
Natural and Pathological Autoantibodies Show Age-Related Changes in a Spontaneous Autoimmune Mouse (NZB) Model

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Pathological Autoantibody Levels Correlate with the Development of Autoimmune Hemolytic Anemia
2.2. Age-Dependent Changes in the Natural Autoantibody Levels of NZB Mice
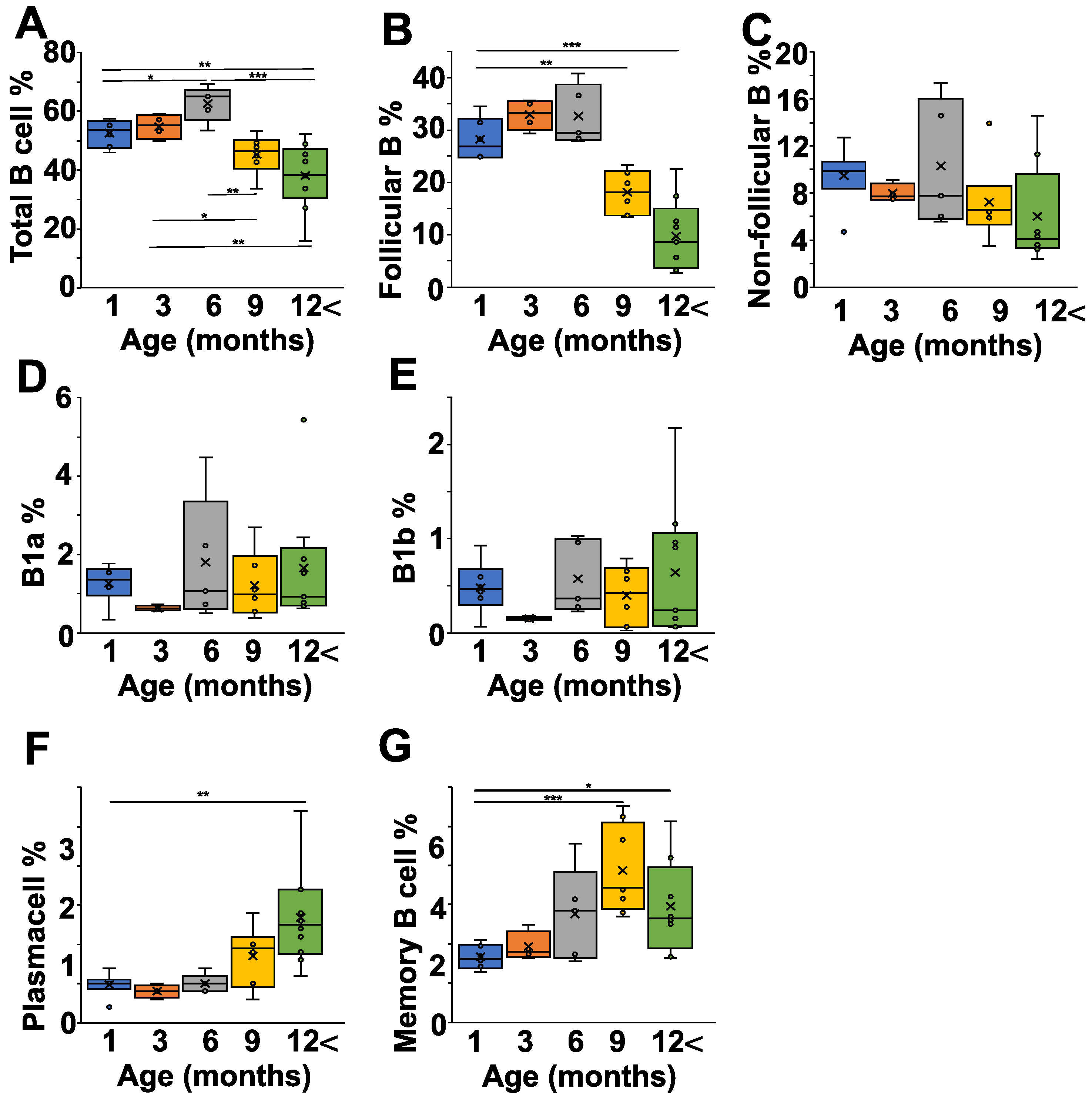
2.3. Age-Dependent Changes of B Cell Composition in the Spleen of NZB Mice
2.4. Age-Dependent Changes of B Cell Composition in the Peritoneum of NZB Mice
3. Discussion
4. Materials and Methods
4.1. Mice and Collection of the Sera
4.2. Antibodies and Reagents
4.3. Coombs Test
4.4. ELISA
4.5. Flow Cytometry
4.6. Statistical Analyisis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abbas, A.; Lichtman, A.; Pillai, S. Cellular and Molecular Immunology, 6th ed.; Saunders: Philadelphia, PA, USA, 2010; p. 3. [Google Scholar]
- Cohen, I.R. The cognitive paradigm and the immunological homunculus. Immunol. Today 1992, 13, 490–494. [Google Scholar] [CrossRef]
- Lacroix-Desmazes, S.; Kaveri, S.V.; Mouthon, L.; Ayouba, A.; Malanchère, E.; Coutinho, A.; Kazatchkine, M.D. Self-reactive antibodies (natural autoantibodies) in healthy individuals. J. Immunol. Methods 1998, 216, 117–137. [Google Scholar] [CrossRef]
- Cohen, I.R.; Young, D.B. Autoimmunity, microbial immunity and the immunological homunculus. Immunol. Today 1991, 12, 105–110. [Google Scholar] [CrossRef]
- Lee, B.H.; Gauna, A.E.; Pauley, K.M.; Park, Y.J.; Cha, S. Animal models in autoimmune diseases: Lessons learned from mouse models for Sjögren’s Syndrome. Clin. Rev. Allergy Immunol. 2012, 42, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Petersen, F. A methodological review of induced animal models of autoimmune diseases. Autoimmun. Rev. 2018, 17, 473–479. [Google Scholar] [CrossRef]
- Howie, H.L.; Hudson, K.E. Murine models of autoimmune hemolytic anemia. Curr. Opin. Hematol. 2018, 25, 473–481. [Google Scholar] [CrossRef]
- Karnopp, T.E.; Chapacais, G.F.; Freitas, E.C.; Monticielo, O.A. Lupus animal models and neuropsychiatric implications. Clin. Rheumatol. 2021, 40, 2535–2545. [Google Scholar] [CrossRef]
- Feige, U.; van Eden, W. Infection, autoimmunity and autoimmune disease. Stress-Inducible Cell. Responses 1996, 77, 359–373. [Google Scholar]
- Hirose, S.; Kinoshita, K.; Nozawa, S.; Nishimura, H.; Shirai, T. Effects of major histocompatibility complex on autoimmune disease of H-2-congenic New Zealand mice. Int. Immunol. 1990, 2, 1091–1095. [Google Scholar] [CrossRef]
- Radons, J. The human HSP70 family of chaperones: Where do we stand? Cell Stress Chaperones 2016, 21, 379–404. [Google Scholar] [CrossRef] [Green Version]
- Czömpöly, T.; Olasz, K.; Simon, D.; Nyárády, Z.; Pálinkás, L.; Czirják, L.; Berki, T.; Németh, P. A possible new bridge between innate and adaptive immunity: Are the anti-mitochondrial citrate synthase autoantibodies components of the natural antibody network? Mol. Immunol. 2006, 43, 1761–1768. [Google Scholar] [CrossRef]
- Kappler, K.; Hennet, T. Emergence and significance of carbohydrate-specific antibodies. Genes Immun. 2020, 21, 224–239. [Google Scholar] [CrossRef] [PubMed]
- Schwartz-Albiez, R.; Monteiro, R.C.; Rodriguez, M.; Binder, C.J.; Shoenfeld, Y. Natural antibodies, intravenous immunoglobulin and their role in autoimmunity, cancer and inflammation. Clin. Exp. Immunol. 2009, 158, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Palma, J.; Tokarz-Deptuła, B.; Deptuła, J.; Deptuła, W. Natural antibodies—Facts known and unknown. Cent. Eur. J. Immunol. 2018, 43, 466–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyneveld, G.I.J.; Savelkoul, H.F.J.; Parmentier, H.K. Current Understanding of Natural Antibodies and Exploring the Possibilities of Modulation Using Veterinary Models. A Review. Front. Immunol. 2020, 11, 2139. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Ng, S.M.; Hassouna, E.; Warrington, A.; Oh, S.H.; Rodriguez, M. Human-derived natural antibodies: Biomarkers and potential therapeutics. Future Neurol. 2015, 10, 25–39. [Google Scholar] [CrossRef] [Green Version]
- Lobo, P.I. Role of natural autoantibodies and natural IgM anti-leucocyte autoantibodies in health and disease. Front. Immunol. 2016, 7, 198. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.; Ding, J.L. Natural Antibodies Bridge Innate and Adaptive Immunity. J. Immunol. 2015, 194, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Notley, C.A.; Baker, N.; Ehrenstein, M.R. Secreted IgM Enhances B Cell Receptor Signaling and Promotes Splenic but Impairs Peritoneal B Cell Survival. J. Immunol. 2010, 184, 3386–3393. [Google Scholar] [CrossRef] [Green Version]
- Ehrenstein, B.M.R.; Cook, H.T.; Neuberger, M.S. Deficiency in Serum Immunoglobulin (Ig) M Predisposes to Development of IgG Autoantibodies. J. Exp. Med. 2000, 191, 1253–1257. [Google Scholar] [CrossRef] [Green Version]
- Boes, M.; Schmidt, T.; Linkemann, K.; Beaudette, B.C.; Marshak-rothstein, A.; Chen, J. Accelerated development of IgG autoantibodies and autoimmune disease in the absence of secreted IgM. Proc. Natl. Acad. Sci. USA 2000, 97, 1184–1189. [Google Scholar] [CrossRef] [Green Version]
- Merbl, Y.; Zucker-Toledano, M.; Quintana, F.J.; Cohen, I.R. Newborn humans manifest autoantibodies to defined self molecules detected by antigen microarray informatics. J. Clin. Investig. 2007, 117, 712–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elias, D.; Reshef, T.; Birk, O.S.; Van Der Zee, R.; Walker, M.D.; Cohen, I.R. Vaccination against autoimmune mouse diabetes with a T-cell epitope of the human 65-kDa heat shock protein. Proc. Natl. Acad. Sci. USA 1991, 88, 3088–3091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintana, F.J.; Carmi, P.; Mor, F.; Cohen, I.R. Inhibition of adjuvant-induced arthritis by DNA vaccination with the 70-kd or the 90-kd human heat-shock protein: Immune cross-regulation with the 60-kd heat-shock protein. Arthritis Rheum. 2004, 50, 3712–3720. [Google Scholar] [CrossRef] [PubMed]
- Almanzar, G.; Öllinger, R.; Leuenberger, J.; Onestingel, E.; Rantner, B.; Zehm, S.; Cardini, B.; van der Zee, R.; Grundtman, C.; Wick, G. Autoreactive HSP60 epitope-specific T-cells in early human atherosclerotic lesions. J. Autoimmun. 2012, 39, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Cohen, I.R. Autoantibody repertoires, natural biomarkers, and system controllers. Trends Immunol. 2013, 34, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Czömpöly, T.; Olasz, K.; Nyárády, Z.; Simon, D.; Bovári, J.; Németh, P. Detailed analyses of antibodies recognizing mitochondrial antigens suggest similar or identical mechanism for production of natural antibodies and natural autoantibodies. Autoimmun. Rev. 2008, 7, 463–467. [Google Scholar] [CrossRef]
- Merashli, M.; Arcaro, A.; Graf, M.; Gentile, F.; Ames, P.R.J. Autoimmune haemolytic anaemia and antiphospholipid antibodies in paediatrics: A systematic review and meta-analysis. Clin. Rheumatol. 2021, 40, 1967–1973. [Google Scholar] [CrossRef]
- Barcellini, W.; Fattizzo, B. The Changing Landscape of Autoimmune Hemolytic Anemia. Front. Immunol. 2020, 11, 946. [Google Scholar] [CrossRef]
- Helyer, B.J.; Howie, J.B. Spontaneous Auto-Immune Disease in NZB/BL Mice. Br. J. Haematol. 1963, 9, 119–131. [Google Scholar] [CrossRef]
- Hentati, B.; Brogard, B.P.; Jouanne, C.; Avrameas, S.; Ternynck, T. Natural Autoantibodies are Involved in the Haemolytic Anaemia of NZB Mice. J. Autoimmun. 1994, 7, 425–439. [Google Scholar] [CrossRef] [PubMed]
- Tarjanyi, O.; Boldizsar, F.; Nemeth, P.; Mikecz, K.; Glant, T.T. Age-related changes in arthritis susceptibility and severity in a murine model of rheumatoid arthritis. Immun. Ageing 2009, 6, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolouri, N.; Akhtari, M.; Farhadi, E.; Mansouri, R.; Faezi, S.T.; Jamshidi, A.; Mahmoudi, M. Role of the innate and adaptive immune responses in the pathogenesis of systemic lupus erythematosus. Inflamm. Res. 2022, 71, 537–554. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.M.; Tsokos, G.C. Mitochondria in the Pathogenesis of Systemic Lupus Erythematosus. Curr. Rheumatol. Rep. 2022, 24, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Böröcz, K.; Simon, D.; Erdő-Bonyár, S.; Kovács, K.T.; Tuba; Czirják, L.; Németh, P.; Berki, T. Relationship between natural and infection-induced antibodies in systemic autoimmune diseases (SAD): SLE, SSc and RA. Clin. Exp. Immunol. 2021, 203, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Andrews, B.S.; Eisenberg, R.A.; Theofilopoulos, A.N.; Izui, S.; Wilson, C.B.; McConahey, P.J.; Murphy, E.D.; Roths, J.B.; Dixon, F.J. Spontaneous murine lupus-like syndromes. Clinical and immunopathological manifestations in several strains. J. Exp. Med. 1978, 148, 1198–1215. [Google Scholar] [CrossRef] [Green Version]
- Khanfar, E.; Olasz, K.; Gajdócsi, E.; Jia, X.; Berki, T.; Balogh, P.; Boldizsár, F. Splenectomy modulates the immune response but does not prevent joint inflammation in a mouse model of RA. Clin. Exp. Immunol. 2022, 209, 201–214. [Google Scholar] [CrossRef]
- Coombs, R.R.A.; Mourant, A.E.; Race, R.R. A new test for the detection of weak and incomplete Rh agglutinins. Br. J. Exp. Pathol. 1945, 26, 255–266. [Google Scholar]
- Böröcz, K.; Kinyó, Á.; Simon, D.; Erdő-Bonyár, S.; Németh, P.; Berki, T. Complexity of the Immune Response Elicited by Different COVID-19 Vaccines, in the Light of Natural Autoantibodies and Immunomodulatory Therapies. Int. J. Mol. Sci. 2023, 24, 6439. [Google Scholar] [CrossRef]





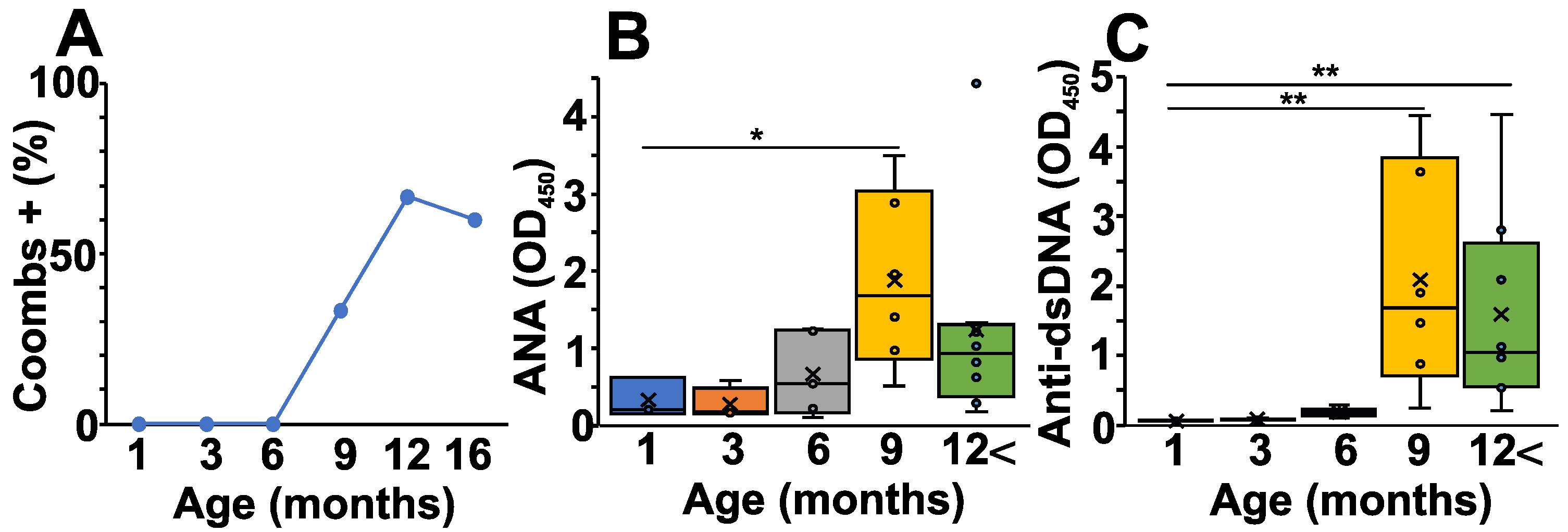{kind=link}
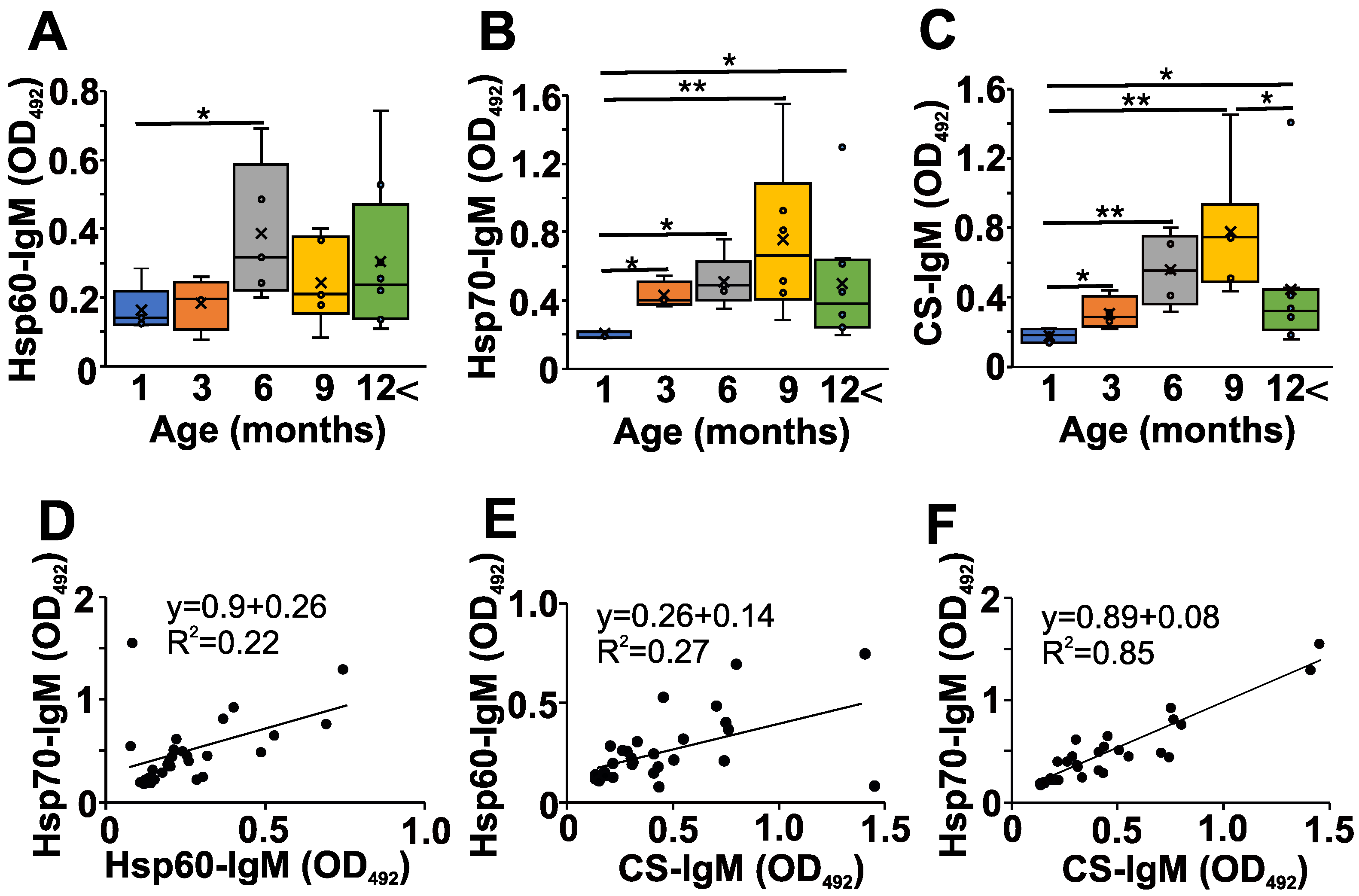{kind=link}
{kind=link}
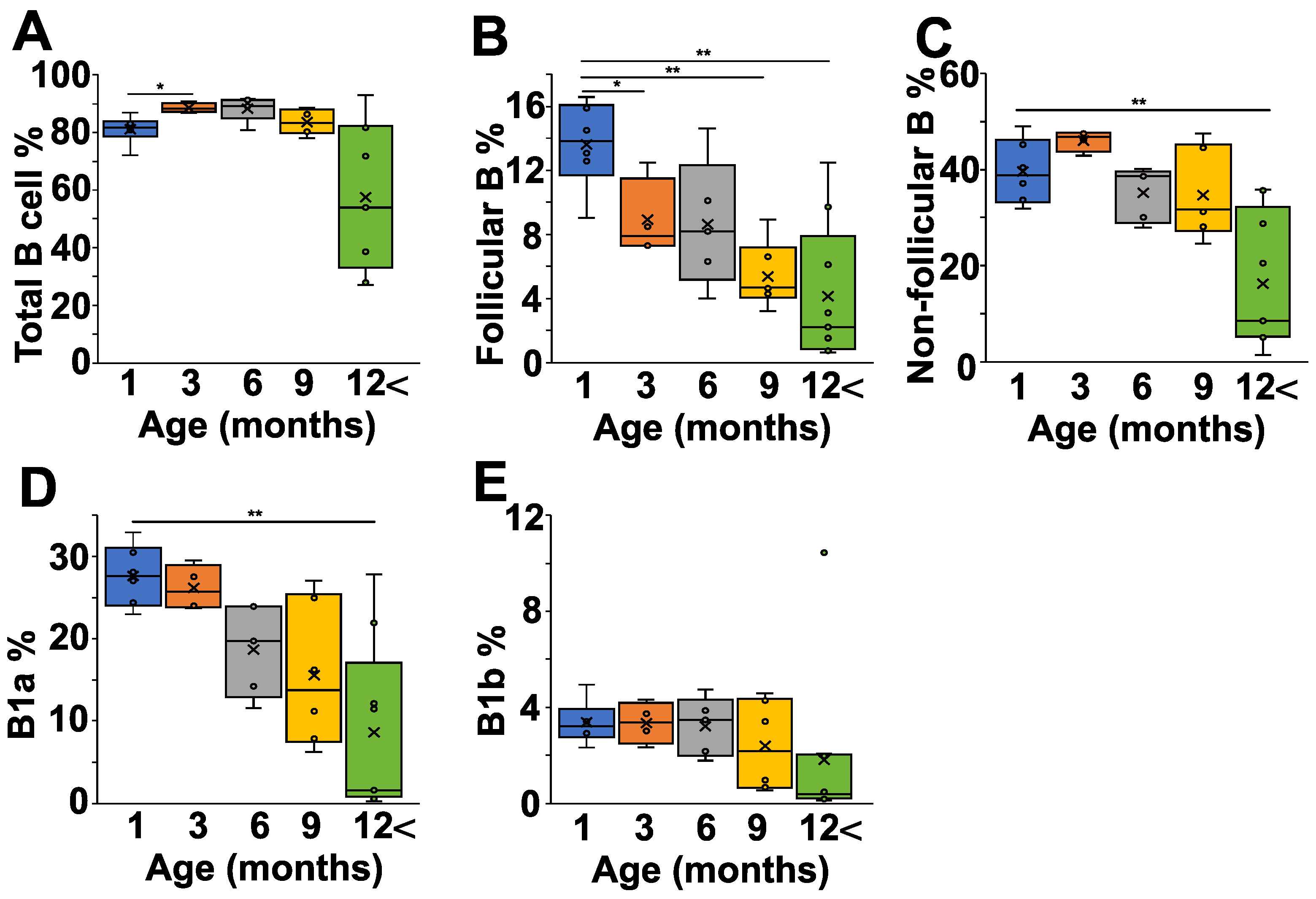{kind=link}
| Hsp60 | Hsp70 | CS | |
|---|---|---|---|
| Hsp60 | 1 | 0.465 * | 0.521 ** |
| Hsp70 | 0.465 * | 1 | 0.923 ** |
| CS | 0.521 ** | 0.923 ** | 1 |
| Hsp60 | Hsp70 | CS | ANA | Anti-dsDNA | Coombs | |
|---|---|---|---|---|---|---|
| ANA | 0.466 * | 0.563 * | 0.667 ** | 1 | 0.689 ** | 0.125 |
| anti-dsDNA | 0.226 | 0.281 | 0.472 * | 0.689 ** | 1 | 0.151 |
| Coombs | −0.012 | 0.205 | 0.151 | 0.125 | 0.151 | 1 |
| Age Range 1 | Average Age 1 | Number |
|---|---|---|
| 1–1.1 | 1.04 | 5 |
| 3–3.3 | 3.2 | 4 |
| 6–6.7 | 6.14 | 5 |
| 8.6–9.4 | 9 | 6 |
| 11.6–17.2 | 14 | 8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gál, S.; Gajdócsi, E.; Khanfar, E.; Olasz, K.; Simon, D.; Balogh, P.; Berki, T.; Németh, P.; Boldizsár, F. Natural and Pathological Autoantibodies Show Age-Related Changes in a Spontaneous Autoimmune Mouse (NZB) Model. Int. J. Mol. Sci. 2023, 24, 9809. https://doi.org/10.3390/ijms24129809
Gál S, Gajdócsi E, Khanfar E, Olasz K, Simon D, Balogh P, Berki T, Németh P, Boldizsár F. Natural and Pathological Autoantibodies Show Age-Related Changes in a Spontaneous Autoimmune Mouse (NZB) Model. International Journal of Molecular Sciences. 2023; 24(12):9809. https://doi.org/10.3390/ijms24129809
Chicago/Turabian StyleGál, Szonja, Erzsébet Gajdócsi, Esam Khanfar, Katalin Olasz, Diána Simon, Péter Balogh, Tímea Berki, Péter Németh, and Ferenc Boldizsár. 2023. "Natural and Pathological Autoantibodies Show Age-Related Changes in a Spontaneous Autoimmune Mouse (NZB) Model" International Journal of Molecular Sciences 24, no. 12: 9809. https://doi.org/10.3390/ijms24129809