
A Comprehensive Look at the -13910 C>T LCT Gene Polymorphism as a Molecular Marker for Vitamin D and Calcium Levels in Young Adults in Central and Eastern Europe: A Preliminary Study

Abstract
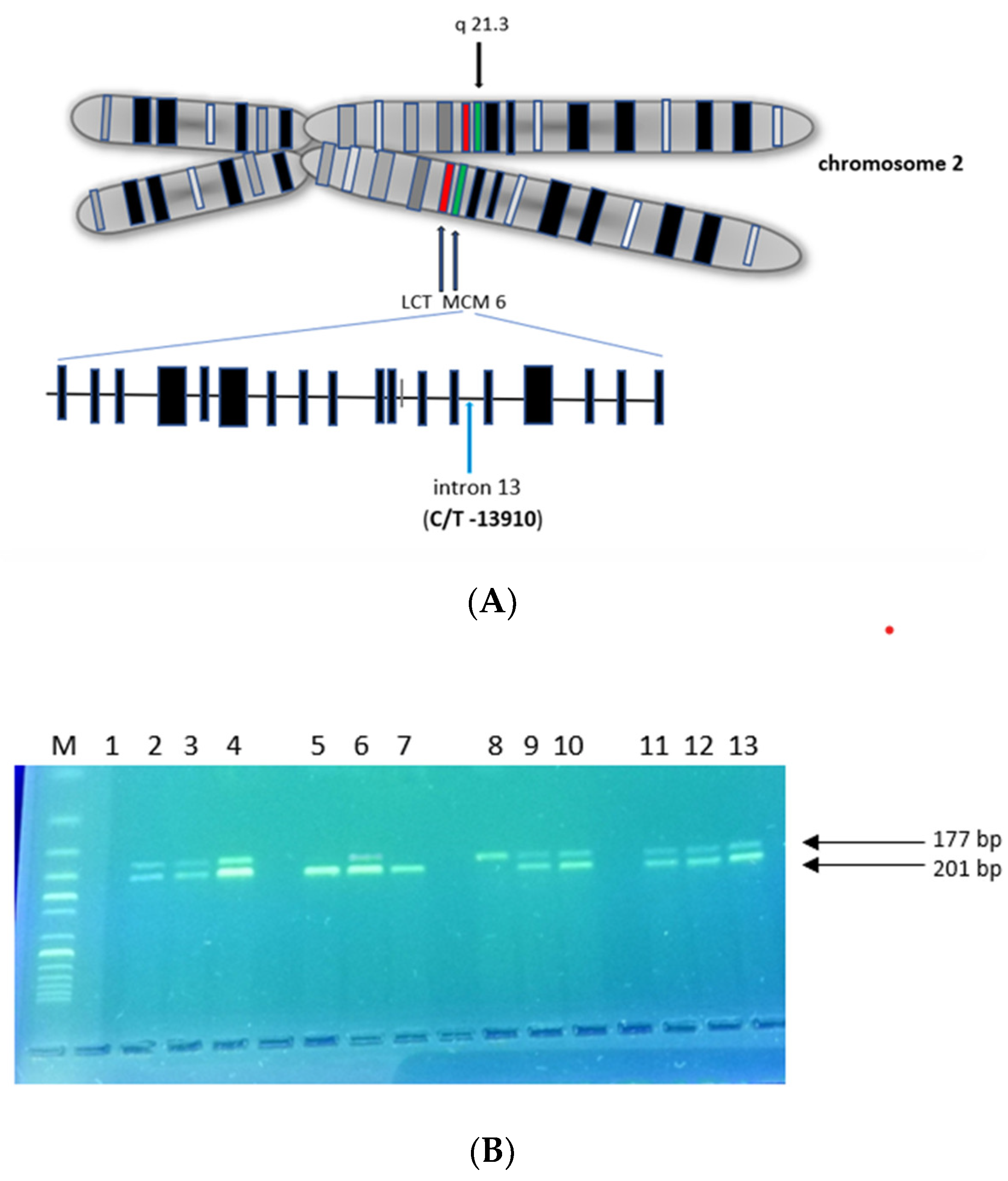
:1. Introduction
- Alactasia, caused by a lack of lactase in the body (rare);
- Secondary lactase deficiency, either transient or chronic, depending on the type and duration of the agent damaging the small intestinal mucosa;
2. Results
3. Discussion
3.1. Potential Differences in Anthropometric Characteristics
3.2. Vitamin D and Calcium Intake vs. Their Serum Concentrations
3.3. Assessment of LCT and VDR Gene Polymorphisms Interactions
Limitations
4. Materials and Methods
4.1. Design of the Study
4.2. Genotypes
- Forward—5′-GCTGGCAATACAGATAAGATAATGGA-3′
- Reverse—5′-CTGCTTTGGTTGAAGCGAAGAT-3′
- F—5′-GGCAACCAGACTACAAGTACC-3′
- R—5′-TCTTCTCACCTCTAACCAGCG-3′
- F—5′-AGCTGGCCCTGGCACTGACTCTGCTCT-3′
- R—5′-ATGGAAACACCTTGCTTCTTCTCCCTC–3′
4.3. Anthropometric Parameters
4.4. Assessment of Nutrition
4.5. Calcium Intake from Dairy Products
4.6. Biochemical Research
4.6.1. Determination of Vitamin D Metabolites
4.6.2. Determination of Calcium
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Płoszaj, T.; Jędrychowska-Dańska, K.; Witas, H. Frequency of Lactase Persistence Genotype in a Healthy Polish Population. Cent. Eur. J. Biol. 2011, 6, 176–179. [Google Scholar] [CrossRef]
- Marasz, A. Frequency and clinical overview of hypolactasia among children, adolescents and students of Szczecin schools. Pomeranian J. Life Sci. 2015, 61, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Tomczonek-Moruś, J.; Wojtasik, A.; Zeman, K.; Smolarz, B.; Bąk-Romaniszyn, L. 13910C>T and 22018G>A LCT Gene Polymorphisms in Diagnosing Hypolactasia in Children. United Eur. Gastroenterol. J. 2019, 7, 210–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mill, D.; Dawson, J.; Johnson, J.L. Managing Acute Pain in Patients Who Report Lactose Intolerance: The Safety of an Old Excipient Re-Examined. Ther. Adv. Drug Saf. 2018, 9, 227–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fassio, F.; Facioni, M.S.; Guagnini, F. Lactose Maldigestion, Malabsorption, and Intolerance: A Comprehensive Review with a Focus on Current Management and Future Perspectives. Nutrients 2018, 10, 1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Facioni, M.S.; Raspini, B.; Pivari, F.; Dogliotti, E.; Cena, H. Nutritional Management of Lactose Intolerance: The Importance of Diet and Food Labelling. J. Transl. Med. 2020, 18, 260. [Google Scholar] [CrossRef]
- Toca, M.D.C.; Fernández, A.; Orsi, M.; Tabacco, O.; Vinderola, G. Lactose Intolerance: Myths and Facts. An Update. Arch. Argent. Pediatr. 2022, 120, 59–66. [Google Scholar] [CrossRef]
- Szilagyi, A. Adaptation to Lactose in Lactase Non Persistent People: Effects on Intolerance and the Relationship between Dairy Food Consumption and Evalution of Diseases. Nutrients 2015, 7, 6751–6779. [Google Scholar] [CrossRef] [Green Version]
- Szilagyi, A. Review Article: Lactose--a Potential Prebiotic. Aliment. Pharmacol. Ther. 2002, 16, 1591–1602. [Google Scholar] [CrossRef]
- Mądry, E.; Ewa, F.; Walkowiak, J. Lactose Intolerance—Current State of Knowledge. Acta Sci. Pol. Technol. Aliment. 2010, 9, 343–350. [Google Scholar]
- Amiri, M.; Diekmann, L.; von Köckritz-Blickwede, M.; Naim, H.Y. The Diverse Forms of Lactose Intolerance and the Putative Linkage to Several Cancers. Nutrients 2015, 7, 7209–7230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porzi, M.; Burton-Pimentel, K.J.; Walther, B.; Vergères, G. Development of Personalized Nutrition: Applications in Lactose Intolerance Diagnosis and Management. Nutrients 2021, 13, 1503. [Google Scholar] [CrossRef] [PubMed]
- Mattar, R.; do Socorro Monteiro, M.; Villares, C.A.; dos Santos, A.F.; Carrilho, F.J. Single Nucleotide Polymorphism C/T-13910, Located Upstream of the Lactase Gene, Associated with Adult-Type Hypolactasia: Validation for Clinical Practice. Clin. Biochem. 2008, 41, 628–630. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.K.; Waud, J.P.; Matthews, S.B. The Molecular Basis of Lactose Intolerance. Sci. Prog. 2005, 88, 157–202. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, T.; D’Angelo, G.; D’Aversa, F.; Campanale, M.C.; Cesario, V.; Montalto, M.; Gasbarrini, A.; Ojetti, V. Lactose Intolerance: From Diagnosis to Correct Management. Eur. Rev. Med. Pharmacol. Sci. 2013, 17 (Suppl. 2), 18–25. [Google Scholar]
- Lukito, W.; Malik, S.G.; Surono, I.S.; Wahlqvist, M.L. From “lactose Intolerance” to “Lactose Nutrition”. Asia Pac. J. Clin. Nutr. 2015, 24 (Suppl. 1), S1–S8. [Google Scholar] [CrossRef]
- Ridefelt, P.; Håkansson, L.D. Lactose Intolerance: Lactose Tolerance Test versus Genotyping. Scand. J. Gastroenterol. 2005, 40, 822–826. [Google Scholar] [CrossRef]
- Deng, Y.; Misselwitz, B.; Dai, N.; Fox, M. Lactose Intolerance in Adults: Biological Mechanism and Dietary Management. Nutrients 2015, 7, 8020–8035. [Google Scholar] [CrossRef] [Green Version]
- Ugidos-Rodríguez, S.; Matallana-González, M.C.; Sánchez-Mata, M.C. Lactose Malabsorption and Intolerance: A Review. Food Funct. 2018, 9, 4056–4068. [Google Scholar] [CrossRef]
- Mądry, E.; Lisowska, A.; Kwiecień, J.; Marciniak, R.; Korzon-Burakowska, A.; Drzymała-Czyż, S.; Mojs, E.; Walkowiak, J. Adult-Type Hypolactasia and Lactose Malabsorption in Poland. Acta Biochim. Pol. 2010, 57, 585–588. [Google Scholar] [CrossRef] [Green Version]
- Catanzaro, R.; Sciuto, M.; Marotta, F. Lactose Intolerance: An Update on Its Pathogenesis, Diagnosis, and Treatment. Nutr. Res. 2021, 89, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Enattah, N.S.; Sahi, T.; Savilahti, E.; Terwilliger, J.D.; Peltonen, L.; Järvelä, I. Identification of a Variant Associated with Adult-Type Hypolactasia. Nat. Genet. 2002, 30, 233–237. [Google Scholar] [CrossRef] [PubMed]
- De Luca, P.; Iaconis, D.; Biffali, E.; Enza, C.; de Magistris, L.; Riegler, G.; Pappalardo, D.; Amato, M.R.; Iardino, P.; Montanino, C.; et al. Development of a Novel SNP Assay to Detect Lactase Persistence Associated Genetic Variants. Mol. Biol. Rep. 2021, 48, 7087–7093. [Google Scholar] [CrossRef] [PubMed]
- Enattah, N.S.; Jensen, T.G.K.; Nielsen, M.; Lewinski, R.; Kuokkanen, M.; Rasinpera, H.; El-Shanti, H.; Seo, J.K.; Alifrangis, M.; Khalil, I.F.; et al. Independent Introduction of Two Lactase-Persistence Alleles into Human Populations Reflects Different History of Adaptation to Milk Culture. Am. J. Hum. Genet. 2008, 82, 57–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ségurel, L.; Bon, C. On the Evolution of Lactase Persistence in Humans. Annu. Rev. Genomics Hum. Genet. 2017, 18, 297–319. [Google Scholar] [CrossRef]
- Hammer, H.F.; Fox, M.R.; Keller, J.; Salvatore, S.; Basilisco, G.; Hammer, J.; Lopetuso, L.; Benninga, M.; Borrelli, O.; Dumitrascu, D.; et al. European Guideline on Indications, Performance, and Clinical Impact of Hydrogen and Methane Breath Tests in Adult and Pediatric Patients: European Association for Gastroenterology, Endoscopy and Nutrition, European Society of Neurogastroenterology and Motility, and European Society for Paediatric Gastroenterology Hepatology and Nutrition Consensus. United Eur. Gastroenterol. J. 2022, 10, 15–40. [Google Scholar] [CrossRef]
- Storhaug, C.L.; Fosse, S.K.; Fadnes, L.T. Country, Regional, and Global Estimates for Lactose Malabsorption in Adults: A Systematic Review and Meta-Analysis. Lancet Gastroenterol. Hepatol. 2017, 2, 738–746. [Google Scholar] [CrossRef] [Green Version]
- Misselwitz, B.; Butter, M.; Verbeke, K.; Fox, M.R. Update on Lactose Malabsorption and Intolerance: Pathogenesis, Diagnosis and Clinical Management. Gut 2019, 68, 2080–2091. [Google Scholar] [CrossRef] [Green Version]
- Itan, Y.; Jones, B.L.; Ingram, C.J.E.; Swallow, D.M.; Thomas, M.G. A Worldwide Correlation of Lactase Persistence Phenotype and Genotypes. BMC Evol. Biol. 2010, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Alonso, N.; Zelzer, S.; Eibinger, G.; Herrmann, M. Vitamin D Metabolites: Analytical Challenges and Clinical Relevance. Calcif. Tissue Int. 2023, 112, 158–177. [Google Scholar] [CrossRef]
- Nikolac Gabaj, N.; Unic, A.; Miler, M.; Pavicic, T.; Culej, J.; Bolanca, I.; Herman Mahecic, D.; Milevoj Kopcinovic, L.; Vrtaric, A. In Sickness and in Health: Pivotal Role of Vitamin D. Biochem. Med. 2020, 30, 020501. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shu, B.; Li, C.-G.; Xie, X.-W.; Liang, D.; Chen, B.-L.; Lin, X.-C.; Wei, X.; Wang, L.; Leng, X.-Y.; et al. Polymorphisms of Genes Related to Vitamin D Metabolism and Transportation and Its Relationship with the Risk of Osteoporosis: Protocol for a Multicentre Prospective Cohort Study in China. BMJ Open 2019, 9, e028084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ammar, M.; Heni, S.; Tira, M.S.; Khalij, Y.; Hamdouni, H.; Amor, D.; Ksibi, S.; Omezzine, A.; Bouslama, A. Variability in Response to Vitamin D Supplementation According to Vitamin D Metabolism Related Gene Polymorphisms in Healthy Adults. Eur. J. Clin. Nutr. 2023, 77, 189–194. [Google Scholar] [CrossRef]
- Abouzid, M.; Kruszyna, M.; Burchardt, P.; Kruszyna, Ł.; Główka, F.K.; Karaźniewicz-Łada, M. Vitamin D Receptor Gene Polymorphism and Vitamin D Status in Population of Patients with Cardiovascular Disease—A Preliminary Study. Nutrients 2021, 13, 3117. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Long, S.R.; Li, X.; Ge, H.; Liu, X.; Wang, T.; Yu, F.; Wang, Y.; Xue, Y.; Li, W. Serum Vitamin D Deficiency and Vitamin D Receptor Gene Polymorphism Are Associated with Increased Risk of Cardiovascular Disease in a Chinese Rural Population. Nutr. Res. 2019, 61, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Seamans, K.M.; Cashman, K.D. Existing and Potentially Novel Functional Markers of Vitamin D Status: A Systematic Review. Am. J. Clin. Nutr. 2009, 89, 1997S–2008S. [Google Scholar] [CrossRef] [Green Version]
- Ross, A.C.; Manson, J.E.; Abrams, S.A.; Aloia, J.F.; Brannon, P.M.; Clinton, S.K.; Durazo-Arvizu, R.A.; Gallagher, J.C.; Gallo, R.L.; Jones, G.; et al. The 2011 Report on Dietary Reference Intakes for Calcium and Vitamin D from the Institute of Medicine: What Clinicians Need to Know. J. Clin. Endocrinol. Metab. 2011, 96, 53–58. [Google Scholar] [CrossRef]
- Lips, P.; Cashman, K.D.; Lamberg-Allardt, C.; Bischoff-Ferrari, H.A.; Obermayer-Pietsch, B.; Bianchi, M.L.; Stepan, J.; El-Hajj Fuleihan, G.; Bouillon, R. Current Vitamin D Status in European and Middle East Countries and Strategies to Prevent Vitamin D Deficiency: A Position Statement of the European Calcified Tissue Society. Eur. J. Endocrinol. 2019, 180, P23–P54. [Google Scholar] [CrossRef] [Green Version]
- Järvelä, I.; Torniainen, S.; Kolho, K.-L. Molecular Genetics of Human Lactase Deficiencies. Ann. Med. 2009, 41, 568–575. [Google Scholar] [CrossRef]
- Home—SNP—NCBI. Available online: https://www.ncbi.nlm.nih.gov/snp/ (accessed on 11 May 2023).
- Nowak, J.K.; Dybska, E.; Dworacka, M.; Tsikhan, N.; Kononets, V.; Bermagambetova, S.; Walkowiak, J. Ileal Lactase Expression Associates with Lactase Persistence Genotypes. Nutrients 2021, 13, 1340. [Google Scholar] [CrossRef]
- Labrie, V.; Buske, O.J.; Oh, E.; Jeremian, R.; Ptak, C.; Gasiūnas, G.; Maleckas, A.; Petereit, R.; Žvirbliene, A.; Adamonis, K.; et al. Lactase Non-Persistence Is Directed by DNA Variation-Dependent Epigenetic Aging. Nat. Struct. Mol. Biol. 2016, 23, 566–573. [Google Scholar] [CrossRef] [Green Version]
- Alharbi, O.; El-Sohemy, A. Lactose Intolerance (LCT-13910C>T) Genotype Is Associated with Plasma 25-Hydroxyvitamin D Concentrations in Caucasians: A Mendelian Randomization Study. J. Nutr. 2017, 147, 1063–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Costanzo, M.; Berni Canani, R. Lactose Intolerance: Common Misunderstandings. Ann. Nutr. Metab. 2018, 73 (Suppl. 4), 30–37. [Google Scholar] [CrossRef] [PubMed]
- Corgneau, M.; Scher, J.; Ritie-Pertusa, L.; Le, D.T.L.; Petit, J.; Nikolova, Y.; Banon, S.; Gaiani, C. Recent Advances on Lactose Intolerance: Tolerance Thresholds and Currently Available Answers. Crit. Rev. Food Sci. Nutr. 2017, 57, 3344–3356. [Google Scholar] [CrossRef] [PubMed]
- Lomer, M.C.E.; Parkes, G.C.; Sanderson, J.D. Review Article: Lactose Intolerance in Clinical Practice--Myths and Realities. Aliment. Pharmacol. Ther. 2008, 27, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Sun, J.; Lu, Y.; Xu, F.; Li, D.; Jiang, F.; Wan, Z.; Li, X.; Qin, L.-Q.; Larsson, S.C. Health Effects of Milk Consumption: Phenome-Wide Mendelian Randomization Study. BMC Med. 2022, 20, 455. [Google Scholar] [CrossRef]
- Chin, E.L.; Huang, L.; Bouzid, Y.Y.; Kirschke, C.P.; Durbin-Johnson, B.; Baldiviez, L.M.; Bonnel, E.L.; Keim, N.L.; Korf, I.; Stephensen, C.B.; et al. Association of Lactase Persistence Genotypes (Rs4988235) and Ethnicity with Dairy Intake in a Healthy U.S. Population. Nutrients 2019, 11, 1860. [Google Scholar] [CrossRef] [Green Version]
- Vitellio, P.; Celano, G.; Bonfrate, L.; Gobbetti, M.; Portincasa, P.; De Angelis, M. Effects of Bifidobacterium Longum and Lactobacillus Rhamnosus on Gut Microbiota in Patients with Lactose Intolerance and Persisting Functional Gastrointestinal Symptoms: A Randomised, Double-Blind, Cross-Over Study. Nutrients 2019, 11, 886. [Google Scholar] [CrossRef] [Green Version]
- Mnich, B.; Spinek, A.E.; Chyleński, M.; Sommerfeld, A.; Dabert, M.; Juras, A.; Szostek, K. Analysis of LCT-13910 Genotypes and Bone Mineral Density in Ancient Skeletal Materials. PLoS ONE 2018, 13, e0194966. [Google Scholar] [CrossRef] [Green Version]
- Popadowska, A.; Kempińska-Podhorodecka, A. LCT-13910 C>T gene polymorphism and obesity in women. Med. Śr. Environ. Med. 2012, 2, 48–54. [Google Scholar]
- Popadowska, A.; Kempinska-Podhorodecka, A. Relation of the C/T-13910 LCT Polymorphism with Body Composition Measures and Their Modulation by Dairy Products in a Caucasian Men. Am. J. Men’s Health 2021, 15, 15579883211007272. [Google Scholar] [CrossRef] [PubMed]
- Setty-Shah, N.; Maranda, L.; Candela, N.; Fong, J.; Dahod, I.; Rogol, A.D.; Nwosu, B.U. Lactose Intolerance: Lack of Evidence for Short Stature or Vitamin D Deficiency in Prepubertal Children. PLoS ONE 2013, 8, e78653. [Google Scholar] [CrossRef] [PubMed]
- Almon, R.; Sjöström, M.; Nilsson, T.K. Lactase Non-Persistence as a Determinant of Milk Avoidance and Calcium Intake in Children and Adolescents. J. Nutr. Sci. 2013, 2, e26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Beer, H. Dairy Products and Physical Stature: A Systematic Review and Meta-Analysis of Controlled Trials. Econ. Hum. Biol. 2012, 10, 299–309. [Google Scholar] [CrossRef]
- Angelin, T.C.; Bardosono, S.; Shinta, D.; Fahmida, U. Growth, Dietary Intake, and Vitamin D Receptor (VDR) Promoter Genotype in Indonesian School-Age Children. Nutrients 2021, 13, 2904. [Google Scholar] [CrossRef]
- Jarosz, M.; Rychlik, E.; Stoś, K.; Charzewska, J. Normy Żywienia Dla Populacji Polski i Ich Zastosowanie; Nutrition Standards for the Polish Population; Narodowy Instytut Zdrowia Publicznego-Państwowy Zakład Higieny: Warsaw, Poland, 2020.
- Płudowski, P.; Kos-Kudła, B.; Walczak, M.; Fal, A.; Zozulińska-Ziółkiewicz, D.; Sieroszewski, P.; Peregud-Pogorzelski, J.; Lauterbach, R.; Targowski, T.; Lewiński, A.; et al. Guidelines for Preventing and Treating Vitamin D Deficiency: A 2023 Update in Poland. Nutrients 2023, 15, 695. [Google Scholar] [CrossRef]
- Petrenya, N.; Lamberg-Allardt, C.; Melhus, M.; Broderstad, A.R.; Brustad, M. Vitamin D Status in a Multi-Ethnic Population of Northern Norway: The SAMINOR 2 Clinical Survey. Public Health Nutr. 2020, 23, 1186–1200. [Google Scholar] [CrossRef] [Green Version]
- Hodges, J.K.; Cao, S.; Cladis, D.P.; Weaver, C.M. Lactose Intolerance and Bone Health: The Challenge of Ensuring Adequate Calcium Intake. Nutrients 2019, 11, 718. [Google Scholar] [CrossRef] [Green Version]
- Obermayer-Pietsch, B.M.; Bonelli, C.M.; Walter, D.E.; Kuhn, R.J.; Fahrleitner-Pammer, A.; Berghold, A.; Goessler, W.; Stepan, V.; Dobnig, H.; Leb, G.; et al. Genetic Predisposition for Adult Lactose Intolerance and Relation to Diet, Bone Density, and Bone Fractures. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2004, 19, 42–47. [Google Scholar] [CrossRef]
- Rana, S.; Morya, R.K.; Malik, A.; Bhadada, S.K.; Sachdeva, N.; Sharma, G. A Relationship between Vitamin D, Parathyroid Hormone, Calcium Levels and Lactose Intolerance in Type 2 Diabetic Patients and Healthy Subjects. Clin. Chim. Acta Int. J. Clin. Chem. 2016, 462, 174–177. [Google Scholar] [CrossRef]
- Stefano, M.D.; Veneto, G.; Malservisi, S.; Cecchetti, L.; Minguzzi, L.; Strocchi, A.; Corazza, G.R. Lactose Malabsorption and Intolerance and Peak Bone Mass. Gastroenterology 2002, 122, 1793–1799. [Google Scholar] [CrossRef] [PubMed]
- Jasielska, M.; Grzybowska-Chlebowczyk, U. Hypocalcemia and Vitamin D Deficiency in Children with Inflammatory Bowel Diseases and Lactose Intolerance. Nutrients 2021, 13, 2583. [Google Scholar] [CrossRef] [PubMed]
- Koek, W.N.H.; van Meurs, J.B.; van der Eerden, B.C.J.; Rivadeneira, F.; Zillikens, M.C.; Hofman, A.; Obermayer-Pietsch, B.; Lips, P.; Pols, H.A.; Uitterlinden, A.G.; et al. The T-13910C Polymorphism in the Lactase Phlorizin Hydrolase Gene Is Associated with Differences in Serum Calcium Levels and Calcium Intake. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2010, 25, 1980–1987. [Google Scholar] [CrossRef] [PubMed]
- Yahya, N.F.S.; Daud, N.M.; Makbul, I.A.A.; Aziz, Q.A.S.A. Association of Calcium Intake, Lactose Intolerance and Physical Activity with Bone Health Assessed via Quantitative Ultrasound among Young Adults of a Malaysian University. Arch. Osteoporos. 2021, 16, 14. [Google Scholar] [CrossRef]
- Pike, J.W.; Meyer, M.B. The Vitamin D Receptor: New Paradigms for the Regulation of Gene Expression by 1,25-Dihydroxyvitamin D3. Endocrinol. Metab. Clin. N. Am. 2010, 39, 255–269. [Google Scholar] [CrossRef] [Green Version]
- Barry, E.L.; Rees, J.R.; Peacock, J.L.; Mott, L.A.; Amos, C.I.; Bostick, R.M.; Figueiredo, J.C.; Ahnen, D.J.; Bresalier, R.S.; Burke, C.A.; et al. Genetic Variants in CYP2R1, CYP24A1, and VDR Modify the Efficacy of Vitamin D3 Supplementation for Increasing Serum 25-Hydroxyvitamin D Levels in a Randomized Controlled Trial. J. Clin. Endocrinol. Metab. 2014, 99, E2133–E2137. [Google Scholar] [CrossRef] [Green Version]
- Wysoczańska-Klaczyńska, A.; Ślęzak, A.; Hetman, M.; Barg, E. The impact of VDR gene polymorphisms on obesity, metabolic changes, bone mass disorders and neoplastic processes. Pediatr. Endocrinol. Diabetes Metab. 2018, 24, 96–105. [Google Scholar] [CrossRef]
- Usategui-Martín, R.; De Luis-Román, D.-A.; Fernández-Gómez, J.M.; Ruiz-Mambrilla, M.; Pérez-Castrillón, J.-L. Vitamin D Receptor (VDR) Gene Polymorphisms Modify the Response to Vitamin D Supplementation: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 360. [Google Scholar] [CrossRef]
- Serrano, J.C.E.; De Lorenzo, D.; Cassanye, A.; Martín-Gari, M.; Espinel, A.; Delgado, M.A.; Pamplona, R.; Portero-Otin, M. Vitamin D Receptor BsmI Polymorphism Modulates Soy Intake and 25-Hydroxyvitamin D Supplementation Benefits in Cardiovascular Disease Risk Factors Profile. Genes Nutr. 2013, 8, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Divanoglou, N.; Komninou, D.; Stea, E.A.; Argiriou, A.; Papatzikas, G.; Tsakalof, A.; Pazaitou-Panayiotou, K.; Georgakis, M.K.; Petridou, E. Association of Vitamin D Receptor Gene Polymorphisms with Serum Vitamin D Levels in a Greek Rural Population (Velestino Study). Lifestyle Genom. 2021, 14, 81–90. [Google Scholar] [CrossRef]
- Santos, B.R.; Mascarenhas, L.P.G.; Satler, F.; Boguszewski, M.C.S.; Spritzer, P.M. Vitamin D Deficiency in Girls from South Brazil: A Cross-Sectional Study on Prevalence and Association with Vitamin D Receptor Gene Variants. BMC Pediatr. 2012, 12, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobayashi, F.; Lourenço, B.H.; Cardoso, M.A. 25-Hydroxyvitamin D3 Levels, BsmI Polymorphism and Insulin Resistance in Brazilian Amazonian Children. Int. J. Mol. Sci. 2015, 16, 12531–12546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valtueña, J.; González-Gross, M.; Huybrechts, I.; Breidenassel, C.; Ferrari, M.; Mouratidou, T.; Gottrand, F.; Dallongeville, J.; Azzini, E.; Sioen, I.; et al. Factors Associated with Vitamin D Deficiency in European Adolescents: The HELENA Study. J. Nutr. Sci. Vitaminol. 2013, 59, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Jakubowska-Pietkiewicz, E.; Fendler, W.; Porczyński, M.; Chlebna-Sokół, D. Vitamin D receptor gene (BsmI and FokI) polymorphism and calcium-phoshporus indices in children from Lodz region. Pediatr. Endocrinol. 2016, 15, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Szilagyi, A.; Ishayek, N. Lactose Intolerance, Dairy Avoidance, and Treatment Options. Nutrients 2018, 10, 1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva, A.T.; de Lima, J.J.; Reis, P.; Passos, M.; Baumgartner, C.G.; Sereno, A.B.; Krüger, C.C.H.; Cândido, L.M.B. Application of Lactose-Free Whey Protein to Greek Yogurts: Potential Health Benefits and Impact on Rheological Aspects and Sensory Attributes. Foods 2022, 11, 3861. [Google Scholar] [CrossRef] [PubMed]
- Silanikove, N.; Leitner, G.; Merin, U. The Interrelationships between Lactose Intolerance and the Modern Dairy Industry: Global Perspectives in Evolutional and Historical Backgrounds. Nutrients 2015, 7, 7312–7331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seremak-Mrozikiewicz, A.; Drews, K.; Mrozikiewicz, P.M.; Bartkowiak-Wieczorek, J.; Marcinkowska, M.; Wawrzyniak, A.; Slomski, R.; Kalak, R.; Czerny, B.; Horst-Sikorska, W. Correlation of Vitamin D Receptor Gene (VDR) Polymorphism with Osteoporotic Changes in Polish Postmenopausal Women. Neuro Endocrinol. Lett. 2009, 30, 540–546. [Google Scholar]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Szymelfejnik, E.J.; Wądołowska, L.; Cichon, R.; Przysławski, J.; Bolesławska, I. Dairy Products Frequency Questionnaire (ADOS-Ca) Calibration for Calcium Intake Evaluation. Pol. J. Food Nutr. Sci. 2006, 15, 229–236. [Google Scholar]
- Turpeinen, U.; Hohenthal, U.; Stenman, U.-H. Determination of 25-Hydroxyvitamin D in Serum by HPLC and Immunoassay. Clin. Chem. 2003, 49, 1521–1524. [Google Scholar] [CrossRef] [PubMed]
- Dembińska-Kieć, A.; Solnica, B.; Naskalski, J. Laboratory Diagnostics with Elements of Clinical Biochemistry; Edra Urban & Partner: Wrocław, Poland, 2017; ISBN 978-83-65625-50-2. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP | Allele/Genotype | Allele Frequency/ Genotypes |
|---|---|---|
| N(%) | ||
| rs4988235 | C/T | 74/52 (58.7/41.3) |
| TT | 10 (15.9) | |
| CT | 32 (50.8) | |
| CC | 21 (33.3) | |
| Parameters | Lactase Non-Persistence LNP (N = 21) | Lactase Persistence LP (N = 42) | p-Value |
|---|---|---|---|
| Male (N %) | 10 (16) | 13 (21) | - |
| Female (N %) | 11 (17) | 29 (46) | - |
| # Age (years) | 23.0 ± 1.50 | 24.0 ± 1.00 | NS |
| # Height (cm) | 174 ± 6.50 | 172 ± 6.50 | NS |
| Weight (kg) | 68.5 ± 12.4 | 67.7 ± 15.9 | NS |
| BMI (kg/m2) | 22.8 ± 2.68 | 22.8 ± 4.63 | NS |
| # Body fat content (%) | 21.4 ± 3.37 | 20.8 ± 6.70 | NS |
| # Bone mass (kg) | 2.40 ± 0.25 | 2.60 ± 0.70 | NS |
| # Body water content (%) | 54.5 ± 3.60 | 55.2 ± 5.20 | NS |
| Parameters | LNP (N = 21) | LP (N = 42) | p-Value |
|---|---|---|---|
| Male (N %) | 10 (16) | 13 (21) | - |
| Female (N %) | 11 (17) | 29 (46) | - |
| # Total energy (kcal/d) | 2033 ± 542 | 2094 ± 499 | NS |
| Protein (g/d) | 85.8 ± 27.4 | 84.1 ± 21.9 | NS |
| Protein (% kcal) | 17.0 ± 2.58 | 16.5 ± 1.69 | NS |
| # Fat (g/d) | 68.8 ± 16.8 | 80.3 ± 29.5 | NS |
| Fat (% kcal) | 32.7 ± 4.42 | 34.3 ± 5.32 | NS |
| Carbohydrate (g/d) | 253 ± 60.3 | 262 ± 66.8 | NS |
| Carbohydrate (% kcal) | 48.9 ± 4.18 | 50.4 ± 5.36 | NS |
| # Vitamin D intake (µg/d) | 2.32 ±0.36 a | 4.57 ± 0.50 b | 0.045 |
| # Calcium intake (mg/d) | 785 ± 97.5 a | 881 ± 128 b | 0.048 |
| Phosphorus intake (mg/d) | 1357 ± 421 | 1566 ± 502 | NS |
| Parameters | LNP (N = 21) | LP (N = 42) | p-Value |
|---|---|---|---|
| Milk (g/d) | 134.7 ± 66.7 a | 342.5 ± 176 b | 0.012 |
| Total dairy products (g/d) | 78.50 ± 36.2 a | 216.3 ± 102 b | 0.008 |
| Parameters | CC (N = 21) | CT (N = 32) | TT (N = 10) | p-Value |
| Median ± IQR (Min–Max) | ||||
| 25(OH)D2 (ng/mL) | 1.42 ± 0.61 | 1.99 ± 0.36 | 1.47 ± 0.22 | NS |
| (1.25–2.40) | (1.20–2.82) | (1.31–2.75) | ||
| 25(OH)D3 (ng/mL) | 17.6 ± 2.19 a | 19.5 ± 2.79 ab | 21.5 ± 3.99 b | 0.025 |
| (8.58–23.9) | (9.26–32.6) | (16.8–43.7) | ||
| Total (D2+D3) (ng/mL) | 18.1 ± 2.91 a | 20.8 ± 2.97 ab | 22.6 ± 4.46 b | 0.022 |
| (9.58–24.8) | (9.28–33.0) | (16.9–44.1) | ||
| Mean values ± SD (Min–Max) | ||||
| * Calcium (mg/dL) | 2.27 ± 0.29 a | 2.56 ± 0.35 ab | 2.62 ± 0.26 b | 0.049 |
| (1.76–2.75) | (1.57–3.17) | (2.28–3.02) | ||
| Variables | Raw Structure Model | |
|---|---|---|
| p-Value | Adjusted OR (95% Cl) | |
| Total vitamin D | ||
| (D2 + D3) in serum | 0.035 | 1.21 (0.92; 1.40) |
| Calcium in serum | 0.022 | 1.15 (0.84; 1.23) |
| Vitamin D intake | 0.040 | 1.11 (1.05; 1.37) |
| Calcium intake | 0.032 | 1.20 (0.98; 1.28) |
| SNP | Allele/Genotype | Allele Frequency/ Genotypes |
|---|---|---|
| N (%) | ||
| BsmI (rs1544410) | G/A | 64/62 (50.8/49.2) |
| GG | 15 (23.8) | |
| GA | 34 (54.0) | |
| AA | 14 (22.2) | |
| FokI (rs2228570) | T/C | 49/77 (38.9/61.1) |
| TT | 12 (19.0) | |
| TC | 25 (39.7) | |
| CC | 26 (41.3) | |
| VDR Genotypes | LNP (N = 21) | LP (N = 42) | p-Value |
|---|---|---|---|
| N (%) | |||
| BsmI | |||
| GG | 6 (28.6) | 9 (21.4) | NS |
| GA | 10 (47.6) | 24 (57.2) | NS |
| AA | 5 (23.8) | 9 (21.4) | NS |
| FokI | |||
| TT | 8 (38.1) | 8 (19.0) | NS |
| TC | 9 (42.9) | 16 (38.1) | NS |
| CC | 4 (19.0) | 18 (42.9) | 0.042 |
| Polymorphism | LNP | LP |
|---|---|---|
| BsmI | 0.05 | 0.85 |
| FokI | 0.23 | 1.41 |
| SNP | LNP | LP | ||
|---|---|---|---|---|
| Total Vitamin D (ng/mL) (Min–Max) | Calcium (mg/dL) (Min–Max) | Total Vitamin D (ng/mL) (Min–Max) | Calcium (mg/dL) (Min–Max) | |
| BsmI | ||||
| GG | 21.6 ± 0.67 a | 2.46 ± 0.26 a | 22.6 ± 3.53 a | 2.50 ± 0.26 |
| (19.3–24.9) | (2.05–2.75) | (9.56–43.7) | (2.21–3.07) | |
| GA | 18.1 ± 1.99 ab | 2.38 ± 0.37 ab | 19.3 ± 2.76 ab | 2.71 ± 0.28 |
| (10.6–23.4) | (1.87–2.75) | (9.26–32.6) | (2.0–3.17) | |
| AA | 10.4 ± 1.25 b | 2.28 ± 0.12 b | 17.0 ± 2.79 b | 2.33 ± 0.32 |
| (9.58–16.8) | (2.11–2.43) | (12.2–23.7) | (1.60–2.70) | |
| p-value | 0.017 | 0.049 | 0.048 | NS |
| FokI | ||||
| CC | 13.2 ± 2.16 a | 2.04 ± 0.06 a | 20.5 ± 5.19 | 2.17 ± 0.26 a |
| (10.4–22.4) | (1.96–2.11) | (9.56–28.7) | (1.57–2.38) | |
| TC | 19.7 ± 1.30 b | 2.52 ± 0.21 b | 21.8 ± 2.36 | 2.55 ± 0.24 b |
| (12.3–24.9) | (2.13–2.75) | (10.0–32.6) | (2.21–3.17) | |
| TT | 17.2 ± 2.29 b | 2.37 ± 0.31 b | 20.0 ± 2.43 | 2.70 ± 0.21 c |
| (9.58–20.8) | (1.87–2.75) | (9.26–43.7) | (2.37–3.07) | |
| p-value | 0.05 | 0.05 | NS | 0.036 |
| Parameters | LNP | LP | |||||
|---|---|---|---|---|---|---|---|
| Total Vitamin D in Serum | |||||||
| <20 ng/mL | >20 <30 ng/mL | p-Value | <20 ng/mL | >20 <30 ng/mL | >30 ng/mL | p-Value * | |
| Weight (kg) | 68.4 ± 10.8 | 68.9 ± 16.9 | NS | 63.5 ± 13.7 | 71.2 ± 17.7 | 69.0 ± 12.6 | NS |
| BMI (kg/m2) | 23.0 ± 2.46 | 22.4 ± 3.38 | NS | 21.5 ± 3.27 | 23.8 ± 5.61 | 22.9 ± 2.53 | NS |
| # Bone mass (kg) | 2.30 ± 0.40 | 2.50 ± 0.25 | NS | 2.35 ± 0.15 | 2.50 ± 0.45 | 2.80 ± 0.55 | NS |
| # Total energy (kcal/d) | 2022 ± 277 | 1638 ± 380 | NS | 1884 ± 453 | 2239 ± 466 | 1933 ± 380 | NS |
| Protein (g/d) | 85.4 ± 15.3 | 64.6 ± 16.1 | NS | 85.1 ± 22.9 | 84.9 ± 22.3 | 72.4 ± 13.5 | NS |
| Fat (g/d) | 70.0 ± 17.3 | 64.1 ± 18.0 | NS | 81.7 ± 14.9 | 79.2 ± 14.1 | 75.9 ± 14.1 | NS |
| Carbohydrate ((g/d) | 258 ± 48.4 | 241 ± 58.0 | NS | 260 ± 67.0 | 266 ± 70.3 | 245 ± 57.1 | NS |
| # Vitamin D intake (µg/d) | 2.23 ± 0.72 a | 3.07 ± 0.51 b | 0.050 | 3.92 ± 1.22 a | 4.37 ± 0.84 ab | 5.13 ± 0.89 b | 0.048 |
| # Calcium intake (mg/d) | 812 ± 133 | 830 ± 65.9 | NS | 899 ± 144 | 915 ± 175 | 1018 ± 173 | NS |
| Phosphorus intake (mg/d) | 1398 ± 369 | 1256 ± 457 | NS | 1528 ± 449 | 1619 ± 570 | 1419 ± 361 | NS |
| Calcium (mg/dL) | 2.28 ± 0.29 | 2.32 ± 0.14 | NS | 2.39 ± 0.38 | 2.57 ± 0.30 | 2.58 ± 0.17 | NS |
| BsmI (N%) | |||||||
| GG | 1 (4.76) | 5 (23.8) | NS | 2 (4.76) | 6 (14.3) | 2 (4.76) | NS |
| GA | 9 (42.9) | 1 (4.76) | NS | 12 (28.6) | 11 (26.2) | 1 (2.38) | NS |
| AA | 5 (23.8) | 0 | - | 4 (9.52) | 2 (4.76) | 0 | NS |
| FokI (N%) | |||||||
| CC | 3 (14.3) | 1 (4.76) | NS | 4 (9.52) | 2 (4.76) | 2 (4.76) | NS |
| TC | 5 (23.8) | 4 (19.0) | NS | 5 (11.9) | 6 (14.3) | 5 (11.9) | NS |
| TT | 7 (33.3) | 1 (4.76) | NS | 1 (2.38) | 15 (35.7) | 2 (4.76) | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalówka, M.; Kosewski, G.; Lipiński, D.; Przysławski, J. A Comprehensive Look at the -13910 C>T LCT Gene Polymorphism as a Molecular Marker for Vitamin D and Calcium Levels in Young Adults in Central and Eastern Europe: A Preliminary Study. Int. J. Mol. Sci. 2023, 24, 10191. https://doi.org/10.3390/ijms241210191
Kowalówka M, Kosewski G, Lipiński D, Przysławski J. A Comprehensive Look at the -13910 C>T LCT Gene Polymorphism as a Molecular Marker for Vitamin D and Calcium Levels in Young Adults in Central and Eastern Europe: A Preliminary Study. International Journal of Molecular Sciences. 2023; 24(12):10191. https://doi.org/10.3390/ijms241210191
Chicago/Turabian StyleKowalówka, Magdalena, Grzegorz Kosewski, Daniel Lipiński, and Juliusz Przysławski. 2023. "A Comprehensive Look at the -13910 C>T LCT Gene Polymorphism as a Molecular Marker for Vitamin D and Calcium Levels in Young Adults in Central and Eastern Europe: A Preliminary Study" International Journal of Molecular Sciences 24, no. 12: 10191. https://doi.org/10.3390/ijms241210191