
A Network Pharmacology and Multi-Omics Combination Approach to Reveal the Effect of Strontium on Ca2+ Metabolism in Bovine Rumen Epithelial Cells
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of Sr on Viability of Rumen Epithelial Cells
2.2. Effect of Sr on Intracellular Ca2+ Level in Rumen Epithelial Cells
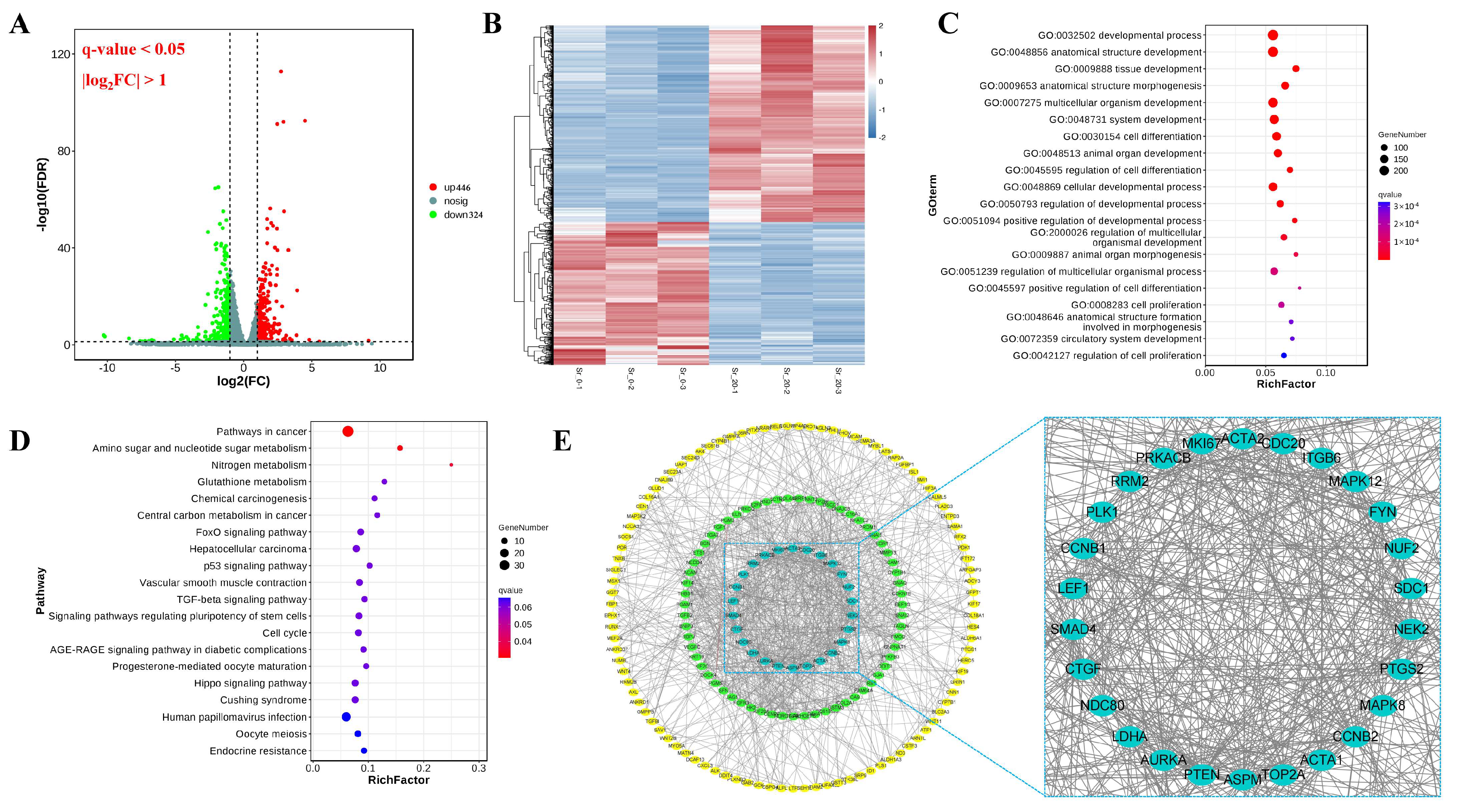
2.3. Analysis of Differentially Expressed Genes (DEGs)
2.4. Analysis of Differentially Expressed Proteins (DEPs)
2.5. Association Analysis of the Transcriptome and Proteome
2.6. Networks and Enriched Functions in Ca2+ Metabolism-Associated Genes
2.7. RT-PCR Analysis of the Effect of Sr on Targets Changes
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of Primary Bovine Rumen Epithelial Cells
4.2. Identification of Primary Bovine Rumen Epithelial Cells
4.3. LDH Activities Analysis
4.4. Cell Viability Analysis
4.5. Cell Cycle Analysis
4.6. Intracellular Ca2+ Analysis
4.7. RNA Extraction and RNA-Sequencing
4.8. Protein Extraction and DIA Labelling
4.9. Identification of Ca2+ Metabolism-Related Targets
4.10. Real-Time Polymerase Chain Reaction (RT-PCR)
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peng, Y.; Ma, F.; Hu, L.; Deng, Y.; He, W.; Tang, B. Strontium based Astragalus polysaccharides promote osteoblasts differentiation and mineralization. Int. J. Biol. Macromol. 2022, 205, 761–771. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Zhang, Y.; Zhou, Y. Strontium Functionalized in Biomaterials for Bone Tissue Engineering: A Prominent Role in Osteoimmunomodulation. Front. Bioeng. Biotechnol. 2022, 10, 928799. [Google Scholar] [CrossRef] [PubMed]
- Cheshmedzhieva, D.; Ilieva, S.; Permyakov, E.A.; Permyakov, S.E.; Dudev, T. Ca2+/Sr2+ Selectivity in Calcium-Sensing Receptor (CaSR): Implications for Strontium’s Anti-Osteoporosis Effect. Biomolecules 2021, 11, 1576. [Google Scholar] [CrossRef]
- Petar Jovanović, S.R.; Roitman, N.; Erel, R. Strontium as a tracer for calcium: Uptake, transport and partitioning within tomato plants. Plant Soil. 2021, 466, 303–316. [Google Scholar] [CrossRef]
- Vologzhannikova, A.A.; Shevelyova, M.P.; Kazakov, A.S.; Sokolov, A.S.; Borisova, N.I.; Permyakov, E.A.; Kircheva, N.; Nikolova, V.; Dudev, T.; Permyakov, S.E. Strontium Binding to alpha-Parvalbumin, a Canonical Calcium-Binding Protein of the “EF-Hand” Family. Biomolecules 2021, 11, 1158. [Google Scholar] [CrossRef]
- Wu, Q.; Hu, L.; Yan, R.; Shi, J.; Gu, H.; Deng, Y.; Jiang, R.; Wen, J.; Jiang, X. Strontium-incorporated bioceramic scaffolds for enhanced osteoporosis bone regeneration. Bone Res. 2022, 10, 55. [Google Scholar] [CrossRef]
- Liu, S.; Shen, B.; Loor, J.J.; Jiang, Q.; Yuan, Y.; Kong, Y.; Tan, P.; Zeng, F.; Zhao, C.; Zhu, X.; et al. Strontium Regulates the Proliferation and Differentiation of Isolated Primary Bovine Chondrocytes via the TGFβ/SMAD Pathway. Front. Pharmacol. 2022, 13, 925302. [Google Scholar] [CrossRef]
- Xie, H.; Gu, Z.; He, Y.; Xu, J.; Xu, C.; Li, L.; Ye, Q. Microenvironment construction of strontium-calcium-based biomaterials for bone tissue regeneration: The equilibrium effect of calcium to strontium. J. Mater. Chem. B 2018, 6, 2332–2339. [Google Scholar] [CrossRef]
- Meo, S.C.; Yamazaki, W.; Leal, C.L.V.; de Oliveira, J.A.; Garcia, J.M. Use of strontium for bovine oocyte activation. Theriogenology. 2005, 63, 2089–2102. [Google Scholar] [CrossRef]
- Coulombe, J.; Faure, H.; Robin, B.; Ruat, M. In vitro effects of strontium ranelate on the extracellular calcium-sensing receptor. Biochem. Biophys. Res. Commun. 2004, 323, 1184–1190. [Google Scholar] [CrossRef] [PubMed]
- Nedic, S.; Palamarevic, M.; Arsic, S.; Jovanovic, L.; Prodanovic, R.; Kirovski, D.; Vujanac, I. Parathyroid hormone response in treatment of subclinical hypocalcemia in postpartum dairy cows. Res. Vet. Sci. 2020, 132, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Colazo, M.G.; De Buck, J. Development of a blood calcium test for hypocalcemia diagnosis in dairy cows. Res. Vet. Sci. 2022, 147, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Barraclough, R.A.C.; Shaw, D.J.; Thorup, V.M.; Haskell, M.J.; Lee, W.; Macrae, A.I. The behavior of dairy cattle in the transition period: Effects of blood calcium status. J. Dairy. Sci. 2020, 103, 10604–10613. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, S.J.; Huzzey, J.M.; Kuhn-Sherlock, B.; Turner, S.A.; Mueller, K.R.; Phyn, C.V.C.; Donaghy, D.J.; Roche, J.R. Associations between lying behavior and activity and hypocalcemia in grazing dairy cows during the transition period. J. Dairy Sci. 2020, 103, 10530–10546. [Google Scholar] [CrossRef] [PubMed]
- Serrenho, R.C.; DeVries, T.J.; Duffield, T.F.; LeBlanc, S.J. Graduate Student Literature Review: What do we know about the effects of clinical and subclinical hypocalcemia on health and performance of dairy cows? J. Dairy Sci. 2021, 104, 6304–6326. [Google Scholar] [CrossRef]
- Hernandez-Castellano, L.E.; Hernandez, L.L.; Bruckmaier, R.M. Review: Endocrine pathways to regulate calcium homeostasis around parturition and the prevention of hypocalcemia in periparturient dairy cows. Animal 2020, 14, 330–338. [Google Scholar] [CrossRef]
- Wadhwa, D.R.; Care, A.D. Effects of strontium on the absorption of calcium, magnesium and phosphate ions from the ovine reticulo-rumen. J. Comp. Physiol. B 2000, 170, 225–229. [Google Scholar] [CrossRef]
- Wilkens, M.R.; Mrochen, N.; Breves, G.; Schroder, B. Gastrointestinal calcium absorption in sheep is mostly insensitive to an alimentary induced challenge of calcium homeostasis. Comp. Biochem. Phys. B 2011, 158, 199–207. [Google Scholar] [CrossRef]
- Hyde, M.L.; Fraser, D.R. In vivo measurement of the absorption of strontium in the rumen and small intestine of sheep as an index of calcium absorption capacity. Br. J. Nutr. 2014, 112, 718–724. [Google Scholar] [CrossRef]
- Kolodziejska, B.; Stepien, N.; Kolmas, J. The Influence of Strontium on Bone Tissue Metabolism and Its Application in Osteoporosis Treatment. Int. J. Mol. Sci. 2021, 22, 6564. [Google Scholar] [CrossRef]
- Dijkgraaf-ten Bolscher, M.; Netelenbos, J.C.; Barto, R.; van der Vijgh, W.J.F. Strontium as a marker for intestinal calcium absorption: The stimulatory effect of calcitriol. Clin. Chem. 2000, 46, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Vezzoli, G.; Baragetti, I.; Zerbi, S.; Caumo, A.; Soldati, L.; Bellinzoni, P.; Centemero, A.; Rubinacci, A.; Moro, G.; Bianchi, G. Strontium absorption and excretion in normocalciuric subjects: Relation to calcium metabolism. Clin. Chem. 1998, 44, 586–590. [Google Scholar] [CrossRef]
- Hyde, M.L.; Wilkens, M.R.; Fraser, D.R. In vivo measurement of strontium absorption from the rumen of dairy cows as an index of calcium absorption capacity. J. Dairy Sci. 2019, 102, 5699–5705. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Kondo, N.; Takagi, K.; Nishikawa, A.; Murakami, Y.; Otsuka, M.; Tsukano, K.; Ikeda, K.; Funakura, H.; Yasutomi, I.; et al. Validation of the bovine blood calcium checker as a rapid and simple measuring tool for the ionized calcium concentration in cattle. J. Vet. Med. Sci. 2021, 83, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Ceciliani, F.; Lecchi, C.; Urh, C.; Sauerwein, H. Proteomics and metabolomics characterizing the pathophysiology of adaptive reactions to the metabolic challenges during the transition from late pregnancy to early lactation in dairy cows. J. Proteom. 2018, 178, 92–106. [Google Scholar] [CrossRef]
- Tomashov-Matar, R.; Tchetchik, D.; Eldar, A.; Kaplan-Kraicer, R.; Oron, Y.; Shalgi, R. Strontium-induced rat egg activation. Reproduction 2005, 130, 467–474. [Google Scholar] [CrossRef]
- Storey, A.; Elgmati, K.; Wang, Y.; Knaggs, P.; Swann, K. The role of ATP in the differential ability of Sr2+ to trigger Ca2+ oscillations in mouse and human eggs. Mol. Hum. Reprod. 2021, 27, gaaa086. [Google Scholar] [CrossRef]
- Wang, G.; Li, H.; Wang, K.; Yang, J.; Duan, M.; Zhang, J.; Ye, N. Regulation of gene expression involved in the remobilization of rice straw carbon reserves results from moderate soil drying during grain filling. Plant J. 2020, 101, 604–618. [Google Scholar] [CrossRef]
- Coelho, I.; Castanheira, I.; Bordado, J.M.; Donard, O.; Silva, J.A.L. Recent developments and trends in the application of strontium and its isotopes in biological related fields. Trac-Trend Anal. Chem. 2017, 90, 45–61. [Google Scholar] [CrossRef]
- Meyer-Binzegger, M.; Ollagnier, C.; Eggerschwiler, L.; Buhler, K.; Meylan, M.; Schlegel, P. Potential of a rumen bolus containing 1,25-dihydroxyvitamin D3 glycosides for the prevention of hypocalcaemia in primiparous and multiparous dairy cows. Animal 2022, 16, 100414. [Google Scholar] [CrossRef]
- Chuang, G.T.; Liu, P.H.; Chyan, T.W.; Huang, C.H.; Huang, Y.Y.; Lin, C.H.; Lin, J.W.; Hsu, C.N.; Tsai, R.Y.; Hsieh, M.L.; et al. Genome-wide association study for circulating fibroblast growth factor 21 and 23. Sci. Rep. 2020, 10, 14578. [Google Scholar] [CrossRef] [PubMed]
- Rosendahl, J.; Braun, H.S.; Schrapers, K.T.; Martens, H.; Stumpff, F. Evidence for the functional involvement of members of the TRP channel family in the uptake of Na (+) and NH4 (+) by the ruminal epithelium. Pflug. Arch. Eur. J. Phy. 2016, 468, 1333–1352. [Google Scholar] [CrossRef] [PubMed]
- Geiger, S.; Patra, A.K.; Schrapers, K.T.; Braun, H.S.; Aschenbach, J.R. Menthol stimulates calcium absorption in the rumen but not in the jejunum of sheep. J. Dairy Sci. 2021, 104, 3067–3081. [Google Scholar] [CrossRef] [PubMed]
- VanHouten, J.N.; Dann, P.; Stewart, A.F.; Watson, C.J.; Pollak, M.; Karaphs, A.C.; Wysolmerski, J.J. Mammary-specific deletion of parathyroid hormone-related protein preserves bone mass during lactation. J. Clin. Investig. 2003, 112, 1429–1436. [Google Scholar] [CrossRef]
- Bach, F.C.; Rutten, K.; Hendriks, K.; Riemers, F.M.; Cornelissen, P.; de Bruin, A.; Arkesteijn, G.J.; Wubbolts, R.; Horton, W.A.; Penning, L.C.; et al. The Paracrine Feedback Loop Between Vitamin D3 (1,25(OH)2D3) and PTHrP in Prehypertrophic Chondrocytes. J. Cell. Physiol. 2014, 229, 1999–2014. [Google Scholar] [CrossRef]
- Okazaki, T.; Nishimori, S.; Ogata, E.; Fujita, T. Vitamin D-dependent recruitment of DNA-PK to the chromatinized negative vitamin D response element in the PTHrP gene is required for gene repression by vitamin D. Biochem. Bioph Res. Commun. 2003, 304, 632–637. [Google Scholar] [CrossRef]
- Zang, W.J.; Li, H.; Zhang, Z.F.; QuZhen, R.; CuoMu, Y.Z.; Zhang, D.K.; Luo, J.; Loor, J.J.; Zheng, H.L. Serotonin induces parathyroid hormone-related protein in goat mammary gland. J. Anim. Sci. 2018, 96, 1010–1016. [Google Scholar] [CrossRef]
- Zhang, Z.; Du, W.; Liu, W.; Wong, B.T.; Zheng, H. Increasing serotonin concentrations alter calcium metabolism in periparturient dairy goats. J. Anim. Sci. 2022, 100, skac065. [Google Scholar] [CrossRef]
- Kim, J.M.; Lin, C.J.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Yamane, M.; Yamashita, N.; Yamamoto, H.; Iizuka, A.; Shouji, M.; Usui, H.; Goshima, Y. Semaphorin3A facilitates axonal transport through a local calcium signaling and tetrodotoxin-sensitive voltage-gated sodium channels. Biochem. Bioph Res. Commun. 2012, 422, 333–338. [Google Scholar] [CrossRef]
- Treinys, R.; Kaselis, A.; Jover, E.; Bagnard, D.; Satkauskas, S. R-Type Calcium Channels Are Crucial for Semaphorin 3A-Induced DRG Axon Growth Cone Collapse. PloS ONE 2014, 9, e102357. [Google Scholar] [CrossRef] [PubMed]
- Duan, M.; Wang, Q.; Liu, Y.; Xie, J. The role of TGF-beta2 in cartilage development and diseases. Bone Jt. Res. 2021, 10, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Ahn, D.; Park, T.I.; Jeong, J.Y. TGFBI Expression Predicts the Survival of Patients with Oropharyngeal Squamous Cell Carcinoma. In Vivo 2020, 34, 3005–3012. [Google Scholar] [CrossRef]
- Guo, Y.S.; Tang, J.; Chen, B.; Huang, W.; Li, Y.; Cui, H.Y.; Zhang, X.; Wang, S.J.; Chen, Z.N.; Jiang, J.L. βig-h3 regulates store-operated Ca2+ entry and promotes the invasion of human hepatocellular carcinoma cells. Cell Biol. Int. 2011, 35, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Riihonen, R.; Supuran, C.T.; Parkkila, S.; Pastorekova, S.; Vaananen, H.K.; Laitala-Leinonen, T. Membrane-bound carbonic anhydrases in osteoclasts. Bone 2007, 40, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Arlot-Bonnemains, Y.; Fouchereau-Peron, M.; Moukhtar, M.S.; Benson, A.A.; Milhaud, G. Calcium-regulating hormones modulate carbonic anhydrase II in the human erythrocyte. Proc. Natl. Acad. Sci. USA 1985, 82, 8832–8834. [Google Scholar] [CrossRef] [PubMed]
- Lehenkari, P.; Hentunen, T.A.; Laitala-Leinonen, T.; Tuukkanen, J.; Vaananen, H.K. Carbonic anhydrase II plays a major role in osteoclast differentiation and bone resorption by effecting the steady state intracellular pH and Ca2+. Exp. Cell Res. 1998, 242, 128–137. [Google Scholar] [CrossRef]
- Hillstrom Shapiro, L.; Venta, P.J.; Yu, Y.S.; Tashian, R.E. Carbonic anhydrase II is induced in HL-60 cells by 1,25-dihydroxyvitamin D3: A model for osteoclast gene regulation. FEBS Lett. 1989, 249, 307–310. [Google Scholar] [CrossRef]
- Slominski, R.M.; Raman, C.; Elmets, C.; Jetten, A.M.; Slominski, A.T.; Tuckey, R.C. The significance of CYP11A1 expression in skin physiology and pathology. Mol. Cell Endocrinol. 2021, 530, 111238. [Google Scholar] [CrossRef]
- Chaiprasongsuk, A.; Janjetovic, Z.; Kim, T.K.; Tuckey, R.C.; Li, W.; Raman, C.; Panich, U.; Slominski, A.T. CYP11A1-derived vitamin D3 products protect against UVB-induced inflammation and promote keratinocytes differentiation. Free. Radic. Biol. Med. 2020, 155, 87–98. [Google Scholar] [CrossRef]
- Ma, Y.; Gong, Y.J.; Xu, Q.Q.; Zou, X. Molecular mechanism of mercuric chloride inhibiting progesterone secretion in ovarian granulosa cells of laying hens. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1533–1542. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.X.; Wang, Y.Z.; Peng, Z.C.; Sun, X.D.; Sun, G.Q.; Yuan, X.; Li, X.W.; Liu, G.W. Subacute ruminal acidosis suppressed the expression of MCT1 in rumen of cows. J. Cell. Physiol. 2019, 234, 11734–11745. [Google Scholar] [CrossRef]
- Zhao, C.X.; Bobe, G.; Wang, Y.Z.; Zhang, X.Y.; Zhao, Z.B.; Zhang, S.Q.; Sun, G.Q.; Yuan, X.; Li, X.W.; Liu, G.W. Potential Role of SLC5A8 Expression in the Etiology of Subacute Ruminal Acidosis. Front. Vet. Sci. 2020, 7, 394. [Google Scholar] [CrossRef]
- Zhang, H.X.; Birch, J.; Pei, J.J.; Ahmed, I.A.M.; Yang, H.Y.; Dias, G.; Abd El-Aty, A.M.; Bekhit, A.E. Identification of Six Phytochemical Compounds from Asparagus officinalis L. Root Cultivars from New Zealand and China Using UAE-SPE-UPLC-MS/MS: Effects of Extracts on H2O2-Induced Oxidative Stress. Nutrients 2019, 11, 107. [Google Scholar] [CrossRef]
- Ming, J.; Wu, S.L.; You, T.Z.; Wang, X.L.; Yu, C.; Luo, P.; Zhang, A.H.; Pan, X.L. Histone Deacetylation in the Promoter of p16 Is Involved in Fluoride-Induced Human Osteoblast Activation via the Inhibition of Sp1 Binding. Biol. Trace Elem. Res. 2019, 188, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol a | Accession No. | Product Size | Primer Sequence (5′ → 3′) |
|---|---|---|---|
| CYP11A1 | NM_176644.2 | 219 bp | F: CTTGGAGGGACCATGTAGCC R: GCAATGTCATGGATGTCGTGT |
| AHCYL2 | XM_005205707.4 | 279 bp | F: GCACAGTCAAGAAGATCCAATTTGC R: GTGCTGGCATTTCTTGCTCA |
| UPK1B | NM_174482.2 | 181 bp | F: GAGGAGAGGGTAAGCTTGGG R: TGGCTTCAAGCAGTGGGTAG |
| WWC2 | XM_024986463.1 | 190 bp | F: CGCCCGGTTCCCCTATG R: GCTTGGTCAACCTGTCCC |
| TGF-β2 | NM_ 001113252.1 | 264 bp | F: TCATGCGCAAGAGGATCGAG R: GCGGGATGGCATTTTCCGAG |
| SEMA3A | NM_001276701.2 | 224 bp | F: TCTTCCGAACTCTTGGGCAC R: GCCCCCAAAGTCATTCTTGC |
| CA2 | NM_178572.2 | 201 bp | F: TCGCGGAGAATGGTCAACAA R: GTGAACCAGGTGTAGCTCGG |
| TGFBI | NM_001205402.1 | 273 bp | F: GAGCTCTGTGAAACTAGCCCC R: TGGGCTAACCGCCATGTTTA |
| PTHLH | NM_174753.1 | 132 bp | F: GGTTATTATTTCGGAGGAGGCG R: CTCTCGCTCTGGGGACTTAT |
| GAPDH | NM_001034034 | 117 bp | F: CCTGCCAAGTATGATGAGAT R: AGTGTCGCTGTTGAAGTC |
| 18S | NR_036642.1 | 130 bp | F: ACCCATTCGAACGTCTGCCCTATT R: TCCTTGGATGTGGTAGCCGTTTCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, P.; Zhao, C.; Dong, Y.; Zhang, Z.; Mei, L.; Kong, Y.; Zeng, F.; Wen, Y.; Zhao, B.; Wang, J. A Network Pharmacology and Multi-Omics Combination Approach to Reveal the Effect of Strontium on Ca2+ Metabolism in Bovine Rumen Epithelial Cells. Int. J. Mol. Sci. 2023, 24, 9383. https://doi.org/10.3390/ijms24119383
Tan P, Zhao C, Dong Y, Zhang Z, Mei L, Kong Y, Zeng F, Wen Y, Zhao B, Wang J. A Network Pharmacology and Multi-Omics Combination Approach to Reveal the Effect of Strontium on Ca2+ Metabolism in Bovine Rumen Epithelial Cells. International Journal of Molecular Sciences. 2023; 24(11):9383. https://doi.org/10.3390/ijms24119383
Chicago/Turabian StyleTan, Panpan, Chenxu Zhao, Yong Dong, Zixin Zhang, Linshan Mei, Yezi Kong, Fangyuan Zeng, Yongqiang Wen, Baoyu Zhao, and Jianguo Wang. 2023. "A Network Pharmacology and Multi-Omics Combination Approach to Reveal the Effect of Strontium on Ca2+ Metabolism in Bovine Rumen Epithelial Cells" International Journal of Molecular Sciences 24, no. 11: 9383. https://doi.org/10.3390/ijms24119383