
Entering, Linked with the Sphinx: Lysophosphatidic Acids Everywhere, All at Once, in the Oral System and Cancer
{kind=link}
{kind=link}
{kind=link}
Abstract
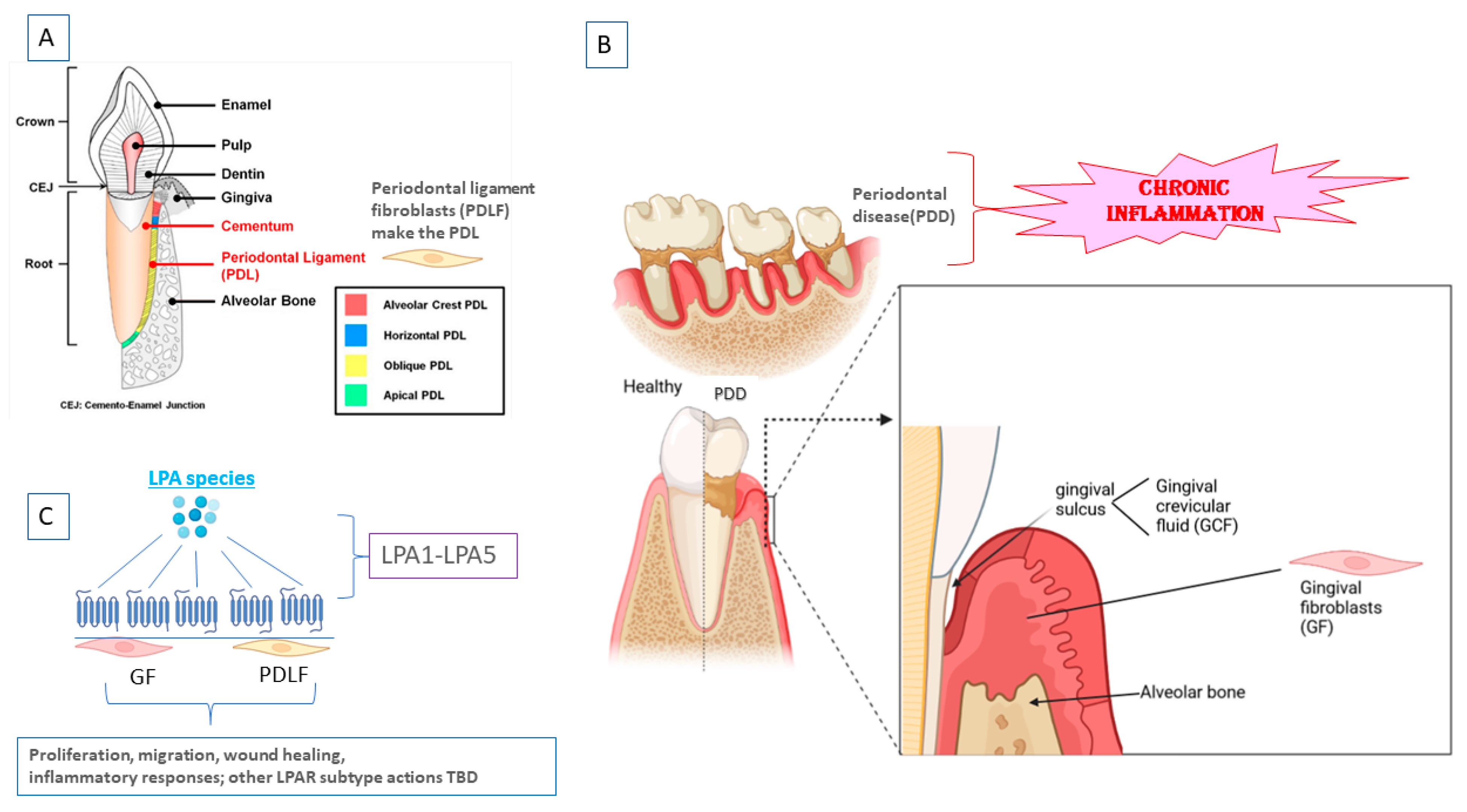
:1. Introduction
1.1. LPA/LPARs
1.2. Autotaxin
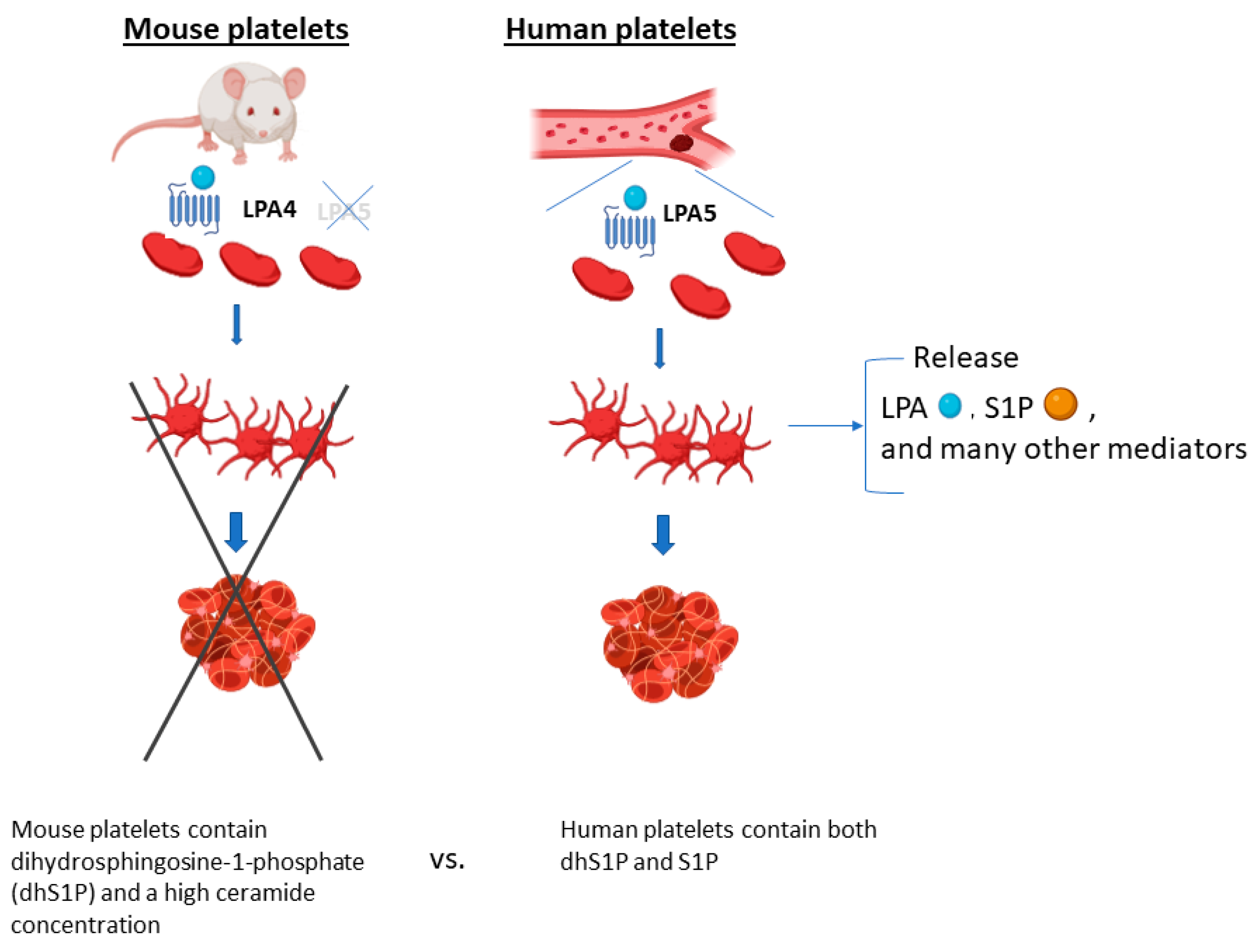
1.3. Sphingosine-1-Phosphate (S1P)
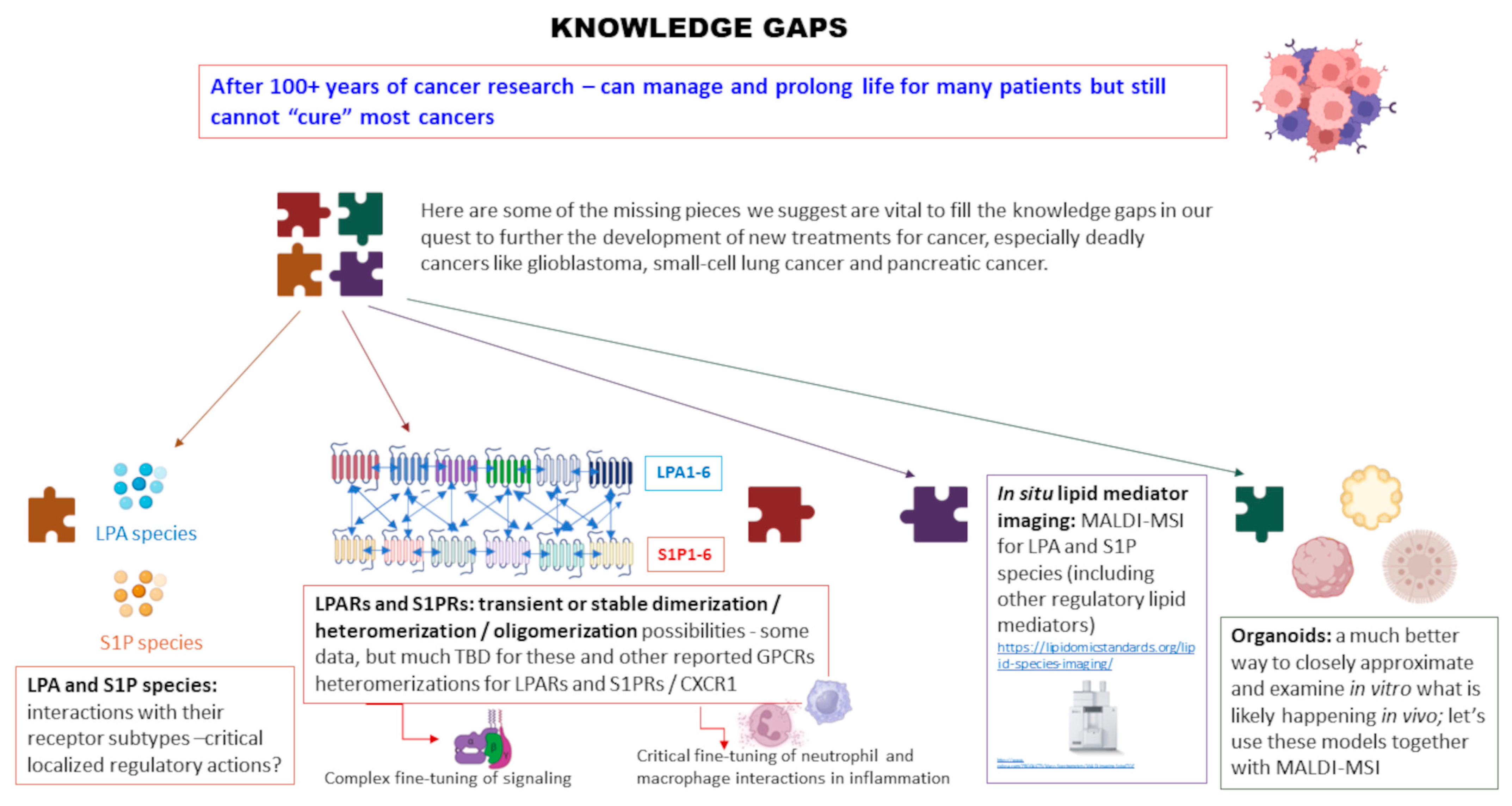
2. Our Perspective: Major Knowledge Gaps in the LPL Field for Investigation
2.1. LPA and S1P Species
2.2. In Situ Lipid Mediator Imaging
2.3. GPCR Multimerization and Signaling Implications
2.4. Organoid Models
3. Major Pieces of a Vast and Complex Puzzle: Conclusions from Our Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gude, D.; Koduganti, R.R.; Prasanna, S.J.; Pothini, L.R. Mouth: A portal to the body. Dent. Res. J. 2012, 9, 659–664. [Google Scholar]
- Weintraub, J.A. The Oral Health in America Report: A Public Health Research Perspective. Prev. Chronic. Dis. 2022, 19, 220067. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.A.; Steiner, B. Oral Health Integration: A Call to Action. Fam. Med. 2018, 50, 637–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silk, H.; Savageau, J.A.; Sullivan, K.; Sawosik, G.; Wang, M. An UpDate of Oral Health Curricula in US Family Medicine Residency Programs. Fam. Med. 2018, 50, 437–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- US Department of Health and Human Services. Oral Health in America: A Report of the Surgeon General; U.S. Department of Health and Human Services, National Institute of Dental and Craniofacial Research, National Institutes of Health: Rockville, MD, USA, 2000. Available online: https://www.nidcr.nih.gov/research/data-statistics/surgeon-general (accessed on 3 May 2023).
- Capodiferro, S.; Limongelli, L.; Favia, G. Oral and Maxillo-Facial Manifestations of Systemic Diseases: An Overview. Medicina 2021, 57, 271. [Google Scholar] [CrossRef]
- Jin, L.J.; Lamster, I.B.; Greenspan, J.S.; Pitts, N.B.; Scully, C.; Warnakulasuriya, S. Global burden of oral diseases: Emerging concepts, management and interplay with systemic health. Oral Dis. 2016, 22, 609–619. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Chavakis, T. Local and systemic mechanisms linking periodontal disease and inflammatory comorbidities. Nat. Rev. Immunol. 2021, 21, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Preshaw, P.M.; Taylor, J.J.; Jaedicke, K.M.; De Jager, M.; Bikker, J.W.; Selten, W.; Bissett, S.M.; Whall, K.M.; van de Merwe, R.; Areibi, A.; et al. Treatment of periodontitis reduces systemic inflammation in type 2 diabetes. J. Clin. Periodontol. 2020, 47, 737–746. [Google Scholar] [CrossRef]
- Michaud, D.S.; Lu, J.; Peacock-Villada, A.Y.; Barber, J.R.; Joshu, C.E.; Prizment, A.E.; Beck, J.D.; Offenbacher, S.; Platz, E.A. Periodontal Disease Assessed Using Clinical Dental Measurements and Cancer Risk in the ARIC Study. J. Natl. Cancer Inst. 2018, 110, 843–854. [Google Scholar] [CrossRef] [Green Version]
- Michaud, D.S.; Kelsey, K.T.; Papathanasiou, E.; Genco, C.A.; Giovannucci, E. Periodontal disease and risk of all cancers among male never smokers: An uPDDated analysis of the Health Professionals Follow-up Study. Ann. Oncol. 2016, 27, 941–947. [Google Scholar] [CrossRef]
- Kim, I.S.; Yang, W.S.; Kim, C.H. Physiological Properties, Functions, and Trends in the Matrix Metalloproteinase Inhibitors in Inflammation-Mediated Human Diseases. Curr. Med. Chem. 2023, 30, 2075–2112. [Google Scholar] [CrossRef] [PubMed]
- Fields, G.B. The Rebirth of Matrix Metalloproteinase Inhibitors: Moving Beyond the Dogma. Cells 2019, 8, 984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, K.A.; Shingala, J.; Evens, A.; Birmann, B.M.; Giovannucci, E.; Michaud, D.S. Periodontal disease and risk of non-Hodgkin lymphoma in the Health Professionals Follow-Up Study. Int. J. Cancer 2017, 140, 1020–1026. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, N.; Ishii, S.; Tsujiuchi, T.; Kagawa, N.; Katoh, K. Comparative analyses of lysophosphatidic acid receptor-mediated signaling. Cell. Mol. Life Sci. 2015, 72, 2377–2394. [Google Scholar] [CrossRef] [PubMed]
- Knowlden, S.; Georas, S.N. The autotaxin-LPA axis emerges as a novel regulator of lymphocyte homing and inflammation. J. Immunol. 2014, 192, 851–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shea, B.S.; Tager, A.M. Role of the lysophospholipid mediators lysophosphatidic acid and sphingosine 1-phosphate in lung fibrosis. Proc. Am. Thorac. Soc. 2012, 9, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Binder, B.Y.; Williams, P.A.; Silva, E.A.; Leach, J.K. Lysophosphatidic Acid and Sphingosine-1-Phosphate: A Concise Review of Biological Function and Applications for Tissue Engineering. Tissue Eng. Part B Rev. 2015, 21, 531–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.H. Biomaterial-Based Approaches for Regeneration of Periodontal Ligament and Cementum Using 3D Platforms. Int. J. Mol. Sci. 2019, 20, 4364. [Google Scholar] [CrossRef] [Green Version]
- Hines, O.J.; Ryder, N.; Chu, J.; McFadden, D. Lysophosphatidic acid stimulates intestinal restitution via cytoskeletal activation and remodeling. J. Surg. Res. 2000, 92, 23–28. [Google Scholar] [CrossRef]
- Ridley, A.J.; Hall, A. The small GTP-binding protein rho regulates the assembly of focal adhesions and actin stress fibers in response to growth factors. Cell 1992, 70, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Ridley, A.J.; Hall, A. Signal transduction pathways regulating Rho-mediated stress fibre formation: Requirement for a tyrosine kinase. EMBO J. 1994, 13, 2600–2610. [Google Scholar] [CrossRef]
- Tomar, A.; George, S.P.; Mathew, S.; Khurana, S. Differential effects of lysophosphatidic acid and phosphatidylinositol 4,5-bisphosphate on actin dynamics by direct association with the actin-binding protein villin. J. Biol. Chem. 2009, 284, 35278–35282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittmar, T.; Hass, R. Intrinsic signalling factors associated with cancer cell-cell fusion. Cell Commun. Signal. 2023, 21, 68. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.P.; Massagué, J. Cancer metastasis: Building a framework. Cell 2006, 127, 679–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melzer, C.; von der Ohe, J.; Hass, R. Enhanced metastatic capacity of breast cancer cells after interaction and hybrid formation with mesenchymal stroma/stem cells (MSC). Cell Commun. Signal. 2018, 16, 2. [Google Scholar] [CrossRef] [Green Version]
- Melzer, C.; Ohe, J.V.; Hass, R. Altered tumor plasticity after different cancer cell fusions with MSC. Int. J. Mol. Sci. 2020, 21, 8347. [Google Scholar] [CrossRef]
- Melzer, C.; von der Ohe, J.; Hass, R. Involvement of Actin Cytoskeletal Components in Breast Cancer Cell Fusion with Human Mesenchymal Stroma/Stem-Like Cells. Int. J. Mol. Sci. 2019, 20, 876. [Google Scholar] [CrossRef] [Green Version]
- Hass, R. Role of MSC in the Tumor Microenvironment. Cancers 2020, 12, 2107. [Google Scholar] [CrossRef]
- Cerutis, D.R.; Dreyer, A.; Cordini, F.; McVaney, T.P.; Mattson, J.S.; Parrish, L.C.; Romito, L.; Huebner, G.R.; Jabro, M. Lysophosphatidic acid modulates the regenerative responses of human gingival fibroblasts and enhances the actions of platelet-derived growth factor. J. Periodontol. 2004, 75, 297–305, Erratum in J. Periodontol. 2004, 75, 1437–1438. [Google Scholar] [CrossRef]
- Cerutis, D.R.; Dreyer, A.C.; Vierra, M.J.; King, J.P.; Wagner, D.J.; Fimple, J.L.; Cordini, F.; McVaney, T.P.; Parrish, L.C.; Wilwerding, T.M.; et al. Lysophosphatidic acid modulates the healing responses of human periodontal ligament fibroblasts and enhances the actions of platelet-derived growth factor. J. Periodontol. 2007, 78, 1136–1145. [Google Scholar] [CrossRef]
- George, J.; Headen, K.V.; Ogunleye, A.O.; Perry, G.A.; Wilwerding, T.M.; Parrish, L.C.; McVaney, T.P.; Mattson, J.S.; Cerutis, D.R. Lysophosphatidic Acid signals through specific lysophosphatidic Acid receptor subtypes to control key regenerative responses of human gingival and periodontal ligament fibroblasts. J. Periodontol. 2009, 80, 1338–1347. [Google Scholar] [CrossRef]
- Cerutis, D.R.; Headen, K.V.; Perry, G.; Parrish, L.C.; McVaney, T.P.; Jordan, C.S. Lysophosphatidic acid (LPA) receptor subtypes on human gingival and periodontal ligament fibroblasts are regulated by PDDGF. FASEB J. 2010, 24, 769.11. [Google Scholar] [CrossRef]
- Lin, C.I.; Chen, C.N.; Lin, P.W.; Chang, K.J.; Hsieh, F.J.; Lee, H. Lysophosphatidic acid regulates inflammation-related genes in human endothelial cells through LPA1 and LPA3. Biochem. Biophys. Res. Commun. 2007, 363, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Kihara, Y.; Mizuno, H.; Chun, J. Lysophospholipid receptors in drug discovery. Exp. Cell Res. 2015, 333, 171–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yatomi, Y.; Ruan, F.; Hakomori, S.; Igarashi, Y. Sphingosine-1-phosphate: A platelet-activating sphingolipid released from agonist-stimulated human platelets. Blood 1995, 86, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.I.; Chen, C.N.; Lin, P.W.; Lee, H. Sphingosine 1-phosphate regulates inflammation-related genes in human endothelial cells through S1P1 and S1P3. Biochem. Biophys. Res. Commun. 2007, 355, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Hashimura, S.; Kido, J.; Matsuda, R.; Yokota, M.; Matsui, H.; Inoue-Fujiwara, M.; Inagaki, Y.; Hidaka, M.; Tanaka, T.; Tsutsumi, T.; et al. A low level of lysophosphatidic acid in human gingival crevicular fluid from patients with periodontitis due to high soluble lysophospholipase activity: Its potential protective role on alveolar bone loss by periodontitis. BBA-Mol. Cell. Biol. Lipids 2020, 1865, 158698. [Google Scholar] [CrossRef]
- Bathena, S.P.; Huang, J.; Nunn, M.E.; Miyamoto, T.; Parrish, L.C.; Lang, M.S.; McVaney, T.P.; Toews, M.L.; Cerutis, D.R.; Alnouti, Y. Quantitative determination of lysophosphatidic acids (LPAs) in human saliva and gingival crevicular fluid (GCF) by LC-MS/MS. J. Pharm. Biomed. Anal. 2011, 56, 402–407. [Google Scholar] [CrossRef] [Green Version]
- Cerutis, D.R.; Weston, M.D.; Ogunleye, A.O.; McVaney, T.P.; Miyamoto, T. Lysophosphatidic acid (LPA) 18:1 transcriptional regulation of primary human gingival fibroblasts. Genom Data 2014, 2, 375–377. [Google Scholar] [CrossRef] [Green Version]
- Cerutis, D.R.; Weston, M.D.; Alnouti, Y.; Bathena, S.P.; Nunn, M.E.; Ogunleye, A.O.; McVaney, T.P.; Headen, K.V.; Miyamoto, T. A Major Human Oral Lysophosphatidic Acid Species, LPA 18:1, Regulates Novel Genes in Human Gingival Fibroblasts. J. Periodontol. 2015, 86, 713–725. [Google Scholar] [CrossRef]
- Gruber, R.; Kandler, B.; Jindra, C.; Watzak, G.; Watzek, G. Dental pulp fibroblasts contain target cells for lysophosphatidic Acid. J. Dent. Res. 2004, 83, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Thorlakson, H.H.; Engen, S.A.; Schreurs, O.; Schenck, K.; Blix, I.J.S. Lysophosphatidic acid induces expression of genes in human oral keratinocytes involved in wound healing. Arch. Oral Biol. 2017, 80, 153–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Seo, E.J.; Tigyi, G.J.; Lee, B.J.; Jang, I.H. The role of lysophosphatidic acid receptor 1 in inflammatory response induced by lip-opolysaccharide from Porphyromonas gingivalis in human periodontal ligament stem cells. International. J. Oral Biol. 2020, 45, 42–50. [Google Scholar] [CrossRef]
- Vatėnas, I.; Linkevičius, T. The use of the connective tissue graft from the palate for vertical soft tissue augmentation during submerged dental implant placement: A case series. Clin. Exp. Dent. Res. 2022, 8, 1103–1108. [Google Scholar] [CrossRef]
- Liu, Y.; Karode, K.; Bodine, P.V.N.; Yaworsky, P.J.; Robinson, J.A.; Billiard, J. LPA induces osteoblast differentiation through interplay of two receptors: LPA1 and LPA4. J. Cell. Biochem. 2010, 109, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Gennero, I.; Laurencin-Dalicieux, S.; Conte-Auriol, F.; Briand-Mésange, F.; Laurencin, D.; Rue, J.; Beton, N.; Malet, N.; Mus, M.; Tokumura, A.; et al. Absence of the lysophosphatidic acid receptor LPA1 results in abnormal bone development and decreased bone mass. Bone 2011, 49, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Song, Z.; Chen, R.; Tan, S.; Huang, C.; Liu, Y.; Cheng, B.; Fu, Q. Lysophosphatidic acid enhanced the osteogenic and angiogenic capability of osteoblasts via LPA1/3 receptor. Connect. Tissue Res. 2019, 60, 85–94. [Google Scholar] [CrossRef]
- Tanaka, M.; Okudaira, S.; Kishi, Y.; Ohkawa, R.; Iseki, S.; Ota, M.; Noji, S.; Yatomi, Y.; Aoki, J.; Arai, H. Autotaxin stabilizes blood vessels and is required for embryonic vasculature by producing lysophosphatidic acid. J. Biol. Chem. 2006, 281, 25822–25830. [Google Scholar] [CrossRef] [Green Version]
- Van Meeteren, L.A.; Ruurs, P.; Stortelers, C.; Bouwman, P.; van Rooijen, M.A.; Pradère, J.P.; Pettit, T.R.; Wakelam, M.J.; Saulnier-Blache, J.S.; Mummery, C.L.; et al. Autotaxin, a secreted lysophospholipase D, is essential for blood vessel formation during development. Mol. Cell Biol. 2006, 26, 5015–5022. [Google Scholar] [CrossRef] [Green Version]
- Economides, A.N.; Frendewey, D.; Yang, P.; Dominguez, M.G.; Dore, A.T.; Lobov, I.B.; Persaud, T.; Rojas, J.; McClain, J.; Lengyel, P.; et al. Conditionals by inversion provide a universal method for the generation of conditional alleles. Proc. Natl. Acad. Sci. USA 2013, 110, E3179–E3188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Hama, K.; Contos, J.J.; Anliker, B.; Inoue, A.; Skinner, M.K.; Suzuki, H.; Amano, T.; Kennedy, G.; Arai, H.; et al. LPA3-mediated lysophosphatidic acid signalling in embryo implantation and spacing. Nature 2005, 435, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Hama, K.; Aoki, J.; Inoue, A.; Endo, T.; Amano, T.; Motoki, R.; Kanai, M.; Ye, X.; Chun, J.; Matsuki, N.; et al. Embryo spacing and implantation timing are differentially regulated by LPA3-mediated lysophosphatidic acid signaling in mice. Biol. Reprod. 2007, 77, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Vittek, J.; Hernandez, M.R.; Wenk, E.J.; Rappaport, S.C.; Southren, A.L. Specific estrogen receptors in human gingiva. J. Clin. Endocrinol. Metab. 1982, 54, 608–612. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, A.; Mawhinney, M. Endocrinology of sex steroid hormones and cell dynamics in the periodontium. Periodontol 2000 2013, 61, 69–88. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.S.; Lee, S.K.; Parke, K.; Chunga, W. Secretion of IL-6 and IL-8 from lysophosphatidic acid-stimulated oral squamous cell carcinoma promotes osteoclastogenesis and bone resorption. Oral Oncol. 2012, 48, 40–48. [Google Scholar] [CrossRef]
- Nakasaki, T.; Tanaka, T.; Okudaira, S.; Hirosawa, M.; Umemoto, E.; Otani, K.; Jin, S.; Bai, Z.; Hayasaka, H.; Fukui, Y.; et al. Involvement of the lysophosphatidic acid-generating enzyme autotaxin in lymphocyte-endothelial cell interactions. Am. J. Pathol. 2008, 173, 1566–1576. [Google Scholar] [CrossRef] [Green Version]
- Payne, S.G.; Milstien, S.; Spiegel, S. Sphingosine-1-phosphate: Dual messenger functions. FEBS Lett. 2002, 531, 54–57. [Google Scholar] [CrossRef] [Green Version]
- Xia, P.; Wadham, C. Sphingosine 1-phosphate, a key mediator of the cytokine network: Juxtacrine signaling. Cytokine Growth Factor Rev. 2011, 22, 45–53. [Google Scholar] [CrossRef]
- Hatoum, D.; Haddadi, N.; Lin, Y.; Nassif, N.T.; McGowan, E.M. Mammalian sphingosine kinase (SphK) isoenzymes and isoform expression: Challenges for SphK as an oncotarget. Oncotarget 2017, 8, 36898–36929. [Google Scholar] [CrossRef] [Green Version]
- Blaho, V.A.; Hla, T. An uPDDate on the biology of sphingosine 1-phosphate receptors. J. Lipid Res. 2014, 55, 1596–1608. [Google Scholar] [CrossRef] [Green Version]
- Olivera, A.; Allende, M.L.; Proia, R.L. Shaping the landscape: Metabolic regulation of S1P gradients. Biochim. Biophys. Acta. 2013, 1831, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Moritz, E.; Wegner, D.; Groß, S.; Bahls, M.; Dörr, M.; Felix, S.B.; Ittermann, T.; Oswald, S.; Nauck, M.; Friedrich, N.; et al. Reference intervals for serum sphingosine-1-phosphate in the population-based Study of Health in Pomerania. Clin. Chim. Acta 2017, 468, 25–31. [Google Scholar] [CrossRef]
- Moritz, E.; Wegner, D.; Groß, S.; Bahls, M.; Dörr, M.; Felix, S.B.; Ittermann, T.; Oswald, S.; Nauck, M.; Friedrich, N.; et al. Data on subgroup specific baseline characteristics and serum sphingosine-1-phosphate concentrations in the Study of Health in Pomerania. Data Brief 2017, 12, 46–50. [Google Scholar] [CrossRef]
- Moritz, E.; Jedlitschky, G.; Negnal, J.; Tzvetkov, M.V.; Daum, G.; Dörr, M.; Felix, S.B.; Völzke, H.; Nauck, M.; Schwedhelm, E.; et al. Increased Sphingosine-1-Phosphate Serum Concentrations in Subjects with Periodontitis: A Matter of Inflammation. J. Inflamm. Res. 2021, 14, 2883–2896. [Google Scholar] [CrossRef]
- Rhim, J.H.; Jang, I.S.; Choi, J.S.; Kwon, H.J.; Yeo, E.J.; Park, S.C. Time-dependent differential gene expression in lysophosphatidic acid-treated young and senescent human diploid fibroblasts. Mech. Ageing Dev. 2009, 130, 648–651. [Google Scholar] [CrossRef]
- Luheshi, N.M.; Giles, J.A.; Lopez-Castejon, G.; Brough, D. Sphingosine regulates the NLRP3-inflammasome and IL-1 beta release from macrophages. Eur. J. Immunol. 2012, 42, 716–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yang, L.; Tian, L.; Ji, X.; Yang, L.; Li, L. Sphingosine 1-phosphate (S1P)/S1P receptor 2/3 axis promotes inflammatory M1 polarization of bone marrow-derived monocyte/macrophage via G(α)i/o/PI3K/JNK pathway. Cell Physiol. Biochem. 2018, 49, 1677–1693. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.J.; Shin, K.O.; Kim, H.; Ahn, S.H.; Lee, S.H.; Seo, C.H.; Byun, S.E.; Chang, J.S.; Koh, J.M.; Lee, Y.M. The effect of sphingosine-1-phosphate on bone metabolism in humans depends on its plasma/bone marrow gradient. J. Endocrinol. Invest. 2016, 39, 297–303. [Google Scholar] [CrossRef]
- Bourgoin, S.G.; Zhao, C. Autotaxin and lysophospholipids in rheumatoid arthritis. Curr. Opin. Investig. Drugs 2010, 11, 515–526. [Google Scholar] [PubMed]
- Burg, N.; Salmon, J.E.; Hla, T. Sphingosine 1-phosphate receptor-targeted therapeutics in rheumatic diseases. Nat. Rev. Rheumatol. 2022, 18, 335–351. [Google Scholar] [CrossRef] [PubMed]
- Eskan, M.A.; Rose, B.G.; Benakanakere, M.R.; Lee, M.J.; Kinane, D.F. Sphingosine 1-phosphate 1 and TLR4 mediate IFN-beta expression in human gingival epithelial cells. J. Immunol. 2008, 180, 1818–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Sun, C.; Argraves, K.M. Periodontal inflammation and alveolar bone loss induced by Aggregatibacter actinomycetemcomitans is attenuated in sphingosine kinase 1-deficient mice. J. Periodontal. Res. 2016, 51, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Meerschaert, K.; De Corte, V.; De Ville, Y.; Vandekerckhove, J.; Gettemans, J. Gelsolin and functionally similar actin-binding proteins are regulated by lysophosphatidic acid. EMBO J. 1998, 17, 5923–5932. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Bi, X.; Luo, Z.; Wang, H.; Ismtula, D.; Guo, C. Gelsolin: A comprehensive pan-cancer analysis of potential prognosis, diagnostic, and immune biomarkers. Front. Genet. 2023, 14, 1093163. [Google Scholar] [CrossRef]
- Thorp, H.H.; Yaffe, M.B. Seeing is great, understanding is better. Sci. Signal. 2023, 16, eadg2868. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y. Targeting Lysophosphatidic Acid in Cancer: The Issues in Moving from Bench to Bedside. Cancers 2019, 11, 1523. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.W.; Herr, D.R.; Noguchi, K.; Yung, Y.C.; Lee, C.W.; Mutoh, T.; Lin, M.E.; Teo, S.T.; Park, K.E.; Mosley, A.N.; et al. LPA receptors: Subtypes and biological actions. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 157–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, Y.; Sardar, V.; Tokumura, A.; Baker, D.; Murakami-Murofushi, K.; Parrill, A.; Tigyi, G. Identification of residues responsible for ligand recognition and regioisomeric selectivity of lysophosphatidic acid receptors expressed in mammalian cells. J. Biol. Chem. 2005, 280, 35038–35050. [Google Scholar] [CrossRef] [Green Version]
- Parrill, A.L. Lysophospholipid interactions with protein targets. Biochim. Biophys. Acta 2008, 1781, 540–546. [Google Scholar] [CrossRef] [Green Version]
- Im, D.S.; Heise, C.E.; Harding, M.A.; George, S.R.; O’Dowd, B.F.; Theodorescu, D.; Lynch, K.R. Molecular cloning and characterization of a lysophosphatidic acid receptor, Edg-7, expressed in prostate. Mol. Pharmacol. 2000, 57, 753–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, M.; Nagai, K.; Kihara, Y.; Kussrow, A.; Kammer, M.N.; Frantz, A.; Bornhop, D.J.; Chun, J. Unlabeled lysophosphatidic acid receptor binding in free solution as determined by a compensated interferometric reader. J. Lipid Res. 2020, 61, 1244–1251. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, H.; Kihara, Y.; Kussrow, A.; Chen, A.; Ray, M.; Rivera, R.; Bornhop, D.J.; Chun, J. Lysophospholipid G protein-coupled receptor binding parameters as determined by backscattering interferometry. J. Lipid Res. 2019, 60, 212–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagida, K.; Masago, K.; Nakanishi, H.; Kihara, Y.; Hamano, F.; Tajima, Y.; Taguchi, R.; Shimizu, T.; Ishii, S. Identification and characteri-zation of a novel lysophosphatidic acid receptor, p2y5/LPA6. J. Biol. Chem. 2009, 284, 17731–17741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannun, Y.; Obeid, L. Principles of bioactive lipid signalling: Lessons from sphingolipids. Nat. Rev. Mol. Cell Biol. 2008, 9, 139–150. [Google Scholar] [CrossRef]
- Leblanc, R.; Peyruchaud, O. New insights into the autotaxin/LPA axis in cancer development and metastasis. Exp. Cell Res. 2015, 333, 183–189. [Google Scholar] [CrossRef]
- Sugiura, T.; Nakane, S.; Kishimoto, S.; Waku, K.; Yoshioka, Y.; Tokumura, A. Lysophosphatidic acid, a growth factor-like lipid, in the saliva. J. Lipid Res. 2002, 43, 2049–2055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chae, C.S.; Sandoval, T.A.; Hwang, S.M.; Park, E.S.; Giovanelli, P.; Awasthi, D.; Salvagno, C.; Emmanuelli, A.; Tan, C.; Chaudhary, V.; et al. Tumor-Derived Lysophosphatidic Acid Blunts Protective Type I Interferon Responses in Ovarian Cancer. Cancer Discov. 2022, 12, 1904–1921. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; DuBois, R.N. The Role of Prostaglandin E(2) in Tumor-Associated Immunosuppression. Trends Mol. Med. 2016, 22, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Bell, C.R.; Pelly, V.S.; Moeini, A.; Chiang, S.C.; Flanagan, E.; Bromley, C.P.; Clark, C.; Earnshaw, C.H.; Koufaki, M.A.; Bonavita, E.; et al. Chemotherapy-induced COX-2 upregulation by cancer cells defines their inflammatory properties and limits the efficacy of chemoimmunotherapy combinations. Nat. Commun. 2022, 3, 2063. [Google Scholar] [CrossRef]
- Reinartz, S.; Lieber, S.; Pesek, J.; Brandt, D.T.; Asafova, A.; Finkernagel, F.; Watzer, B.; Nockher, W.A.; Nist, A.; Stiewe, T.; et al. Cell type-selective pathways and clinical associations of lysophosphatidic acid biosynthesis and signaling in the ovarian cancer microenvironment. Mol. Oncol. 2019, 13, 185–201. [Google Scholar] [CrossRef] [Green Version]
- Dacheux, M.A.; Lee, S.C.; Shin, Y.; Norman, D.D.; Lin, K.H.; E, S.; Yue, J.; Benyó, Z.; Tigyi, G.J. Prometastatic Effect of ATX Derived from Alveolar Type II Pneumocytes and B16-F10 Melanoma Cells. Cancers 2022, 14, 1586. [Google Scholar] [CrossRef]
- Turner, J.A.; Fredrickson, M.A.; D’Antonio, M.; Katsnelson, E.; MacBeth, M.; Van Gulick, R.; Chimed, T.S.; McCarter, M.; D’Alessandro, A.; Robinson, W.A.; et al. Lysophosphatidic acid modulates CD8 T cell immunosurveillance and metabolism to impair anti-tumor immunity. Nat. Commun. 2023, 14, 214. [Google Scholar] [CrossRef] [PubMed]
- Abdul Rahman, M.; Mohamad Haron, D.E.; Hollows, R.J.; Abdul Ghani, Z.D.F.; Ali Mohd, M.; Chai, W.L.; Ng, C.C.; Lye, M.S.; Karsani, S.A.; Yap, L.F.; et al. Profiling lysophosphatidic acid levels in plasma from head and neck cancer patients. PeerJ. 2020, 8, e9304. [Google Scholar] [CrossRef] [PubMed]
- Cerutis, D.R.; Kumar, D.; Nichols, M.G.; Roemer, G.; Fluent, M.; Beller, L.; Miyamoto, T.; Alnouti, Y. Lysophosphatidic Acid (LPA) Salivary Species Detection and In Situ LPAR Localization in the Intact Mouse Salivary Gland. J. Pharmacol. Exp. Ther. 2023, 385, 79. [Google Scholar] [CrossRef]
- Goetzl, E.J.; Lee, H.; Azuma, T.; Stossel, T.P.; Turck, C.W.; Karliner, J.S. Gelsolin binding and cellular presentation of lysophosphatidic acid. J Biol Chem. 2000, 275, 14573–14578. [Google Scholar] [CrossRef] [Green Version]
- Watt, M.J.; Hoy, A.J. Lipid metabolism in skeletal muscle: Generation of adaptive and maladaptive intracellular signals for cellular function. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1315–E1328. [Google Scholar] [CrossRef] [Green Version]
- Piktel, E.; Levental, I.; Durnaś, B.; Janmey, P.A.; Bucki, R. Plasma Gelsolin: Indicator of Inflammation and Its Potential as a Diagnostic Tool and Therapeutic Target. Int. J. Mol. Sci. 2018, 19, 2516. [Google Scholar] [CrossRef] [Green Version]
- Feldt, J.; Schicht, M.; Garreis, F.; Welss, J.; Schneider, U.W.; Paulsen, F. Structure, regulation and related diseases of the actin-binding protein gelsolin. Expert Rev Mol Med. 2019, 20, e7. [Google Scholar] [CrossRef]
- Fleming, J.K.; Glass, T.R.; Lackie, S.J.; Wojciak, J.M. A novel approach for measuring sphingosine-1-phosphate and lysophosphatidic acid binding to carrier proteins using monoclonal antibodies and the Kinetic Exclusion Assay. J. Lipid Res. 2016, 57, 1737–1747. [Google Scholar] [CrossRef] [Green Version]
- Galvani, S.; Sanson, M.; Blaho, V.A.; Swendeman, S.L.; Obinata, H.; Conger, H.; Dahlbäck, B.; Kono, M.; Proia, R.L.; Smith, J.D.; et al. HDL-bound sphingosine 1-phosphate acts as a biased agonist for the endothelial cell receptor S1P1 to limit vascular inflammation. Sci Signal. 2015, 8, ra79, Erratum in Sci Signal. 2015, 8, er8. [Google Scholar] [CrossRef] [Green Version]
- Blaho, V.A.; Galvani, S.; Engelbrecht, E.; Lin, C.; Swendeman, S.L.; Kono, M.; Proia, R.L.; Steinman, L.; Han, M.H.; Hla, T. HDL-bound sphingosine-1-phosphate restrains lymphopoesis and neuroinflammation. Nature 2015, 523, 342–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkerson, B.A.; Grass, G.D.; Wing, S.B.; Argraves, W.S.; Argraves, K.M. Sphingosine-1-phosphate (S1P) carrier-dependent regulation of endothelial barrier: High density lipoprotein (HDL)-S1P prolongs endothelial barrier enhancement as compared with albumin-S1P via effects on levels, trafficking, and signaling of S1P1. J. Biol. Chem. 2012, 287, 44645–44653. [Google Scholar] [CrossRef] [Green Version]
- Dahm, F.; Nocito, A.; Bielawska, A.; Lang, K.S.; Georgiev, P.; Asmis, L.M.; Bielawski, J.; Madon, J.; Hannun, Y.A.; Clavien, P.A. Distribution and dynamic changes of sphingolipids in blood in response to platelet activation. J. Thromb. Haemost. 2006, 4, 2704–2709. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, A.A.; Horrocks, L.A.; Farooqui, T. Interactions between neural membrane glycerophospholipid and sphingolipid mediators: A recipe for neural cell survival or suicide. J. Neurosci. Res. 2007, 85, 1834–1850. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez de San Roman, E.; Llorente-Ovejero, A.; Martinez-Gardeazabal, J.; Moreno-Rodriguez, M.; Gimenez-Llort, L.; Manuel, I.; Rodriguez-Puertas, R. Modulation of neurolipid signaling and specific lipid species in the triple transgenic mouse model of Alzheimer’s disease. Int. J. Mol. Sci. 2021, 22, 12256. [Google Scholar] [CrossRef]
- Schwartz, S.A.; Reyzer, M.L.; Caprioli, R.M. Direct tissue analysis using matrix-assisted laser desorption/ionization mass spectrometry: Practical aspects of sample preparation. J. Mass Spectrom. 2003, 38, 699–708. [Google Scholar] [CrossRef]
- Wang, J.; Wang, C.; Han, X. Enhanced coverage of lipid analysis and imaging by matrix-assisted laser desorption/ionization mass spectrometry via a strategy with an optimized mixture of matrices. Anal. Chim. Acta 2018, 1000, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Lipidomics Standards Initiative Consortium. Lipidomics needs more standardization. Nat. Metab. 2019, 1, 745–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, H.; Kawasaki, A.; Takahashi, M.; Yamada, A.; Koike, T. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry of phosphorylated compounds using a novel phosphate capture molecule. Rapid Commun. Mass Spectrom. 2003, 17, 2075–2081. [Google Scholar] [CrossRef]
- Morishige, J.; Urikura, M.; Takagi, H.; Hirano, K.; Koike, T.; Tanaka, T.; Satouchi, K. A clean-up technology for the simultaneous determination of lysophosphatidic acid and sphingosine-1-phosphate by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry using a phosphate-capture molecule, Phos-tag. Rapid Commun. Mass Spectrom. 2010, 24, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Iwama, T.; Kano, K.; Saigusa, D.; Ekroos, K.; van Echten-Deckert, G.; Vogt, J.; Aoki, J. Development of an on-tissue derivatization method for MALDI mass spectrometry imaging of bioactive lipids containing phosphate monoester using Phos-tag. Anal. Chem. 2021, 93, 3867–3875. [Google Scholar] [CrossRef] [PubMed]
- Alekseenko, I.; Kondratyeva, L.; Chernov, I.; Sverdlov, E. From the catastrophic objective irreproducibility of cancer research and unavoidable failures of molecular targeted therapies to the sparkling hope of supramolecular targeted strategies. Int. J. Mol. Sci. 2023, 24, 2796. [Google Scholar] [CrossRef] [PubMed]
- Casadó, V.; Cortés, A.; Mallol, J.; Pérez-Capote, K.; Ferré, S.; Lluis, C.; Franco, R.; Canela, E.I. GPCR homomers and heteromers: A better choice as targets for drug development than GPCR monomers? Pharmacol. Ther. 2009, 124, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, V.V.; Gurevich, E.V. How and why do GPCRs dimerize? Trends Pharmacol. Sci. 2008, 29, 234–240. [Google Scholar] [CrossRef] [Green Version]
- Hazum, E.; Chang, K.J.; Cuatrecasas, P. Opiate (Enkephalin) receptors of neuroblastoma cells: Occurrence in clusters on the cell surface. Science 1979, 206, 1077–1079. [Google Scholar] [CrossRef]
- Vischer, H.F.; Castro, M.; Pin, J.P. G Protein-Coupled Receptor Multimers: A Question Still Open Despite the Use of Novel Ap-proaches. Mol. Pharmacol. 2015, 88, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Albizu, L.; Cottet, M.; Kralikova, M.; Stoev, S.; Seyer, R.; Brabet, I.; Roux, T.; Bazin, H.; Bourrier, E.; Lamarque, L.; et al. Time-resolved FRET between GPCR ligands reveals oli-gomers in native tissues. Nat. Chem. Biol. 2010, 6, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Rivero-Müller, A.; Chou, Y.Y.; Ji, I.; Lajic, S.; Hanyaloglu, A.C.; Jonas, K.; Rahman, N.; Ji, T.H.; Huhtaniemi, I. Rescue of defective G pro-tein-coupled receptor function in vivo by intermolecular cooperation. Proc. Natl. Acad. Sci. USA 2010, 107, 2319–2324. [Google Scholar] [CrossRef] [Green Version]
- Kasai, R.S.; Ito, S.V.; Awane, R.M.; Fujiwara, T.K.; Kusumi, A. The Class-A GPCR Dopamine D2 Receptor Forms Transient Dimers Stabi-lized by Agonists: Detection by Single-Molecule Tracking. Cell Biochem. Biophys. 2018, 76, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Pei, L.; Fletcher, P.J.; Kapur, S.; Seeman, P.; Liu, F. Schizophrenia, amphetamine-induced sensitized state and acute ampheta-mine exposure all show a common alteration: Increased dopamine D2 receptor dimerization. Mol. Brain 2010, 3, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Tang, H.; Xu, C.; Zhou, S.; Zhu, X.; Li, Y.; Prézeau, L.; Xu, T.; Pin, J.P.; Rondard, P.; et al. Biased signaling due to oligomerization of the G protein-coupled platelet-activating factor receptor. Nat. Commun. 2022, 13, 6365. [Google Scholar] [CrossRef]
- Zhu, T.; Gobeil, F.; Vazquez-Tello, A.; Leduc, M.; Rihakova, L.; Bossolasco, M.; Bkaily, G.; Peri, K.; Varma, D.R.; Orvoine, R.; et al. In-tracrine signaling through lipid mediators and their cognate nuclear G-protein-coupled receptors: A paradigm based on PGE2, PAF, and LPA1 receptors. Can. J. Physiol. Pharmacol. 2006, 84, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Godbole, A.; Lyga, S.; Lohse, M.J.; Calebiro, D. Internalized TSH receptors en route to the TGN induce local Gs-protein signaling and gene transcription. Nat. Commun. 2017, 8, 443, Erratum in Nat. Commun. 2018, 9, 5459. [Google Scholar] [CrossRef] [Green Version]
- Cerutis, D.R.; Nichols, M.G.; Khan, S.A.; Miyamoto, T. Localization of the Platelet-Activating Factor Receptor on the Intact Human Periodontal Ligament. FASEB J. 2017, 31, lb24. [Google Scholar]
- Cerutis, D.R.; Nichols, M.; Hironaka, M.; Miyamoto, T.; Khan, S.; Ogunleye, A.; McVaney, T. Complementing Confocal Detection of Antibody-labeled Lysophosphatidic Acid Receptors in Human Gingivae with Label-free Second Harmonic Generation Confocal Microscopy Detection of Collagen. FASEB J. 2015, 29, LB26. [Google Scholar] [CrossRef]
- Milstein, J.N.; Nino, D.F.; Zhou, X.; Gradinaru, C.C. Single-molecule counting applied to the study of GPCR oligomerization. Biophys. J. 2022, 121, 3175–3187. [Google Scholar] [CrossRef]
- Shonberg, J.; Scammells, P.J.; Capuano, B. Design strategies for bivalent ligands targeting GPCRs. ChemMedChem 2011, 6, 963–974. [Google Scholar] [CrossRef]
- Lee, S.C.; Fujiwara, Y.; Tigyi, G.J. Uncovering unique roles of LPA receptors in the tumor microenvironment. Recept. Clin. Investig. 2015, 2, e440. [Google Scholar] [CrossRef]
- Rahaman, M.; Costello, R.W.; Belmonte, K.E.; Gendy, S.S.; Walsh, M.T. Neutrophil sphingosine 1-phosphate and lysophosphatidic acid receptors in pneumonia. Am. J. Respir. Cell Mol. Biol. 2006, 34, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panigrahy, D.; Gilligan, M.M.; Serhan, C.N.; Kashfi, K. Resolution of inflammation: An organizing principle in biology and medicine. Pharmacol. Ther. 2021, 227, 107879. [Google Scholar] [CrossRef] [PubMed]
- Zaslavsky, A.; Singh, L.S.; Tan, H.; Ding, H.; Liang, Z.; Xu, Y. Homo- and hetero-dimerization of LPA/S1P receptors, OGR1 and GPR4. Biochim. Biophys. Acta 2006, 1761, 1200–1212. [Google Scholar] [CrossRef] [PubMed]
- Hisano, Y.; Kono, M.; Cartier, A.; Engelbrecht, E.; Kano, K.; Kawakami, K.; Xiong, Y.; Piao, W.; Galvani, S.; Yanagida, K.; et al. Lysolipid receptor cross-talk regulates lymphatic endothelial junctions in lymph nodes. J. Exp. Med. 2019, 216, 1582–1598. [Google Scholar] [CrossRef] [PubMed]
- GPCR Interaction Network. Available online: http://www.gpcr-hetnet.com/ (accessed on 3 May 2023).
- Stoeber, M.; Jullié, D.; Lobingier, B.T.; Laeremans, T.; Steyaert, J.; Schiller, P.W.; Manglik, A.; von Zastrow, M. A Genetically Encoded Biosensor Reveals Location Bias of Opioid Drug Action. Neuron 2018, 98, 963–976.e5. [Google Scholar] [CrossRef] [Green Version]
- Shariati, L.; Esmaeili, Y.; Haghjooy Javanmard, S.; Bidram, E.; Amini, A. Organoid technology: Current standing and future perspectives. Stem Cells 2021, 39, 1625–1649. [Google Scholar] [CrossRef]
- Zhao, X.; Xu, Z.; Xiao, L.; Shi, T.; Xiao, H.; Wang, Y.; Li, Y.; Xue, F.; Zeng, W. Review on the vascularization of organoids and organoids-on-a-chip. Front. Bioeng. Biotechnol. 2021, 9, 637048. [Google Scholar] [CrossRef]
- Bhushan, A.; Senutovich, N.; Bale, S.S.; McCarty, W.J.; Hegde, M.; Jindal, R.; Golberg, I.; Usta, O.B.; Yarmush, M.L.; Vernetti, L.; et al. Towards a three-dimensional microfluidic liver platform for predicting drug efficacy and toxicity in humans. Stem Cell Res. Ther. 2013, 4 (Suppl 1), S16. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, B.; Zinocker, S.; Holm, S.; Lewis, J.; Kavouras, P. Organoids in the clinic: A systematic review of outcomes. Cells Tissues Organs 2022. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerutis, D.R.; Weston, M.D.; Miyamoto, T. Entering, Linked with the Sphinx: Lysophosphatidic Acids Everywhere, All at Once, in the Oral System and Cancer. Int. J. Mol. Sci. 2023, 24, 10278. https://doi.org/10.3390/ijms241210278
Cerutis DR, Weston MD, Miyamoto T. Entering, Linked with the Sphinx: Lysophosphatidic Acids Everywhere, All at Once, in the Oral System and Cancer. International Journal of Molecular Sciences. 2023; 24(12):10278. https://doi.org/10.3390/ijms241210278
Chicago/Turabian StyleCerutis, D. Roselyn, Michael D. Weston, and Takanari Miyamoto. 2023. "Entering, Linked with the Sphinx: Lysophosphatidic Acids Everywhere, All at Once, in the Oral System and Cancer" International Journal of Molecular Sciences 24, no. 12: 10278. https://doi.org/10.3390/ijms241210278