. Int. J. Mol. Sci. 2023, 24, 9859. https://doi.org/10.3390/ijms24129859
Ai N, Yu Z, Xu X, Liufu S, Wang K, Huang S, Li X, Liu X, Chen B, Ma H, et al. Circular Intronic RNA circTTN Inhibits Host Gene Transcription and Myogenesis by Recruiting PURB Proteins to form Heterotypic Complexes. International Journal of Molecular Sciences. 2023; 24(12):9859. https://doi.org/10.3390/ijms24129859
Chicago/Turabian StyleAi, Nini, Zonggang Yu, Xueli Xu, Sui Liufu, Kaiming Wang, Shengqiang Huang, Xintong Li, Xiaolin Liu, Bohe Chen, Haiming Ma, and et al. 2023. "Circular Intronic RNA circTTN Inhibits Host Gene Transcription and Myogenesis by Recruiting PURB Proteins to form Heterotypic Complexes" International Journal of Molecular Sciences 24, no. 12: 9859. https://doi.org/10.3390/ijms24129859