
Plasma Lipidomics Analysis Reveals the Potential Role of Lysophosphatidylcholines in Abdominal Aortic Aneurysm Progression and Formation
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Baseline Characteristics of Enrolled Subjects
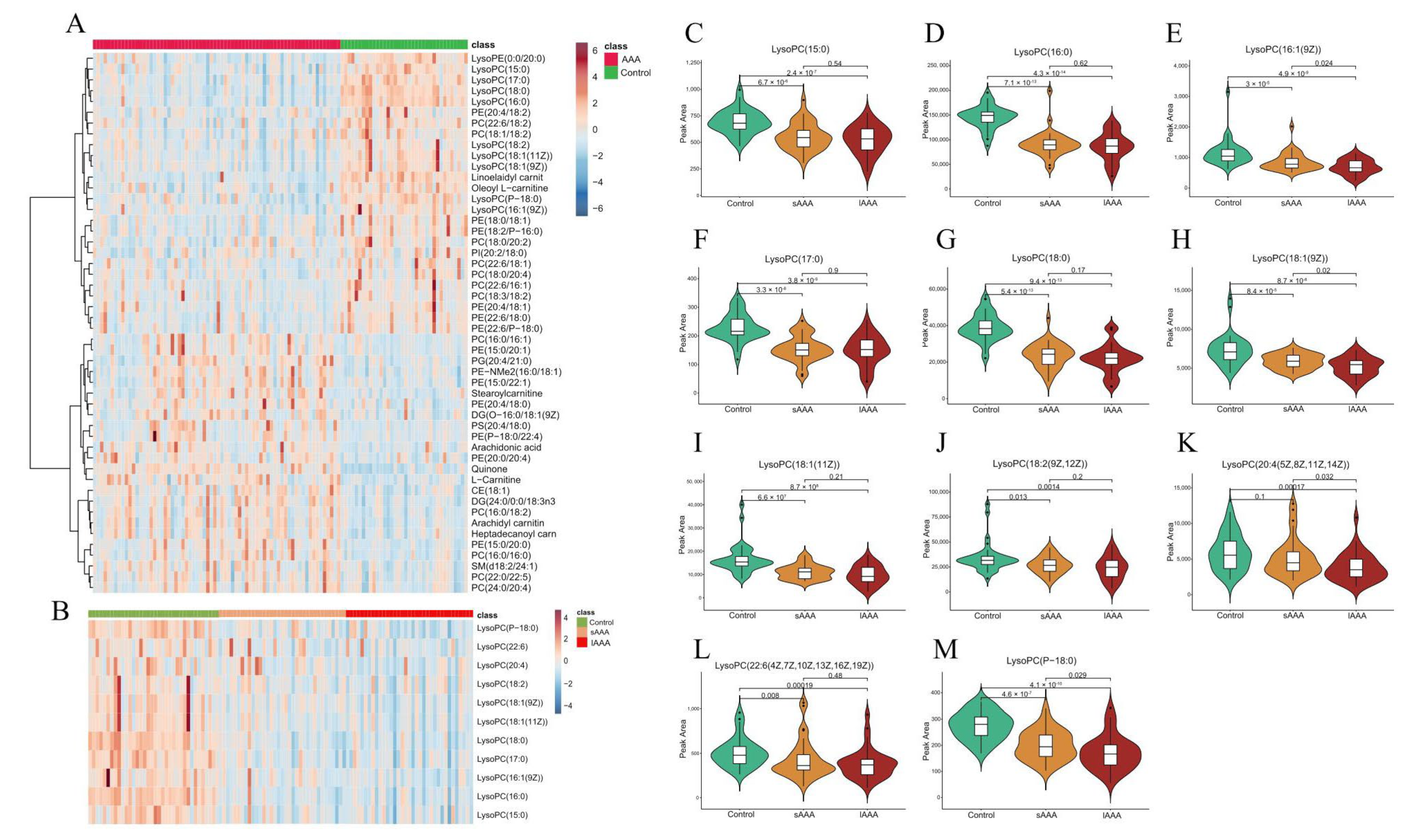
2.2. Changes in Plasma Lipidomics between Control and AAA Subjects
2.3. HDL-c and hsCRP Were Distinctly Associated with Altered Profiles of LysoPCs in AAA
2.4. AAA Formation in ApoE −/− Mice and LysoPCs Decrease with AAA Progression
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Animal Model and Sample Collection
4.3. Plasma Collection and Lipidomics Analysis
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Avishay, D.M.; Reimon, J.D. Abdominal Aortic Repair. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2022. [Google Scholar]
- Golledge, J.; Tsao, P.S.; Dalman, R.L.; Norman, P.E. Circulating markers of abdominal aortic aneurysm presence and progression. Circulation 2008, 118, 2382–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakalihasan, N.; Limet, R.; Defawe, O.D. Abdominal aortic aneurysm. Lancet 2005, 365, 1577–1589. [Google Scholar] [CrossRef] [PubMed]
- Peshkova, I.; Schaefer, G.; Koltsova, E. Atherosclerosis and Aortic Aneurysm: Is inflammation a common denominator? FEBS J. 2015, 283, 1636–1652. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.T.; Tromp, G.; Kuivaniemi, H.; Gretarsdottir, S.; Baas, A.F.; Giusti, B.; Strauss, E.; Van’t Hof, F.N.; Webb, T.R.; Erdman, R.; et al. Meta-Analysis of Genome-Wide Association Studies for Abdominal Aortic Aneurysm Identifies Four New Disease-Specific Risk Loci. Circ. Res. 2017, 120, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.C.; Holmes, M.V.; Burgess, S.; Asselbergs, F.W.; Jones, G.T.; Baas, A.F.; van’t Hof, F.N.; de Bakker, P.I.W.; Blankensteijn, J.D.; Powell, J.T.; et al. Genetic Association of Lipids and Lipid Drug Targets with Abdominal Aortic Aneurysm: A Meta-analysis. JAMA Cardiol. 2018, 3, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Forsdahl, S.H.; Singh, K.; Solberg, S.; Jacobsen, B.K. Risk Factors for Abdominal Aortic Aneurysms. Circulation 2009, 119, 2202–2208. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Gross, R. Global analyses of cellular lipidomes directly from crude extracts of biological samples by ESI mass spectrometry: A bridge to lipidomics. J. Lipid Res. 2003, 44, 1071–1079. [Google Scholar] [CrossRef] [Green Version]
- Ciborowski, M.; Teul, J.; Martin-Ventura, J.L.; Egido, J.; Barbas, C. Metabolomics with LC-QTOF-MS permits the prediction of disease stage in aortic abdominal aneurysm based on plasma metabolic fingerprint. PLoS ONE 2012, 7, e31982. [Google Scholar] [CrossRef] [Green Version]
- Ciborowski, M.; Martin-Ventura, J.L.; Meilhac, O.; Michel, J.B.; Ruperez, F.J.; Tuñon, J.; Egido, J.; Barbas, C. Metabolites secreted by human atherothrombotic aneurysms revealed through a metabolomic approach. J. Proteome Res. 2011, 10, 1374–1382. [Google Scholar] [CrossRef]
- Guo, Y.; Wan, S.; Han, M.; Zhao, Y.; Li, C.; Cai, G.; Zhang, S.; Sun, Z.; Hu, X.; Cao, H.; et al. Plasma Metabolomics Analysis Identifies Abnormal Energy, Lipid, and Amino Acid Metabolism in Abdominal Aortic Aneurysms. Med. Sci. Monit. 2020, 26, e926766. [Google Scholar] [CrossRef]
- Iribarren, C.; Darbinian, J.; Go, A.; Fireman, B.; Lee, C.; Grey, D. Traditional and Novel Risk Factors for Clinically Diagnosed Abdominal Aortic Aneurysm: The Kaiser Multiphasic Health Checkup Cohort Study. Ann. Epidemiol. 2007, 17, 669–678. [Google Scholar] [CrossRef]
- Mannheim, D.; Herrmann, J.; Versari, D.; Gössl, M.; Meyer, F.B.; McConnell, J.P.; Lerman, L.O.; Lerman, A. Enhanced expression of Lp-PLA2 and lysophosphatidylcholine in symptomatic carotid atherosclerotic plaques. Stroke 2008, 39, 1448–1455. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Fang, P.; Li, Y.; Kuo, Y.M.; Andrews, A.J.; Nanayakkara, G.; Johnson, C.; Fu, H.; Shan, H.; Du, F.; et al. Mitochondrial Reactive Oxygen Species Mediate Lysophosphatidylcholine-Induced Endothelial Cell Activation. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1090–1100. [Google Scholar] [CrossRef] [Green Version]
- Zakiev, E.R.; Sukhorukov, V.N.; Melnichenko, A.A.; Sobenin, I.A.; Ivanova, E.A.; Orekhov, A.N. Lipid composition of circulating multiple-modified low density lipoprotein. Lipids Health Dis. 2016, 15, 134. [Google Scholar] [CrossRef] [Green Version]
- Orsó, E.; Matysik, S.; Grandl, M.; Liebisch, G.; Schmitz, G. Human native, enzymatically modified and oxidized low density lipoproteins show different lipidomic pattern. Biochim. Biophys. Acta 2015, 1851, 299–306. [Google Scholar] [CrossRef]
- Law, S.H.; Chan, M.L.; Marathe, G.K.; Parveen, F.; Chen, C.H.; Ke, L.Y. An Updated Review of Lysophosphatidylcholine Metabolism in Human Diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef] [Green Version]
- Barber, M.N.; Risis, S.; Yang, C.; Meikle, P.J.; Staples, M.; Febbraio, M.A.; Bruce, C.R. Plasma lysophosphatidylcholine levels are reduced in obesity and type 2 diabetes. PLoS ONE 2012, 7, e41456. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Luo, F.; Wu, P.; Huang, Y.; Das, A.; Chen, S.; Chen, J.; Hu, X.; Li, F.; Fang, Z.; et al. Metabolomics reveals metabolite changes of patients with pulmonary arterial hypertension in China. J. Cell. Mol. Med. 2020, 24, 2484–2496. [Google Scholar] [CrossRef]
- Semba, R.D.; Zhang, P.; Adelnia, F.; Sun, K.; Gonzalez-Freire, M.; Salem Jr, N.; Brennan, N.; Spencer, R.G.; Fishbein, K.; Khadeer, M.; et al. Low plasma lysophosphatidylcholines are associated with impaired mitochondrial oxidative capacity in adults in the Baltimore Longitudinal Study of Aging. Aging Cell 2019, 18, e12915. [Google Scholar] [CrossRef] [Green Version]
- Abel, S.; Kock, M.; Schalkwyk, D.; Swanevelder, S.; Gelderblom, W. Altered lipid profile, oxidative status and hepatitis B virus interactions in human hepatocellular carcinoma. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 391–399. [Google Scholar] [CrossRef]
- Krautbauer, S.; Eisinger, K.; Wiest, R.; Liebisch, G.; Buechler, C. Systemic saturated lysophosphatidylcholine is associated with hepatic function in patients with liver cirrhosis. Prostaglandins Other Lipid Mediat. 2016, 124, 27–33. [Google Scholar] [CrossRef]
- Lindholt, J.S.; Kristensen, K.L.; Burillo, E.; Martinez-Lopez, D.; Calvo, C.; Ros, E.; Martín-Ventura, J.L.; Sala-Vila, A. Arachidonic Acid, but Not Omega-3 Index, Relates to the Prevalence and Progression of Abdominal Aortic Aneurysm in a Population-Based Study of Danish Men. J. Am. Heart Assoc. 2018, 7, e007790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanna, V.S.; Hafez, E.A.A. Synopsis of arachidonic acid metabolism: A review. J. Adv. Res. 2018, 11, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Quehenberger, O.; Armando, A.M.; Brown, A.H.; Milne, S.B.; Myers, D.S.; Merrill, A.H.; Bandyopadhyay, S.; Jones, K.N.; Kelly, S.; Shaner, R.L.; et al. Lipidomics reveals a remarkable diversity of lipids in human plasma. J. Lipid Res. 2010, 51, 3299–3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.T.; Tran, K.; Pierce, G.N.; Chan, A.C.; Karmin, O.; Choy, P.C. Lysophosphatidylcholine Stimulates the Release of Arachidonic Acid in Human Endothelial Cells. J. Biol. Chem. 1998, 273, 6830–6836. [Google Scholar] [CrossRef] [Green Version]
- Aiyar, N.; Disa, J.; Ao, Z.; Ju, H.; Nerurkar, S.; Willette, R.N.; Macphee, C.H.; Johns, D.G.; Douglas, S.A. Lysophosphatidylcholine induces inflammatory activation of human coronary artery smooth muscle cells. Mol. Cell. Biochem. 2007, 295, 113–120. [Google Scholar] [CrossRef]
- Williams, B.; Mancia, G.; Spiering, W.; Agabiti Rosei, E.; Azizi, M.; Burnier, M.; Clement, D.L.; Coca, A.; de Simone, G.; Dominiczak, A.; et al. 2018 ESC/ESH Guidelines for the management of arterial hypertension: The Task Force for the management of arterial hypertension of the European Society of Cardiology (ESC) and the European Society of Hypertension (ESH). Eur. Heart J. 2018, 39, 3021–3104. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes, A. Diagnosis and classification of diabetes mellitus. Diabetes Care 2010, 33 (Suppl. 1), S62–S69. [Google Scholar] [CrossRef] [Green Version]
- Xie, T.; Yin, L.; Guo, D.; Zhang, Z.; Chen, Y.; Liu, B.; Wang, W.; Zheng, Y. The potential role of plasma fibroblast growth factor 21 as a diagnostic biomarker for abdominal aortic aneurysm presence and development. Life Sci. 2021, 274, 119346. [Google Scholar] [CrossRef]
- Daugherty, A.; Manning, M.W.; Cassis, L.A. Angiotensin II promotes atherosclerotic lesions and aneurysms in apolipoprotein E-deficient mice. J. Clin. Investig. 2000, 105, 1605–1612. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Control (n = 36) | AAA (n = 70) | p-Value a | p-Value b |
|---|---|---|---|---|
| Age (years) | 70.11 ± 8.67 | 70.37 ± 8.59 | 0.88 | - |
| Gender (M/F) | 33/3 | 61/9 | 0.75 | - |
| BMI (Kg/m2) | 25.3 ± 4.01 | 24.36 ± 3.49 | 0.214 | - |
| Hypertension | 17 (47.2%) | 55 (78.6%) | *** (0.002) | *** (0.000) |
| Diabetes | 12 (33.3%) | 11 (15.7%) | * (0.048) | 0.057 |
| CAD | 1 (0.03%) | 20 (28.6%) | *** (0.0015) | *** (0.001) |
| Glucose (mmol/L) | 5.60 (1.68) | 5.00 (1.15) | *** (0.002) | * (0.021) |
| HbA1c (%) | 5.7 (1.05) | 5.85 (1.48) | - | - |
| HCY (μmol/L) | 13.5 (2.8) | 15.2 (7.7) | * (0.029) | * (0.016) |
| TC (mmol/L) | 4.76 ± 0.76 | 4.37 ± 1.28 | - | * (0.023) |
| TG (mmol/L) | 1.26 (1.24) | 1.21 (1.29) | - | - |
| HDL-c (mmol/L) | 1.14 ± 0.29 | 0.98 ± 0.24 | *** (0.004) | *** (0.000) |
| LDL-c (mmol/L) | 2.91 ± 0.79 | 2.79 ± 1.00 | - | - |
| hsCRP (mg/L) | 1.05 (0.52) | 1.85 (3.64) | * (0.022) | * (0.048) |
| Urea (mmol/L) | 5.48 (2.09) | 6.32 (3.26) | - | - |
| UA (μmol/L) | 322.03 ± 69.30 | 368.27 ± 76.79 | ** (0.004) | *** (0.000) |
| Cr(E) (μmol/L) | 83 (20.75) | 85 (31) | - | - |
| WBC (109/L) | 6.08 (2.12) | 6.38 (2.33) | NS | NS |
| Platelet (109/L) | 213.03 ± 59.05 | 185.4 ± 64.2 | * (0.034) | * (0.029) |
| Monocyte (109/L) | 0.355 (0.13) | 0.39 (0.22) | * (0.03) | ** (0.012) |
| Neutrophil (109/L) | 3.82 (1.68) | 3.89 (1.81) | - | - |
| Lymphocyte (109/L) | 1.78 ± 0.60 | 1.56 ± 0.56 | - | - |
| NLR | 2.09 (1.21) | 2.47 (1.66) | - | - |
| Rank | Lipids | VIP Score | Fold Change |
|---|---|---|---|
| 1 | LysoPC(16:0) | 1.955664231 | 0.60829 |
| 2 | LysoPC(18:0) | 1.938433674 | 0.58536 |
| 3 | Linoelaidyl carnitine | 1.933833948 | 0.43217 |
| 4 | LysoPC(17:0) | 1.679655063 | 0.66623 |
| 5 | Quinone | 1.656635156 | 1.6062 |
| 6 | LysoPC(15:0) | 1.597648917 | 0.77493 |
| 7 | LysoPE(0:0/20:0) | 1.581730908 | 0.66221 |
| 8 | LysoPC(P-18:0) | 1.535502986 | 0.67357 |
| 9 | Oleoyl L-carnitine | 1.395659551 | 0.58249 |
| 10 | LysoPC(18:1(11Z)) | 1.364155556 | 0.62939 |
| 11 | Arachidyl carnitine | 1.34257126 | 1.1618 |
| 12 | DG(24:0/0:0/18:3n3) | 1.341787518 | 1.2566 |
| 13 | PC(22:6(4Z,7Z,10Z,13Z,16Z,19Z)/18:2(9Z,12Z)) | 1.323787935 | 0.68801 |
| 14 | LysoPC(18:1(9Z)) | 1.323317442 | 0.75435 |
| 15 | CE(18:1(11Z)) | 1.317468625 | 1.4777 |
| 16 | LysoPC(16:1(9Z)) | 1.287512004 | 0.66547 |
| 17 | PC(16:0/18:2(9Z,12Z)) | 1.23655766 | 1.2012 |
| 18 | Heptadecanoyl carnitine | 1.221290467 | 1.134 |
| 19 | L-Carnitine | 1.219465979 | 1.3779 |
| 20 | PE(18:0/18:1(9Z)) | 1.172025083 | 0.64375 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, T.; Lei, C.; Song, W.; Wu, X.; Wu, J.; Li, F.; Lv, Y.; Chen, Y.; Liu, B.; Zheng, Y. Plasma Lipidomics Analysis Reveals the Potential Role of Lysophosphatidylcholines in Abdominal Aortic Aneurysm Progression and Formation. Int. J. Mol. Sci. 2023, 24, 10253. https://doi.org/10.3390/ijms241210253
Xie T, Lei C, Song W, Wu X, Wu J, Li F, Lv Y, Chen Y, Liu B, Zheng Y. Plasma Lipidomics Analysis Reveals the Potential Role of Lysophosphatidylcholines in Abdominal Aortic Aneurysm Progression and Formation. International Journal of Molecular Sciences. 2023; 24(12):10253. https://doi.org/10.3390/ijms241210253
Chicago/Turabian StyleXie, Ting, Chuxiang Lei, Wei Song, Xunyao Wu, Jianqiang Wu, Fangyuan Li, Yanze Lv, Yuexin Chen, Bao Liu, and Yuehong Zheng. 2023. "Plasma Lipidomics Analysis Reveals the Potential Role of Lysophosphatidylcholines in Abdominal Aortic Aneurysm Progression and Formation" International Journal of Molecular Sciences 24, no. 12: 10253. https://doi.org/10.3390/ijms241210253